
M. Alamer
Department of Animal and Fish Production, College of Agricultural Sciences and Foods, King Faisal University, P.O. Box 400, Hofuf 31982, Saudi Arabia
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 12 | Page No.: 1153-1169







ABSTRACT
Prolactin is a multi functional hormone that is believed to possess various diverse biological potencies than all other pituitary hormones combined. It is widely understood that prolactin is positively associated with ambient temperature that may indicate involvement of prolactin in an acclimatory responses to higher ambient temperature. The current review presented the evidences that indicate the possible modulatory role of prolactin in some thermoregulatory mechanisms during hot climates in domestic ruminants. The observed thermoregulatory failure with the suppression of prolactin response to heat exposure in ruminants suggests the modulation of prolactin of some thermoregulatory mechanisms. Down regulation in the expression of genes associated with prolactin signaling pathways in some target tissues which is induced by thermal exposure may be consistent with mechanisms to down-regulate some metabolic process directed to reduce heat increment. Evidence suggests that prolactin may affect body fluid regulation by maintaining extracelluar fluid volume during heat exposure and hence supporting heat dissipation. Prolactin may also control seasonal pelage growth cycle probably directed to facilitate heat loss during summer season. It can be concluded that higher circulating prolactin may modulate some thermoregulatory processes during heat exposure. This is likely to be associated with modulation of some mechanisms of heat dissipation and heat production oriented to support homeothemy.
PDF Abstract XML References Citation
Received: May 10, 2011;
Accepted: August 27, 2011;
Published: November 05, 2011
How to cite this article
M. Alamer, 2011. The Role of Prolactin in Thermoregulation and Water Balance During Heat Stress in Domestic Ruminants. Asian Journal of Animal and Veterinary Advances, 6: 1153-1169.
DOI: 10.3923/ajava.2011.1153.1169
URL: https://scialert.net/abstract/?doi=ajava.2011.1153.1169
DOI: 10.3923/ajava.2011.1153.1169
URL: https://scialert.net/abstract/?doi=ajava.2011.1153.1169
INTRODUCTION
Prolactin is a polypeptide hormone secreted by lactotrophs, acidophilic-staining cells in the anterior pituitary gland, given the hormone has a broad variety of biological actions. Prolactin is therefore known to have over 300 separate biological actions in a number of diverse species which is far beyond the sum of all recognized biological roles of other pituitary hormones (Bole-Feysot et al., 1998). The biological activities of this polypeptide hormone are extensively reciprocal with reproduction, osmoregulation, metabolism, immunoregulation, brain function and behavior (Freeman