
H. Koknaroglu
Department of Animal Science
M.P. Hoffman
Department of Animal Science
D.D. Loy
Department of Animal Science
A. Trenkle
Department of Animal Science
J.D. Lawrence
Department of Economics, Iowa State University, Ames, IA 50011, Turkey
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 132-154







ABSTRACT
Purpose of the study was to integrate pasturing systems with drylot feeding systems and to compare performance, carcass characteristics and profitability of steers finished in the feedlot or backgrounded for various time on pasture and finished in the feedlot. For this purpose a three-year study, using 84 fall-born and 28 spring-born calves of similar genotypes each year, was designed. Fall and spring-born calves were started on test in May and October, respectively. Seven treatments were imposed: (1) fall-born calves directly into feedlot (FEEDLOT); (2 and 3) fall-born calves put on pasture with or without an ionophore and moved to the feedlot at the end of July (JI, JNI); (4 and 5) fall-born calves put on pasture with or without an ionophore and moved to the feedlot at the end of October (OI, ONI); (6 and 7) spring-born calves put on pasture with or without an ionophore and moved to the feedlot at the end of October (SI, SNI). Cattle on pasture receiving ionophore tended to gain faster (p>0.11), but lost this advantage in drylot (p>0.10). Overall, cattle started directly in the feedlot gained faster (p<0.001). All treatment groups graded 75% Choice or higher. Cattle started directly in the feedlot were the least profitable ($-46 vs $2; p<0.001). These results indicate that cattle fed on pasture for varying periods of time produced acceptable carcasses and that carcass price was an important variable affecting profitability. Thus, the timing of marketing is critical for optimizing profit.
PDF Abstract XML References Citation
Received: March 26, 2010;
Accepted: May 13, 2010;
Published: July 10, 2010
How to cite this article
H. Koknaroglu, M.P. Hoffman, D.D. Loy, A. Trenkle and J.D. Lawrence, 2011. Integration of Pasturing Systems for Cattle Finishing Programs. Asian Journal of Animal and Veterinary Advances, 6: 132-154.
DOI: 10.3923/ajava.2011.132.154
URL: https://scialert.net/abstract/?doi=ajava.2011.132.154
DOI: 10.3923/ajava.2011.132.154
URL: https://scialert.net/abstract/?doi=ajava.2011.132.154
INTRODUCTION
As the world population increases more cereal grain is needed for human use. About one sixth of the energy required by livestock is derived from grains (Oltjen and Becket, 1996). Thus alternative methods using less cereal grains in animal production must be developed. Ruminant livestock production takes advantage of forage production on approximately 25% of potentially arable land to minimize water and soil erosion. Cattle have the unique ability to use vast amounts of renewable resources from rangeland, pasture and crop residues or other by-products and convert them into food. Economics, energy saving and environmental issues such as soil conservation are becoming driving forces behind cattle feeding and thus one way to reduce the cost of production and improve soil conservation is to use pastures on highly erodable lands for grazing. Most countries produce beef by integrating pastures into feeding (Field and Taylor, 2003). However, all-forage systems for finishing cattle have not been particularly successful. Forage-based production systems may present some disadvantages in terms of product quality (Sainz and Paganini, 2004). The carcasses from cattle on the all-forage systems generally have not made the USDA Select grade level. Without grain, the final product typically has a poor quality and a milky or grassy flavor (Davis et al., 1981; Hedrick et al., 1983) and carcasses from these cattle are discriminated against by packers due to their lowered dressing percentage, higher cooler shrinkage and lower quality grade (Schroeder et al., 1980; Schoonmaker et al., 2002). In addition grass-finished cattle are discounted due to their differences in tenderness, color, juiciness and flavor (Chrystall, 1994; Baardseth et al., 1988; Hutchings and Illford, 1988; Melton, 1990). There is research to indicate that finishing cattle on corn in the feedlot for anytime beyond 30 days improves tenderness (Harrison et al., 1978; Dolezal et al., 1982). Maximum tenderness and lean growth can be achieved with 50 to 100 days grain feeding (Hedrick et al., 1983; Larick and Turner, 1990). Research have shown effectiveness of inclusion of pastures into beef cattle and lamb production in terms of cultural energy saving (Koknaroglu et al., 2006a, 2007a, b). Economic analyses of cattle finishing programs showed that feed costs are a major proportion of total variable costs (Koknaroglu et al., 2005; Demircan et al., 2007) and efficiently managed grazed grass can be a good alternative to decrease feed cost (O’Riordan and O’Kiely, 1996).
However there is limited or no research regarding the effects of grazing duration of fall and spring-born cattle on feedlot performance, carcass characteristics and economics. Thus the objective of this study was to integrate cattle pasturing systems with conventional feedlot feeding systems and to compare the systems in terms of cattle feedlot performance, carcass characteristics and economics.
MATERIALS AND METHODS
The three-year study was initiated with the establishment of a cool season grass pasture, smooth bromegrass, in May 1995, at the Western Iowa Research and Demonstration Farm at Castana, Iowa and was concluded in June 1999. In order to reduce the genetic variation and backgrounding differences among calves, the Stuart Ranch near Caddo, OK, was chosen as the provider of the calves. The composition of breed consisted of Hereford and Angus x Hereford cows and the cows were mated to either Angus or Hereford bulls to provide mainly crossbred calves. The cow herd was large enough to provide homogenous spring- and fall-born calves. Each year eighty-four fall-born calves were used in the initial phase of the study. The calves were backgrounded and given their calfhood vaccinations at the ranch. After 12 h of transportation they arrived at the research farm on April 17, 15 and 15, in the first, second and third years of the study, respectively. In order to alleviate the transportation stress and make calves accustomed to their environment, calves were given ground, mid-bloom alfalfa hay on arrival until May 7, 8 and 5, in the first, second and third years of the study, respectively. As a health precaution calves received one gram per head per day of chlortetracycline, which was fed at the rate of 113.5 gram per animal of four gram per 0.454 g AS-700® crumbles, top-dressed on the hay each morning. To aid in controlling coccidiosis, Amprolium® was added to the water source for two weeks after arrival of the calves. Before being placed on test on May 7, 8 and 5, calves were identified with an ear tag, implanted with Compudose® and injected with Ivomec® plus Flukocide ®. Cattle were tagged with Saber™ insecticide ear tags in the first week of June and this was repeated in the first week of August of each year. They were randomly allotted into 12 groups of 7 animals each and weighed on the average 167, 160 and 196 kg in the first, second and third years, respectively.
Five treatments, which involved four grazing and one control treatment, were assigned at random. On pasture supplement blocks either with monensin or without monensin were provided. The first treatment involved 14 steers (JI) receiving an ionophore and were stocked on smooth bromegrass pasture on May 7, 8 and 5, respectively and then moved to the feedlot on July 30, 29 and 28, respectively, to be fed the finishing diet during the remainder of the trial. A second treatment involved 14 steers (JNI) not receiving an ionophore and were stocked on smooth bromegrass pasture on May 7, 8 and 5, respectively and then moved to the feedlot on July 30, 29 and 28, respectively. A third pasture treatment involved 14 steers (OI) receiving an ionophore and were stocked on smooth bromegrass pasture on May 7, 8 and 5, respectively and then moved to the feedlot on October 22, 21 and 16, respectively. A fourth pasture treatment involved 14 steers (ONI) not receiving an ionophore and were stocked on smooth bromegrass pasture on May 7, 8 and 5, respectively and then moved to the feedlot on October 22, 21 and 16, respectively. A control group (FEEDLOT), 28 steers (seven head per pen), was placed directly into the feedlot after acclimation and was gradually adapted to an 82% concentrate diet containing whole shelled corn, ground alfalfa hay and a natural protein, vitamin and mineral supplement containing an ionophore and molasses. Cattle moved from pasture to the feedlot at various times received the same feed the control group received. In the feedlot when animals reached 363 kg, the supplement was changed from natural protein to a urea-based 40% crude protein, vitamin and mineral premix. About 100 days prior to slaughter, cattle were implanted with Revelor®.
The remaining two treatments involved obtaining 28 spring-born calves from the same ranch on September 17, 15 and 15, respectively and processing them in the same manner as fall-born calves. A sixth pasture treatment (SI) involved 14 spring-born steers receiving an ionophore and were stocked on smooth bromegrass pasture on October 1, September 30 and 29, respectively and then moved to the feedlot on October 22, 21 and 16, respectively, to be finished. A seventh pasture treatment (SNI) involved 14 spring-born steers not receiving an ionophore on pasture that were stocked on smooth bromegrass pasture on October 1, September 30 and 29, respectively and moved to the feedlot on October 22, 21 and 16, respectively, to be finished.
The pasture was enclosed by a 5 wire barbed wire fence with wood corner posts and 183 cm feet metal T-posts spaced every 6.10-7.62 m. An electrified wire was placed on the inside of the barbed wire fence on offset” insulators. The pasture consisted of 16 paddocks, each 0.69 ha acres in size and was subdivided using metal T-posts and braided 5-wire electric cable. Posts were spaced every 7.62 m with 7.62 cm wooden posts in draws and high spots. Two wires were strung with the bottom 9 notches high and the top 16 notches high.
Each grazing group had access to one paddock at a time. Cattle on the pasture were rotated on the basis of forage availability. In early summer, the cattle were not capable of consuming adequate forage to match the growth of the forage in all the paddocks, therefore they were rotated to a new paddock every three to four days. However later in the season when grass growth slowed, cattle were rotated about every two days to a new paddock. Nitrogen fertilizer was applied in two applications: one application of 45.4 kg per 0.405 ha applied in late April and the other of 36.3 kg per 0.405 ha applied in mid-August.
The feedlot facility consisted of pens with concrete floors, 26.5 by 4.3 m, with 7 m of overhead shelter at the north end of each lot. Steers were fed in fence-line concrete bunks, providing 53 cm of feedbunk space per animal, on the south side of the lot and one automatic waterer was shared between every two pens.
Feed allotment was determined daily prior to the morning feeding. Cattle were fed ad libitum and feed intake levels were provided such that feed was always available in the feedbunks. Feed provided was increased when the bunks in approximately one-half of the pens were completely empty at 0700 prior to the morning feeding.
Daily DMI was determined for each pen by recording the amount of air-dry feed fed from a feed wagon equipped with a digital scale and converting the amount to a DM basis. Feed samples were collected twice per week for dry matter determination. Every 28 days steers were weighed individually and average daily gain for that period and throughout the experiment were calculated. Dry matter intake by individual steers was represented by pen dry matter intake.
Carcass measurements: When the average weight of the pen of steers reached 522 kg, cattle were shipped for processing into beef to IBP in Denison, IA, which was 52 km from the farm. Cattle were shipped at 1700 the evening prior to slaughter and remained overnight in pens with access to water but not to feed at the packing plant. Steers were slaughtered between 0600 and 0730 following the overnight stay at the plant. Presence of liver abscesses was determined within 15 min after slaughter when the livers and other internal organs were removed on the processing line.
After a 24 h chill, backfat and ribeye area were measured on the 12th rib on the left half of each carcass. Backfat was measured to the nearest 0.11 cm using a ruler along the edge of the ribeye area grid. Ribeye area was measured to the nearest 0.645 square centimeter using a plastic grid with 10 dots per 6.45 cm2.
Carcass quality, yield grades and % KPH fat were called by USDA Meat Grading Service personnel. Quality grades, as provided by the USDA Meat Grading Service, to the nearest one-third of a grade, were converted to a numerical value. A quality grade of Select was equal to a value of five, high Select was equal to a value of six, low Choice was equal to a value of seven, average Choice was equal to a value of eight.
Economic analyses: A budget worksheet was prepared based on the finishing yearling steers budget worksheet in Livestock Enterprise Budgets for Iowa by John Lawrence, extension economist and Alan Vontalge, extension associate. Values used in the calculations were from the corresponding year of the experiment.
When calves were bought from Stuart Ranch money was paid for the group of calves. Thus feeder price per calf was calculated by dividing total money paid by total purchase weight of the calves which was the average weight at which cattle were started on feed and multiplying this figure by the individual weight of the calves. It was assumed that 100% of the money spent on buying cattle was borrowed and interest rate on that money was 10%. Days on feed were from the day cattle started on test through the day they were weight and shipped to the packing plant. Thus interest on feeder cattle price differed among treatment due to their days on feed at the farm. Prices received by farmers for each month in Iowa for corn and alfalfa were used in calculating corn and alfalfa costs. Natural and urea based supplement prices were obtained from the Livestock Enterprise Budgets for Iowa. Soybean meal prices for each month of each year were obtained from a source in Decatur, Illinois. Molasses prices were obtained from feedstuffs which is a weekly newspaper for agribusiness. In Feedstuffs the molasses price for Iowa was not mentioned, thus the average of Kansas City and Minneapolis was used as the price for Iowa. Improved pasture prices per acre were obtained from Livestock Enterprise Budgets for Iowa and pasture cost by treatments was calculated by considering the time cattle spent on pasture.
Interest on feed and other variable costs was 10% and was calculated as interest on these variables for half the days on feed.
Total revenue for each individual animal was obtained by multiplying hot carcass weight in kilogram with the price received for the carcass grade for that individual animal. The discount for Select and yield grade 4 carcasses differed each year and was included in the calculation.
Total variable costs were the sum of the feeder animal cost, feed cost, veterinary and health, machinery and equipment, marketing and miscellaneous and interest on feed and other costs. Fixed costs were associated with machinery, equipment and housing.
Income over variable cost was the product of subtraction of total variable cost from total revenue. Income over all costs, which is the equivalent of profit, was obtained by subtracting fixed costs from income over variable cost. Breakeven selling price for all costs was obtained by dividing the sum of total variable costs and fixed costs by the actual sale weight.
For price sensitivity analysis, the effect of a 5% increase or decrease in feeder price, carcass price and corn price was determined to observe their effects on profitability and breakeven price.
Statistical analyses: The experimental unit is a pen of cattle consisting of seven steers. There are seven treatment combinations, six with two replications and one with four replications. The analysis will take the form of a one-way analysis of variance with six degrees of freedom for treatments and 9 degrees of freedom within treatments or experimental error. The data were analyzed using the General Linear Model procedure of SAS and PDIFF statements were used to compare treatments.
RESULTS AND DISCUSSION
Performance of cattle both in feedlot and on pasture is provided in Table 1, cattle which spent more time on pasture, gained faster while on pasture than those that spent less time. OI cattle gained significantly faster than JNI, SI and SNI (p <0.03). ONI, JI and JNI cattle were intermediate in terms of ADG, however they differed significantly from SI and SNI (p<0.0001).
Relative larger frame size and age of OI and ONI cattle could be the reason for their faster gain on pasture. Because of their heavier weight on pasture OI and ONI had greater intestinal tract capacity relative to body size while basal metabolic rate was related to surface area of the animal (Van Soest, 1982). This assumes the animal’s capacity to consume forage increases faster than its surface area thus resulting in an increasing proportion of the consumed forage utilized for gain rather than maintenance (Bagley and Feazel, 1988).
The reason for SI and SNI cattle gaining less than others was the forage availability and quality. These cattle were stocked on pasture in the beginning of October and were removed around the end of October when the forage availability and quality was lower than other months. Higher lignin concentration, longer retention time in the rumen, decreased digestible dry matter intake, higher stem to leaf ratio with increased maturity were factors affecting performance of SI and SNI cattle (Grabber et al., 2009; Coleman et al., 2003; Fieser and Vanzant, 2004; Poppi et al., 1980, 1981; Van Soest, 1982; Jung et al., 1983; Burrit et al., 1984; Casler, 1986; Buxton, 1990; Burns et al., 1991; Mullahey et al., 1992).
| Table 1: | Performance of cattle both in feedlot and on pasture |
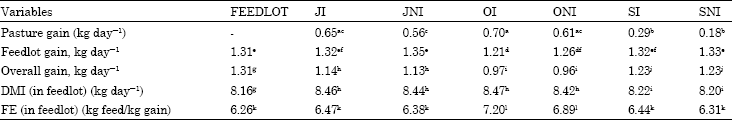 | |
| abcMeans with different superscripts in the same row are significantly different (p<0.03); defMeans with different superscripts in the same row are significantly different (p<0.05); ghijMeans with different superscripts in the same row are significantly different (p<0.01); klMeans with different superscripts in the same row are significantly different (p<0.02) | |
Cattle receiving an ionophore (OI, JI) tended to gain faster than their non-receiving ionophore counterparts (p> 0.1). SI cattle performed a little better than SNI cattle (p>0.09).
Similar results were also observed by others (Horn et al., 2005; Horn, 2006). Monensin increases molar proportion of propionate and decreases molar proportions of acetate and butyrate, methane production, protein degradation and ammonia production in the rumen (Min et al., 2005; Dinius et al., 1976; Richardson et al., 1976; Van Nevel and Demeyer, 1977; Prange et al., 1978; Bergen and Bates, 1984; Beede et al., 1986; Yang and Russell, 1993; Callaway et al., 1997). Thus with the use of monensin, the shift in VFA ratio in favor of propionate is a favorable change for meat producing animals (Min et al., 2005; Schelling, 1984) and decreased ruminal degradation of dietary protein results in more protein escaping the rumen to the lower tract thus increasing the efficiency of dietary protein by the ruminant animal (Whetstone et al., 1981).
When animals were moved into the feedlot, the trend seen on pasture was reversed and fall-born and spring-born cattle which spent less time on pasture (JI, JNI, SI and SNI) gained faster than those fall-born cattle that spent more time on pasture (OI, ONI; p<0.05). FEEDLOT cattle performed better than OI and ONI (p<0.04) but did not differ from JI, JNI, SI and SNI (p>0.9). It seems that cattle that spent less time on pasture (JI, JNI, SI and SNI) compensated for their relatively low gains on pasture by gaining faster when they were moved to the feedlot. These results agree with Ridenour et al. (1982) who found that on wheat pasture, cattle that grazed longer had higher ADG than those that spent less time on pasture and when they were moved to the feedlot they had lower gains than those that spent less time on wheat pasture and Choat et al. (2003) and Perry et al. (1972) who found that cattle gaining faster on pasture gained less in the feedlot when they were moved to the feedlot. OI and ONI had higher body weights than other treatments when they were moved to the feedlot. Thus one would expect them to gain faster in the feedlot since they were heavier. These results disagree with Koknaroglu et al. (2005) and Koknaroglu et al. (2006b) who found that as initial weight increases ADG increases.
It seems that the effect of monensin on pasture was reversed in the feedlot when cattle not receiving monensin on pasture tended to perform better (p>0.7). The reason for improvement in ADG in the feedlot by cattle not receiving ionophore on pasture could be the compensation of the cattle due to having lower gains during the grazing period. When Perry et al. (1971) fed a group of cattle in the feedlot and other groups of cattle on pasture receiving no concentrate, one third, two thirds and the same amount of concentrate as the ones in the feedlot, they found that ADG on pasture increased with increasing concentrate supplementation and when those cattle were moved to the feedlot, those which gained less on pasture gained faster in the feedlot.
Gain throughout experiment is also given in Table 1. As can be observed from the Table 1, FEEDLOT cattle had the highest gain throughout the experiment (p<0.001). Since FEEDLOT cattle spent all their time in the feedlot their lot gain is their gain throughout the experiment. SI and SNI groups were second in terms of gain throughout the experiment and differed significantly from other treatments (p<0.001). This group spent a relatively short time on pasture and most of their time in feedlot. That’s why they had better gains than JI, JNI and OI, ONI groups. Overall gain throughout the experiment for JI and JNI did not differ from each other (p>0.8), however it was different from other treatments (p<0.002). OI and ONI also did not differ from each other (p>0.6) but differed from other treatments (p<0.001). The overall gain results are in agreement with Oltjen et al. (1971). In their study steers fed 77 days initially on a forage diet gained less than those fed a concentrate diet and when forage receiving cattle received concentrate during the second period they did not sufficiently outgain the steers on a continues high concentrate diet. Thus, the steers on a continuous high concentrate diet in their study showed a slight advantage in ADG over the entire feeding trial. Similar results were also obtained by Perry et al. (1972), Flipot et al. (1986) and Berge et al. (1991) who reported experiments in which a feedlot system was compared with a system in which cattle were grazed initially and then finished in feedlot. The results of their experiments showed higher gains for the continuous feedlot cattle for the overall periods.
FEEDLOT cattle had lower DMI than other treatments (p<0.001). SI and SNI cattle had lower DMI than JI, JNI, OI and ONI cattle (p<0.004). SI and SNI cattle had similar DMI (8.32 and 8.30 kg d-1; p>0.5). JI and JNI did not differ from each other (p>0.6) and had similar DMI with OI and ONI (p>0.83). OI and ONI also did not differ from each other (p>0.08). The same results were obtained by Gill et al. (1991, 1992) who found that steers and heifers grazed shorter time in an Intensive Early Stocking (IES) system had lower dry matter intake in the feedlot than those grazed longer time in a Season Long Stocking (SLS). Lower dry matter intake of cattle started in the feedlot compared to cattle started on pasture and then moved to the feedlot later has been documented by other researchers (Sainz and Paganini, 2004; Oltjen et al., 1971; Lewis et al., 1990; Gill et al., 1993a; Myers et al., 1999). A similar result on effect of monensin on feed intake was obtained by Talbert et al. (1979) who found that in feedlot, cattle not receiving monensin on pasture had lower dry matter intake than cattle that received ionophore on pasture.
One of the reasons for cattle not receiving ionophore on pasture to have a lower dry matter intake in the feedlot could be due to the monensin they received in the feedlot. Baile et al. (1979) found that cattle receiving feed containing monensin showed an immediate and marked aversion to feed. A decrease in feed intake by cattle is frequently observed during the first four weeks of introduction of monensin (Gill et al., 1976; Perry et al., 1976; Dinius and Baile, 1977).
FEEDLOT cattle had lower dry matter intakes than other cattle because of their relatively young age and their lighter initial weights. In order to relate Initial Body Weight (IBW) to Dry Matter Intake (DMI), National Research Council (1996) used initial weight and DMI data obtained from commercial feedlots and found general equation of DMI = 4.54 + 0.0125 x IBW.The equation states that DMI increases with increase in initial weight. Koknaroglu et al. (2008) developed a DMI prediction equation in feedlot by using information generated by Iowa State University Feedlot Performance and Cost Monitoring program and found that average DMI during a feeding period is related to initial weight of cattle. Koknaroglu et al. (2005) found that steers started on feed at heavier weights had higher ADG than those started at lighter weights. Saubidet and Verde (1976) concluded that feed intake is related more to age than to weight. FEEDLOT cattle were younger and smaller framed when they were put in the feedlot therefore they consumed less feed.
Cattle spending less time on pasture had better FE than those spending more time. FEEDLOT, SI, SNI, JI and JNI did not differ in terms of FE (p>0.9). OI was the least efficient and was followed by ONI and these two groups differed from other treatments (p<0.02) but not from each other (p>0.08). Cattle not receiving an ionophore on pasture tended to be more efficient than their monensin receiving counterparts (p>0.08). It is reasonable to expect that cattle not receiving an ionophore on pasture to have a better feed efficiency since they tended to eat less in the feedlot and gain faster than those that received an ionophore on pasture. The reason for OI and ONIcattle to be less efficient was their relatively older age when they entered the feedlot. These cattle spent more time on pasture and when they were finished they were older than other treatments. These results agree with Koknaroglu et al. (2005, 2006b), whose findings suggest that lighter animals are more efficient than heavier animals. When compared either longer grazed cattle with shorter grazed cattle and finished in the feedlot or initially grazed cattle finished in the feedlot with those that went directly to the feedlot, Sainz and Paganini (2004), Gill et al. (1991, 1992, 1993a), Brandt et al. (1995) and Myers et al. (1999) found similar results.
The fact those lighter or younger cattle are more efficient than heavier or older animals can be explained by their growth potential due to growth hormone level circulating in their blood plasma (Verde and Trenkle, 1987). As cattle age, concentration of hormones circulating change and extent of chewing feed decreases. As hormones may change body composition and less chewing can reduce digestibility of incompletely processed diets, aging can reduce energetic efficiency (Gill et al., 1993a).
Another reason for OI and ONI to be less efficient is their higher empty body protein and increased size of the alimentary tract (Gill et al., 1993b; Rompala et al., 1990).
Carcass characteristics of cattle are given in Table 2, Attempts were made to finish cattle at 522 kg and as can be observed in Table 2 there were some small deviations from this weight. FEEDLOT cattle tended to be heavier than ONI (p>0.09) and all other treatments had similar final weights (p>0.9).
JNI cattle had the highest dressing percentage and differed significantly from FEEDLOT, OI, ONI and SI (p<0.02) but not from JI and SNI (p>0.3). Conversely FEEDLOT cattle had the lowest dressing percentage and were significantly lower than JI, JNI and SNI cattle (p<0.05). Most of the weight lost during transit is due to urinary and fecal losses. Because of this, the cattle having more feed consumption would be expected to defecate more per day and have a larger amount of fecal loss in transit. This might have been the case for OI and ONI to have a lower dressing percentage and the reason for cattle receiving ionophore on pasture to have lower dressing percentage.
Treatment did not have an effect on ribeye area (p>0.9). OI and ONI cattle, which spent more time on pasture, tended to have smaller REA than other treatments (p>0.7). Cattle receiving ionophore on pasture had slightly smaller REA (p>0.9) than those not receiving ionophore, with the exception of SI having larger REA than SNI (p>0.9).
FEEDLOT cattle had the highest backfat and differed significantly from OI, ONI and SNI cattle (p<0.004), but did not differ form JI, JNI and SI cattle (p>0.8). On the other hand there was a tendency for cattle spending more time on pasture to deposit less backfat. ONI had less backfat than FEEDLOT, JI, SI cattle (p<0.05). OI cattle had lower backfat then FEEDLOT and JI cattle (p<0.02). Cattle receiving ionophore on pasture had higher backfat than those not receiving ionophore (p>0.09).
KPH values are provided in Table 2. Cattle spending more time on pasture (OI, ONI) had lower KPH than cattle spending less time on pasture (JI, JNI and SNI; p<0.006). SNI and JNI had the highest KPH and it was significantly different from FEEDLOT, OI, ONI and SI cattle (p<0.02).
| Table 2: | Carcass characteristics of cattle |
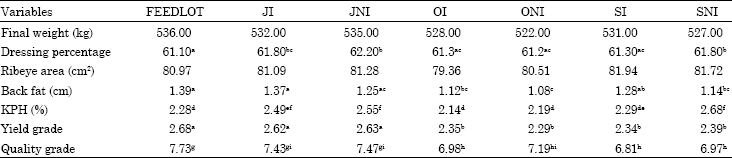 | |
| abcMeans with different superscripts in the same row are significantly different (p<0.05); defMeans with different superscripts in the same row are significantly different (p<0.04). ghiMeans with different superscripts in the same row are significantly different (p<0.03) | |
Cattle receiving an ionophore on pasture had lower KPH than those not receiving an ionophore on pasture (p<0.009).
Cattle receiving an ionophore on pasture had lower KPH and higher backfat than cattle not receiving an ionophore on pasture. This result disagrees with Lancaster et al. (1973), Davis et al. (1981), Bertrand et al. (1985), Schaake et al. (1993) and Sapp et al. (1996), who found that there was a positive relationship between backfat and KPH meaning that cattle which had more backfat also had more KPH. However, similar results to ours have been observed by Prichard et al. (1988), Bennett et al. (1995) and Sainz et al. (1995) who found that backfat decreased as KPH increased Sainz et al. (1995) or backfat increased as KPH decreased (Prichard et al., 1988; Bennett et al., 1995). Sainz et al. (1995) interpreted their results as not conclusive but suggested that there may be differential changes in different fat depots that may be related to the timing of the growth restriction. Limited intake of dietary energy during the period of subcutaneous adipose tissue development could have lasting effects on deposition of lipid in this depot without impairing fat accretion in later developing sites.
The growth rate of fatty tissues varies widely according to their location and the period of growth, early or late postnatal (Robelin, 1986). The growth of fatty tissue is mainly due to hypertrophy of adipose tissue cells and this growth in size is accompanied by an increase in lipid content of the fatty tissue. Thus subcutaneous fat develops later whereas internal fat matures earlier (Robelin, 1986). Ingle et al. (1972) compared the lipogenic capacity of adipose tissue obtained from different body sites and found that the internal fat depots were most active in younger lambs and calves, while the subcutaneous fat sites tended to have the greatest activity in mature ruminants (sheep and steers). Smith and Crouse (1984) investigated relative contributions of acetate, lactate and glucose to lipogenesis in bovine intramuscular and subcutaneous adipose tissue and found that acetate provided 70-80% of the acetyl units to in vitro lipogenesis in subcutaneous adipose tissue but only 10-25% in intramuscular adipose tissue. Conversely, glucose provided 1-10% of the acetyl units in subcutaneous adipose tissue, but 50-75% in the intramuscular depot.
One of the effects of using monensin could be the increased glucose production. Lomax and Baird (1983) and Armentano and Young (1983) found that propionate was the predominant precursor of glucose production and contributed up to 46 and 25 to 32% of the glucose output by the liver in lactating cattle and Holstein steers.
Even though glucose carbon is little used for fatty acid synthesis in ruminant adipose tissue, additions of glucose increased the rate of fatty acid synthesis from acetate by 3-10 times and by 20-50 times in caprine adipose tissue (Ballard et al., 1972; Yang and Baldwin, 1973; Liepa et al., 1978). On the other hand, acetate inhibited fatty acid synthesis from glucose in bovine and caprine adipose tissue in vitro (Bartos and Skarda, 1970; Whitehurst et al., 1978).
The reasons for cattle not receiving ionophore on pasture to have higher KPH could be the fact that these cattle had higher proportions of acetate in the VFA. And since the acetate is the main substrate for lipogenesis and inhibits fatty acid synthesis from glucose, they had a higher rate of lipogenesis and the relative early development of internal fat depots might have occurred during higher lipogenesis time.
In addition, the reason for cattle receiving monensin on pasture to have a higher backfat could be the fact that these cattle had higher ratios of propionate in total VFA production and thus higher glucose production. Since glucose increases fatty acid synthesis from acetate by several folds this increase in lipogenesis coinciding with fat deposition in subcutaneous adipose tissue might affect backfat.
Cattle spending longer time on pasture (OI and ONI) were leaner than other cattle. during the restrictive period, beef calves continue to develop skeletal structure, accumulate less body fat and more body protein than calves fed at a higher rate of gain (Phillips et al., 1991). Some research has shown that differences in body composition induced during feed restriction could be retained through the finishing period (Fox et al., 1972; Coleman and Evans, 1986).
Cattle spending longer time in the feedlot were fatter than those spending less time. Smith et al. (1984) reported that backfat thickness and the activities of several enzymes involved in lipogenesis were greater in steers fed a high concentrate, corn based diet versus steers fed a forage based, alfalfa pellet diet, even though the metabolizable energy intake was higher for the pelleted alfalfa diet. Thus a longer stay in the feedlot increased fat accumulated in the body.
In general cattle spending longer time in the feedlot had better quality grades and higher yield grades. The only exceptions for this were spring-born cattle which spent a long time in the feedlot and had low yield and quality grades. Harrison et al. (1978), Schroeder et al. (1980) and Skelley et al. (1978) found that increasing time in the feedlot increased marbling scores and quality grades.
Monensin increases propionate production, which in turn increases glucose production (Fieser, 2007; Lomax and Baird, 1983; Armentano and Young, 1983). Thus the reason for cattle not receiving monensin on pasture to have better quality grades could be that cattle not receiving an ionophore on pasture produce more VFA and probably more propionate and thus the effect of propionate on glucose production and intramuscular fat deposition.
For the economics part, four scenarios were considered. In the first scenario, the economics discussed used actual prices paid for feeder cattle and feed components and prices received for carcasses. In the second scenario, it was assumed that the cattle received the same feeder and fed cattle price using a 10 year average for feed components, feeder and fed cattle prices. For the third option, the same criteria as the second option were assumed except prices for feed components were derived from the corresponding months. In the fourth option, prices for corresponding months for feeder, fed cattle and feed components were used.
The reason for considering other scenarios is that feeder cattle, fed cattle and feed prices change depending upon time of purchase and marketing. In order to determine how profitability and other variables change based upon time it was decided to use this approach.
Scenario 1: Some economics variables are given in Table 3, spring-born cattle had lower feeder price than fall born cattle. Even though spring-born cattle had lower feeder price they had higher purchase price (p<0.0001) due to their heavier purchase weights.
Carcass price received for cattle reveals the importance of marketing time on carcass price received. Carcass price was lowest for FEEDLOT cattle (p<0.001). ONI cattle received the second lowest carcass price and it was significantly lower than other treatments except OI (p<0.024).
FEEDLOT cattle had significantly lower total revenue than JI, JNI, SI and SNI cattle (p<0.04) and similar total revenue with OI and ONI cattle (p>0.8). ONI cattle also had lower total revenue and significantly differed from JI and JNI cattle (p<0.02). The reason for these two groups to have low total revenue was that FEEDLOT cattle had lower carcass price and ONI cattle had low hot carcass weight and thus received relatively low revenue.
Feed cost was highest for FEEDLOT and lowest for SI and SNI cattle (p<0.001). Fall-born cattle removed from pasture in July and in October had significantly different feed costs than each other and than other treatments (p<0.001). It is expected for the FEEDLOT cattle to have the highest feed cost because of their longer days in the feedlot.
| Table 3: | Economic variables for treatments, 1st option |
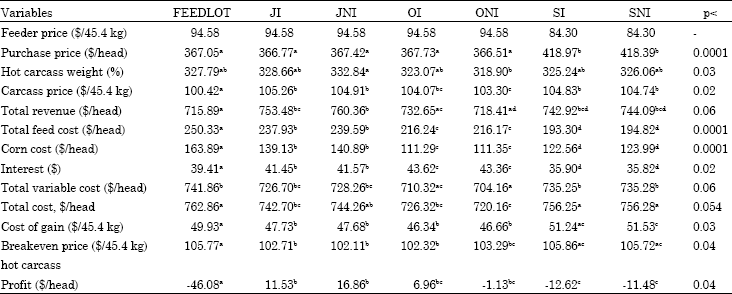 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
Even though SI and SNI cattle spent more time in the feedlot they had lower feed costs than OI and ONI because pasture cost associated with grazing was included in the feed cost section and since OI and ONI spent more time on pasture they had higher pasture costs.
Corn cost was directly related to time spent in the feedlot and thus cattle spending longer time in the feedlot had higher corn consumption. The cattle receiving the same pasture treatment and excluding the ionophore treatment (JI vs JNI, OI vs ONI, SI vs SNI) did not differ from each other (p>0.9), however they differed significantly from other pasture treatments (p<0.0001).
Total variable cost for ONI cattle was significantly lower than other treatments except OI cattle (p<0.06). FEEDLOT cattle had higher total variable cost than OI and ONI cattle (p<0.03). Other treatments were intermediate and they did not differ from each other (p>0.9). FEEDLOT cattle had higher total variable cost because of the high total feed cost and ONI and OI cattle had low total variable cost because of their low feed cost. Even though SI and SNI cattle had lower total feed cost than OI and ONI cattle they still had higher total variable cost because of the higher purchase price.
Breakeven price is given in Table 3 for hot carcass weight. JI, JNI and OI cattle had the lower breakeven hot carcass price than FEEDLOT, SI and SNI cattle (p<0.04). JNI and OI cattle had a little lower breakeven hot carcass price than JI and ONI because of their heavier hot carcass weights.
Cost of gain for 45.4 kg is also given in Table 3, the formula for total cost of gain is the total of all costs excluding purchase price divided by final weight minus initial weight. Total cost of gain eliminates the impact of purchase price on profitability and reflects the cost of production associated with performance of the cattle on pasture and in the feedlot. Total cost of production was related to time spent on pasture and in the feedlot. OI, ONI, JI and JNI cattle had lower total cost of gain than FEEDLOT, SI and SNI cattle (p<0.03). SNI had the highest cost of gain and significantly differed from FEEDLOT cattle. Cattle spending longer time on pasture had a lower cost of gain because the weight they gained on pasture was at a relatively cheaper cost. In their investigation of effect of backgrounding and season of birth on growth and cost of gain in cattle of two frame sizes, Buchanan-Buchanan-Smith et al. (1995) found that fall-born cattle which were not backgrounded had lower cost of gain than spring-born cattle which were backgrounded and not backgrounded.
Coffey and Moyer (1992) grazed a group of cattle for 75 days and another group 207 days and then finished them in the feedlot and obtained similar results. Cattle, with the shorter grazing period remained 56 days longer in the feedlot and had higher dry matter consumption, total feed cost and higher cost of gain than cattle grazed longer on pasture.
FEEDLOT cattle had the lowest profitability (p<0.001). JI and JNI had the highest profitability and significantly differed from SI and SNI (p<0.04) and did not differ from OI and ONI (p>0.7). FEEDLOT cattle had the lowest profitability because of their revenue due to lower carcass prices received and the higher corn consumption reflected in the total feed cost in the feedlot. JNI had better profitability than other treatments because of their heavier carcass weights and relatively higher total revenue.
Scenario 2: Economic variables for the second scenario are presented in Table 4, SI and SNI cattle had higher purchase prices than other treatments (p<0.001) because of their heavier initial weights. Treatments did not differ from each other in terms of total revenue (p>0.1).
SI and SNI cattle had the lowest feed costs in the feedlot (p<0.0001) whereas FEEDLOT cattle had the highest feed cost (p<0.0001). Corn cost in the feedlot was highest for FEEDLOT cattle and lowest for OI and ONI cattle and these treatments were different from other treatments (p<0.0001). JNI cattle had higher corn costs than SI cattle (p<0.04) and similar corn costs as JI and SNI cattle (p>0.6).
SI and SNI cattle had higher total variable costs and total all costs (p<0.0001) than other treatments and all other treatments had similar values (p>0.9). Cost of gain for 45.4 kg was higher for SI and SNI cattle (p<0.001) and JI and JNI cattle had lower cost of gain and differed from ONI cattle (p<0.03).
| Table 4: | Economic variables for treatments, 2nd option |
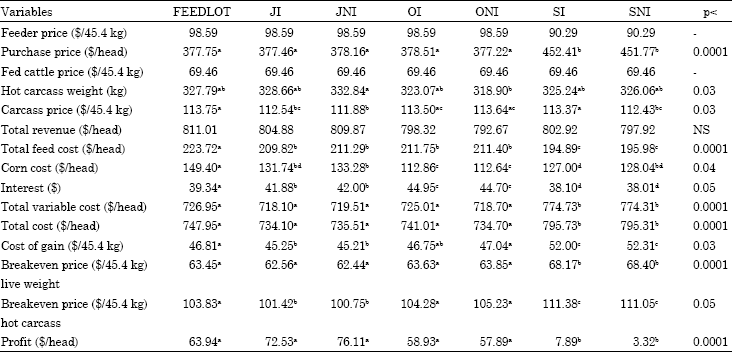 | |
| abcdeMeans with different superscripts in the same row are significantly different with respect to their p-values | |
SI and SNI cattle had higher breakeven prices for hot carcass weight (p<0.001). ONI and OI cattle had the next highest breakeven prices after SI and SNI cattle and were significantly different from JI and JNI cattle (p<0.04). SI and SNI cattle had high breakeven prices for hot carcass weight because of their heavier initial weight and thus higher purchase price. OI and ONI cattle also had higher breakeven hot carcass prices because of their lower overall ADG due to the longer time they spent on pasture.
SI and SNI had the lowest profitability (p<0.001) and JNI tended to differ from OI and ONI (p>0.08 and p>0.065, respectively). FEEDLOT cattle had similar profitability with JI, JNI, OI and ONI (p>0.6). Dikeman et al. (1985) found that cattle placed in the feedlot directly had lower breakeven live price, lower cost of retail product and were more profitable than those that were backgrounded on prairie hay and sorghum grain then later finished in the feedlot. They mentioned that rate of growth and carcass cutability were the main reasons for cost of retail product.
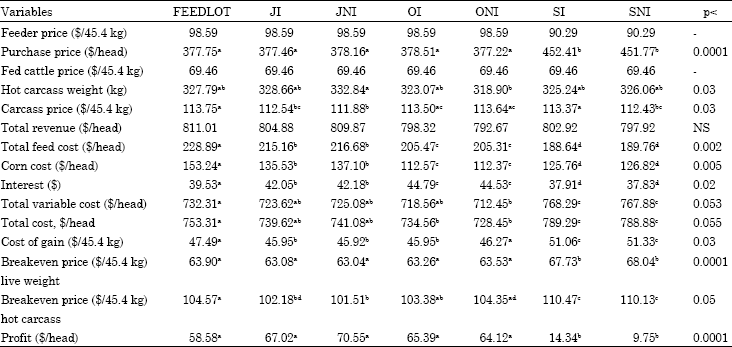
Scenario 3: Economic variables for the third scenario are presented in Table 5, since the same prices were used for feeder and fed cattle prices as in the second scenario; values and significance levels for feeder price, purchase price, carcass price, total revenue were the same as the ones in the second scenario.
FEEDLOT cattle had the highest feed cost followed by JI and JNI cattle (p<0.002). SI and SNI cattle had the lowest feed costs and were significantly different from OI and ONI (p<0.002) which were intermediate in terms of total feed cost.
Corn cost was higher for cattle spending longer time in the feedlot. Spring-born cattle and fall-born cattle removed to the feedlot in July and in October had different corn costs except those, which received the same treatment on pasture except for monensin (p<0.005).
Total cost including variable and fixed cost was higher for SI and SNI cattle (p<0.0005). OI and ONI had the lower total costs and differed from FEEDLOT cattle (p<0.055).
JI, JNI and ONI cattle had lower cost of gain than other treatments (p<0.03). SI and SNI cattle had the highest cost of gain whereas FEEDLOT and ONI cattle were intermediate.
| Table 5: | Economic variables for treatments, 3rd option |
 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
SI and SNI had higher total variable costs, costs of gain and breakeven prices for hot carcass and final weights and lower profitability than other treatments (p<0.0001).
SI and SNI cattle had lower profitability than other treatments because of their higher purchase price due to heavier initial weights. They also had higher costs of gain because of their heavier initial weights. Since animals were finished around 522 kg they put on about 295 kg, whereas cattle on other treatments put on 350 kg. Thus SI and SNI cattle had higher costs of gain due to fixed costs and other variable costs reflected in their gain.
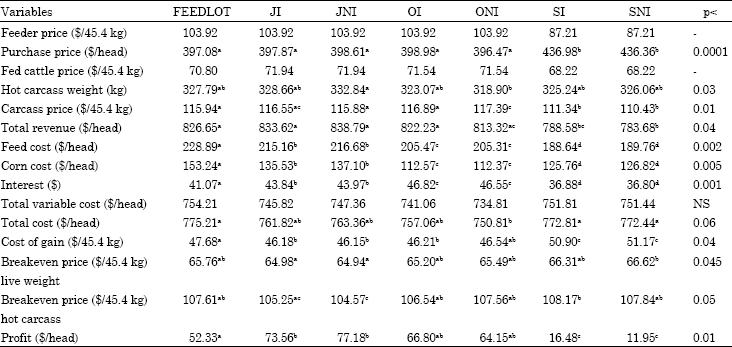
Scenario 4: Economic variables for the fourth scenario are presented in Table 6, SI and SNI cattle had higher purchase prices than other treatments (p<0.0001) even though they had lower feeder prices. Carcass prices, adjusted by using fed cattle price and dressing percentage, were lower for SI and SNI (p<0.01) because of their lower fed cattle price. Because of their heavier carcass weights or higher carcass prices FEEDLOT, JI, JNI and OI cattle had higher total revenue than spring-born cattle (p<0.04). ONI cattle had higher total revenue than SNI cattle (p<0.04) but had similar total revenue with SI (p>0.09).
Spring-born cattle and fall-born cattle removed to the feedlot in July and in October had different corn costs except those, which received the same treatment on pasture except for monensin. FEEDLOT cattle had the highest and SI and SNI cattle had the lowest feed costs (p<0.002).
In terms of corn cost, feed cost increased as time in the feedlot increased. FEEDLOT cattle had the highest corn cost and OI and ONI had the lowest corn cost and differed from each other (p<0.005). Cattle removed from pasture and placed in the feedlot at different times had different corn costs (p<0.005) except the cattle removed from pasture at the same time but differing in their monensin treatment on pasture.
ONI cattle had lower total cost than FEEDLOT and SI and SNI cattle (p<0.06). FEEDLOT and SI and SNI cattle had higher total cost because of higher purchase price (SI and SNI) and higher feed costs (FEEDLOT).
| Table 6: | Economic variables for treatments 4th option |
 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
Cost of gain for SI and SNI was higher than other treatments (p<0.0001) and cattle spending more time on pasture (except ONI) had lower cost of gain than FEEDLOT cattle (p<0.04). JNI had the lowest breakeven price for hot carcass and live weight and this was reflected in their profitability.
Generally spring-born cattle had higher costs of gain, breakeven price and lower profit than other treatments. The reason for SI and SNI having higher costs of gain, breakeven prices and lower profitability is because of their heavier initial weights and thus this is reflected in the calculations.
Price sensitivity analysis
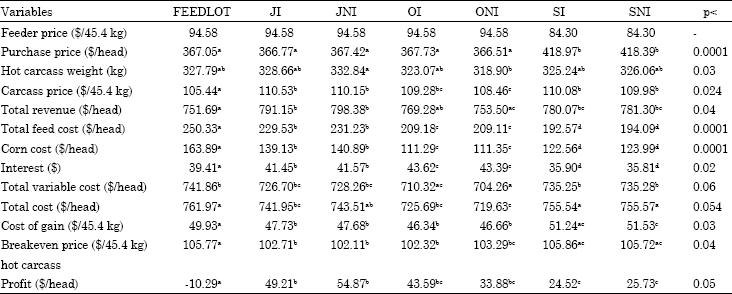
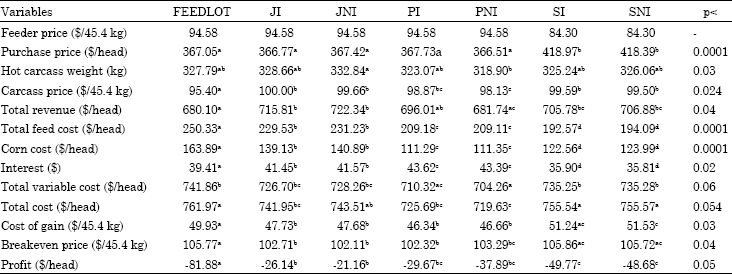
Carcass price: Price sensitivity analysis for carcass price is shown in Table 7 and 8. The values in Table 7 are the values when carcass price increases 5% and Table 8 values represent the values when carcass price decreases 5%. Since carcass price does not affect production and buying costs all the values except carcass price, total revenue and profit are the same for the treatments as the ones in Table 3.
| Table 7: | Economic variables when carcass price increases 5% |
 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
| Table 8: | Economic variables when carcass price decreases 5% |
 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
When carcass prices increased 5% from the actual case, FEEDLOT cattle still had negative profitability and other treatments had higher profitability with the same rankings as in the actual case. When carcass price decreased 5% all the treatments had negative profitability. Since the carcass is the end product that brings in revenue, the price received highly affects overall profitability. A better example of this was shown in the original case in which FEEDLOT cattle had similar carcass breakeven prices with SI (p>0.9) however they had lower profitability due to the lower carcass prices they received during marketing time. This illustrates the importance of marketing time.
Fluctuations in the fed cattle price exist depending upon the time cattle arrive at the market. Analyzing the data obtained from Iowa cattle producers using the Iowa State University Feedlot Performance and Cost Monitoring program between January 1988 and December 1997 Koknaroglu et al. (2005) found that cattle started on feed in the feedlot in spring had lower fed cattle prices than those that started in summer and fall.
Corn price: Price sensitivity analysis for corn price is provided in Table 9 and 10, when corn price increased or decreased 5%, corn cost increased or decreased accordingly and this price change in corn cost naturally affected feed costs and other costs associated with production. Change in corn price affected FEEDLOT cattle more than other cattle since they had higher corn consumption than other cattle. Five-percent change in corn price resulted in FEEDLOT cattle’s feed cost changing $8.19 whereas it was $6.24 on average for other treatments. When corn price increased or decreased 5%, the effect of this change reflected on hot carcass breakeven price was $1.19 for FEEDLOT cattle and it was $0.89 for SI and SNI cattle.
Since, cattle in Midwest feedlots are fed a ration consisting mostly of corn, the price of corn and the time cattle spent in the feedlot become more important. When the corn is relatively cheaper feeding cattle in the feedlot might be more advantageous, whereas when corn is relatively expensive utilization of pasture might be to the advantage of producers.
| Table 9: | Economic variables when corn price increases 5% |
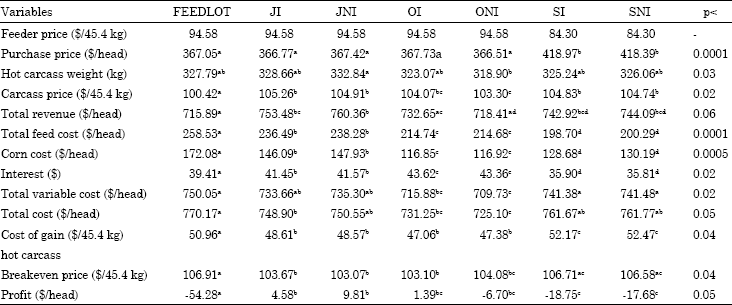 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
Feeder cattle price: Price sensitivity analysis for feeder cattle price is given in Table 11 and 12, since feeder cattle purchase price is a part of total variable costs and consequently the total of all costs, the change in feeder cattle price was reflected in hot carcass breakeven price and profit. SI and SNI cattle were heavier than other treatments when cattle were bought and thus they were influenced more by changes in feeder cattle price.
When feeder price increased or decreased 5%, SI and SNI cattle had higher change in their profitability ($23.23) than FEEDLOT cattle ($19.68).
Feeder cattle price also fluctuates with supply of calves coming to the market. As was the case in this study, spring-born cattle had lower feeder cattle prices but were heavier. Thus in the Midwest it might be more beneficial to buy lighter fall-born calves in spring and backgrounding them on pasture at a low cost during summer and fall. In the South where winter pasture is available to cattle, buying spring-born calves at a cheaper price in fall and backgrounding them on pasture before finishing could be more profitable.
| Table 10: | Economic variables when corn price decreases 5% |
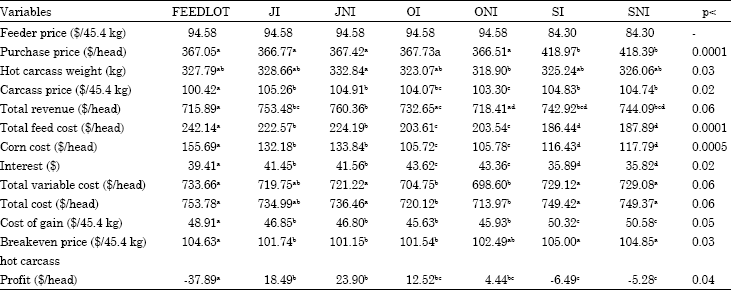 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
| Table 11: | Economic variables when feeder price increases 5% |
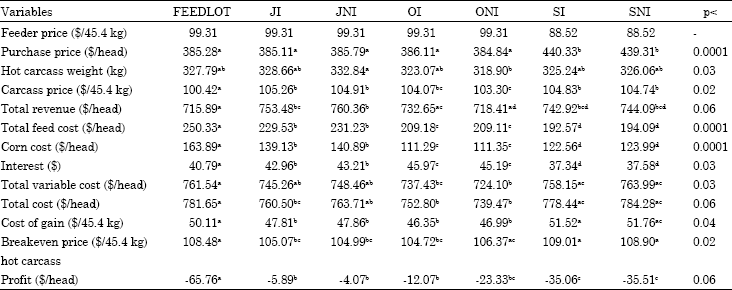 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
| Table 12: | Economic variables when feeder price decreases 5% |
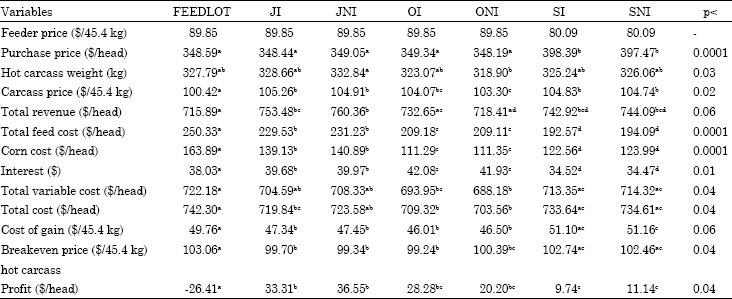 | |
| abcdMeans with different superscripts in the same row are significantly different with respect to their p-values | |
IMPLICATIONS
This study indicates that cattle receiving an ionophore on pasture and cattle spent more time on pasture gained faster while on pasture than those that spent less time and not receiving an ionophore and when cattle were moved to the feedlot, the trend seen on pasture was reversed and cattle not receiving an ionophore on pasture and cattle spent less time on pasture gained faster than those that spent more time on pasture and receiving an ionophore. Thus it would be more beneficial to provide an ionophore when cattle spend longer time on pasture and grazing cattle until end of July would result in better overall performance than other pasture treatments. These results also show that cattle grazed on pasture for various time periods had acceptable and comparable carcass grades. The price received for carcasses was influenced by the time of year the cattle were marketed and this was an important factor affecting profitability.
REFERENCES
- Armentano, L.E. and J.W. Young, 1983. Production and metabolism of volatile fatty acids, glucose and CO2 in steers and the effects of monensin on volatile fatty acid kinetics. J. Nutr., 113: 1265-1277.
Direct Link - Baardseth, P., G. Skrede, T. Naes, M.S. Thomassen, A. Iversen and L. Kaaber, 1988. A comparison of CIE (1976) L*a*b* values obtained from two different instruments on several food commodities. J. Food Sci., 53: 1737-1742.
CrossRefDirect Link - Bagley, C.P. and J.I. Feazel, 1988. Production, economics and acceptability of forage-fed beef. Beef Cattle Sci. Book, 22: 295-301.
Direct Link - Baile, C.A., C.L. McLaughlin, E.L. Potter and W. Chalupa, 1979. Feeding behavior changes of cattle during introduction of monensin with roughage or concentrate diets. J. Anim. Sci., 48: 1501-1508.
Direct Link - Ballard, F.J., O.H. Fissell and I.G. Jarrett, 1972. Effects of carbohydrate availability on lipogenesis in sheep. Biochem. J., 126: 193-200.
Direct Link - Beede, D.K., G.T. Schelling, G.E. Mitchell Jr., R.E. Tucker, W.W. Gill, S.E. Koenig and T.O. Lindsey, 1986. Nitrogen utilization and digestibility by growing steers and goats of diets that contain monensin and low crude protein. J. Anim. Sci., 62: 857-863.
Direct Link - Bennett, L.L., A.C. Hammond, M.J. Williams, W.E. Kunkle, D.D. Johnson, R.L. Preston and M.F. Miller, 1995. Performance, carcass yield and carcass quality characteristics of steers finished on rhizoma peanut (Arachis glabrata)-tropical grass pasture or concentrate. J. Anim. Sci., 73: 1881-1887.
PubMedDirect Link - Berge, P., Y. Geay and D. Micol, 1991. Effect of feeds and growth rate during the growing phase on subsequent performance during the fattening period and carcass composition of young dairy breed bulls. Livest. Prod. Sci., 28: 203-222.
CrossRef - Bergen, W.G. and D.B. Bates, 1984. Ionophores: Their effect on production efficiency and mode of action. J. Anim. Sci., 58: 1465-1483.
PubMedDirect Link - Burns, J.C., K.R. Pond and D.S. Fisher, 1991. Effect of grass species on grazing steers: II. Dry matter intake and digesta kinetics. J. Anim. Sci., 69: 1199-1204.
Direct Link - Burrit, E.A., A.S. Bittner, J.C. Street and M.J. Anderson, 1984. Corrrelations of phenolic acids and xylose content of cell wall with in vitro dry matter digestibility of three maturing grasses. J. Dairy Sci., 67: 1209-1213.
Direct Link - Buxton, D.R., 1990. Cell-wall component in divergent germplasm of four perennial forage grass species. Crop Sci., 30: 402-408.
Direct Link - Callaway, T.R., A.M.S. Carneiro de Melo and J.B. Russell, 1997. The effect of nisin and monensin on ruminal fermentations in vitro. Curr. Microbiol., 35: 90-96.
Direct Link - Choat, W.T., C.R. Krehbiel, G.C. Duff, R.E. Kirksey and L.M. Lauriault et al., 2003. Influence of grazing dormant native range or winter wheat pasture on subsequent finishing cattle performance, carcass characteristics and ruminal metabolism. J. Anim. Sci., 81: 3191-3201.
Direct Link - Coleman, S.W. and B.C. Evans, 1986. Effect of nutrition, age and size on compensatory growth in two breeds of steers. J. Anim. Sci., 63: 1968-1982.
Direct Link - Davis, G.W., A.B. Cole, W.R. Backus and S.L. Melton, 1981. Effect of electrical stimulation on carcass quality and meat palatability of beef from forage and grain-finished steers. J. Anim. Sci., 53: 651-657.
Direct Link - Demircan, V., H. Koknaroglu, H. Yilmaz and Z. Dernek, 2007. Economic analysis of beef cattle farms in Turkey. J. Applied Anim. Res., 31: 143-148.
CrossRefDirect Link - Dikeman, M.E., K.N. Nagele, S.M. Myers, R.R. Schalles, D.H. Kroph, C.L. Kastner and F.A. Russo, 1985. Accelerated versus conventional beef production and processing. J. Anim. Sci., 61: 137-150.
Direct Link - Dinius, D.A., M.E. Simpson and P.B. Marsh, 1976. Effect of monensin fed with forage on digestion and ruminal ecosystem of steers. J. Anim. Sci., 42: 229-234.
PubMedDirect Link - Dinius, D.A. and C.A. Baile, 1977. Beef cattle response to a feed intake stimulant given alone and in combination with propionate enhancer. J. Anim. Sci., 45: 147-153.
Direct Link - Fieser, B.G. and E.S. Vanzant, 2004. Interactions between supplement energy source and tall fescue hay maturity on forage utilization by beef steers. J. Anim. Sci., 82: 307-318.
Direct Link - Fox, D.G., R.R. Johnson, R.L. Preston, T.R. Dockerty and E.W. Klosterman, 1972. Protein and energy utilization during compensatory growth in beef cattle. J. Anim. Sci., 34: 310-318.
Direct Link - Gill, D.R., J.R. Martin and R. Lake, 1976. High, medium and low corn silage diets with and without monensin for feedlot steers. J. Anim. Sci., 43: 363-368.
Direct Link - Gill, D.R., F.T. McCollum, C.A. Strasia, J.J. Martin and R.L. Ball, 1991. Effect of Growing Diets and Grazing Regimen on Feedlot Performance and Carcass Traits. Oklahoma State University, Miscellaneous publication- Agricultural Experiment Station, USA., pp: 114-121.
Direct Link - Grabber, J.H., D.R. Mertens, H. Kim, C. Funk, F. Lu and J. Ralph, 2009. Cell wall fermentation kinetics are impacted more by lignin content and ferulate cross-linking than by lignin composition. J. Sci. Food Agric., 89: 122-129.
CrossRef - Harrison, A.R., M.E. Smith, D.M. Allen, M.C. Hunt, C.L. Kastner and D.H. Kroph, 1978. Nutritional regime on quality and yield characteristics of beef. J. Anim. Sci., 47: 383-388.
Direct Link - Hedrick, H.B., J.A. Paterson, A.G. Matches, J.D. Thomas, R.E. Morrow, W.C. Stringer and R.J. Lipsey, 1983. Carcass and palatability characteristics of beef produced on pasture, corn silage and corn grain. J. Anim. Sci., 57: 791-801.
Direct Link - Horn, G.W., P.A. Beck, J.G. Andrae and S.I. Paisley, 2005. Designing supplements for stocker cattle grazing wheat pasture. J. Anim. Sci., 83: E69-E78.
Direct Link - Horn, G.W., 2006. Growing cattle on winter wheat pasture: Management and herd health considerations. Vet. Clin. Food Anim., 22: 335-356.
CrossRef - Hutchings, J.B. and P.J. Illford, 1988. The perception of food texture the philosophy of the breakdown path. J. Texture Stud., 19: 103-109.
CrossRef - Ingle, D.L., D.E. Bauman and U.S. Garrigus, 1972. Lipogenesis in the ruminant: In vitro study of tissue sites, carbon source and reducing equivalent generation for fatty acid synthesis. J. Nutr., 102: 609-616.
Direct Link - Jung, H.G., G.C. Fahey Jr. and N.R. Merchen, 1983. Effects of ruminant digestion and metabolism on phenolic monomers of forages. Br. J. Nutr., 50: 637-651.
CrossRef - Koknaroglu, H., D.D. Loy, D.E. Wilson, M.P. Hoffman and J.D. Lawrence, 2005. Factors affecting beef cattle performance and profitability. Prof. Anim. Sci., 21: 286-296.
Direct Link - Koknaroglu, H., A. Ali, D.G. Morrical and M.P. Hoffman, 2006. Breeding for sustainability: Effect of breed on cultural energy expenditure of lamb production. Czech J. Anim. Sci., 51: 391-399.
Direct Link - Koknaroglu, H., M.T. Toker and Y. Bozkurt, 2006. Effect of zeolite and initial weight on feedlot performance of brown Swiss cattle. Asian J. Anim. Vet. Adv., 1: 49-54.
CrossRefDirect Link - Koknaroglu, H., K. Ekinci and M.P. Hoffman, 2007. Cultural energy analysis of pasturing systems for cattle finishing programs. J. Sustain. Agric., 30: 5-20.
CrossRefDirect Link - Koknaroglu, H., A. Ali, K. Ekinci, D.G. Morrical and M.P. Hoffman, 2007. Cultural energy analysis of lamb production in the feedlot or on pasture and in the feedlot. J. Sustain. Agric., 30: 95-108.
CrossRefDirect Link - Lancaster, L.R., R.R. Frahm and D.R. Gill, 1973. Comperative feedlot performance and carcass traits between steers allowed a postweaning growing period and steers placed on a finishing ration at weaning. J. Anim. Sci., 37: 632-636.
Direct Link - Larick, D.K. and B.E. Turner, 1990. Flavor characteristics of forage and green fed beef as influence by phospholipid and fatty acid composition differences. J. Food Sci., 55: 312-317.
Direct Link - Lewis, J.M., T.J. Klopfenstein, R.A. Stock and M.K. Nielsen, 1990. Evaluation of intensive vs extensive systems of beef production and the effect of level of beef cow milk production on postweaning performance. J. Anim. Sci., 68: 2517-2524.
Direct Link - Liepa, G.U., D.C. Beitz and J.R. Linder, 1978. Fatty acid synthesis in ruminating and non-ruminating goats. J. Nutr., 108: 1733-1739.
Direct Link - Lomax, M.A. and G.D. Baird, 1983. Blood flow and nutrient exchange across the liver and gut of the dairy cow: Effect of lactation and fasting. Br. J. Nutr., 49: 481-496.
CrossRef - Melton, S.L., 1990. Effects of feeds on flavor of red meat: A review. J. Anim. Sci., 68: 4421-4435.
Direct Link - Min, B.R., W.E. Pinchak, J.D. Fulford and R. Puchala, 2005. Effect of feed additives on in vitro and in vivo rumen characteristics and frothy bloat dynamics in steers grazing wheat pasture. Anim. Feed Sci. Technol., 123-124: 615-629.
CrossRef - Myers, S.E., D.B. Faulkner, T.G. Nash, L.L. Berger and D.F. Parret, 1999. Performance and carcass traits of early-weaned steers receiving either a pasture growing period or a finishing diet at weaning. J. Anim. Sci., 77: 311-322.
Direct Link - Oltjen, R.R., T.S. Rumsey and P.A. Putnam, 1971. All forage diets for finishing beef cattle. J. Anim. Sci., 32: 327-333.
Direct Link - Oltjen, R.R. and J.L. Becket, 1996. Role of ruminant livestock in sustainable agricultural systems. J. Anim. Sci., 74: 1406-1409.
Direct Link - Perry, T.W., D.A. Huber, G.O. Mott and C.L. Rhykerd, 1971. Effect of level of pasture supplementation on pasture, drylot and total performance of beef cattle: I. Spring pasture. J. Anim. Sci., 32: 744-748.
Direct Link - Perry, T.W., D.A. Huber, G.O. Mott, C.L. Rhykerd and R.W. Taylor, 1972. Effect of level of pasture supplementation on pasture, drylot and total performance of beef cattle: II. Spring plus summer pasture. J. Anim. Sci., 34: 647-652.
Direct Link - Perry, T.W., W.M. Beeson and M.T. Mohler, 1976. Effect of monensin on beef cattle performance. J. Anim. Sci., 42: 761-765.
Direct Link - Phillips, W.A., J.W. Holloway and S.W. Coleman, 1991. Effect of pre- and post weaning management system on the performance of Brahman crossbred feeder calves. J. Anim. Sci., 69: 3102-3111.
Direct Link - Poppi, D.P., D.J. Minson and J.H. Ternouth, 1980. Studies of cattle and sheep eating leaf and stem fractions of grasses. I. The voluntary intake, digestibility and retention time in the reticulo-rumen. Aust. J. Agric. Res., 32: 99-108.
Direct Link - Poppi, D.P., D.J. Minson and J.H. Ternouth, 1981. Studies of cattle and sheep eating leaf and stem fractions of grasses. 3. The retention time in the rumen of large feed particles. Aust. J. Agric. Res., 32: 123-137.
CrossRef - Prange, R.W., C.L. Davis and J.H. Clark, 1978. Propionate production in the rumen of Holstein steers fed either a control or monensin supplemented diet. J. Anim. Sci., 46: 1120-1124.
Direct Link - Prichard, D.L., J.E. Bertrand and D.D. Johnson, 1988. Effect of backgrounding regime on animal performance and carcass characteristics of beef steers finished for various periods in the feedlot. Florida Beef Cattle Research Report, pp: 177-183. http://agris.fao.org/agris-search/search/display.do?f=1990/v1605/US9011760.xml;US9011760.
- Richardson, L.F., A.P. Raun, E.L. Potter, C.O. Cooley and R.P. Ratchmacher, 1976. Effect of monensin on ruminal fermentation in vitro and in vivo. J. Anim. Sci., 43: 657-664.
Direct Link - Ridenour, K.W., H.E. Kiesling, G.P. Lofgreen and D.M. Stiffler, 1982. Feedlot performance and carcass characteristics of beef steers grown and finished under different nutrition and management programs. J. Anim. Sci., 54: 1115-1119.
Direct Link - Robelin, J., 1986. Growth of adipose tissues in cattle; Partitioning between depots, chemical composition and cellularity: A review. Livest. Prod. Sci., 14: 349-364.
CrossRef - Rompala, R.E., T.A. Hoagland and J.A. Meister, 1990. Modifications in growth and morphology of ovine jejunal and ruminal epithelia as affected by inert dietary substances. J. Anim. Sci., 68: 2530-2535.
Direct Link - Sainz, R.D., F. de la Torre and J.W. Oltjen, 1995. Compensatory growth and carcass quality in growth-restricted and refed beef steers. J. Anim. Sci., 73: 2971-2979.
Direct Link - Sainz, R.D. and R.F.V. Paganini, 2004. Effects of different grazing and feeding periods on performance and carcass traits of beef steers. J. Am. Sci., 82: 292-297.
Direct Link - Saubidet, C.L. and L.S. Verde, 1976. Relationship between liveweight, age and dry matter intake for beef cattle after different levels of food restriction. Anim. Prod., 22: 61-69.
CrossRef - Schaake, S.L., G.C. Skelly, E. Halpin, L.W. Grimes, R.B. Brown, D.L. Cross and C.E. Thompson, 1993. Carcass and meat sensory traits of steers finished on fescue and clover, summer forage, or different periods in drylot. J. Am. Sci., 71: 3199-3205.
Direct Link - Schelling, G.T., 1984. Monensin mode of action in the rumen. J. Anim. Sci., 58: 1518-1527.
Direct Link - Schoonmaker, J.P., S.C. Loerch, F.L. Fluharty, H.N. Zerby and T.B. Turner, 2002. Effect of age at feedlot entry on performance and carcass characteristics of bulls and steers. J. Am. Sci., 80: 2247-2254.
Direct Link - Skelley, G.C., R.L. Edward, F.B. Wardlaw and A.K. Torrence, 1978. Selected high forage rations and their relationship to beef quality, fatty acids and amino acids. J. Am. Sci., 47: 1102-1108.
Direct Link - Smith, S.B. and J.D. Crouse, 1984. Relative contributions of acetate, lactate and glucose to lipogenesis in bovine intramuscular and subcutaneous adipose tissue. J. Nutr., 114: 792-800.
Direct Link - Smith, S.B., R.L. Prior, C.L. Ferrel and H.J. Mersmann, 1984. Interrelationships among diet, age, fat deposition and lipid metabolism in growing steers. J. Nutr., 114: 153-162.
Direct Link - Van Nevel, C.J. and D.I. Demeyer, 1977. Effect of monensin on rumen metabolism in vitro. Applied Environ. Microbiol., 34: 251-257.
Direct Link - Verde, L.S. and A. Trenkle, 1987. Concentrations of hormones in plasma from cattle with different growth potentials. J. Anim. Sci., 64: 426-432.
Direct Link - Whetstone, H.D., C.L. Davis and M.P. Bryant, 1981. Effect of monensin on breakdown of protein by ruminal microorganisms in vitro. J. Am. Sci., 53: 803-809.
PubMedDirect Link - Whitehurst, G.B., D.C. Beitz, M.A. Pothoven, W.R. Ellison and M.H. Crump, 1978. Lactate as a precursor of fatty-acid in bovine adipose tissue. J. Nutr., 108: 1806-1811.
Direct Link - Yang, Y.T. and R.L. Baldwin, 1973. Preparation and metabolism of isolated cell from ovine adipose tissue. J. Am. Sci., 56: 350-365.
PubMedDirect Link - Yang, C.M. and J.B. Russell, 1993. The effect of monensin supplementation on ruminal ammonia accumulation in vivo and the numbers of amino acid-fermenting bacteria. J. Anim. Sci., 71: 3470-3476.
PubMedDirect Link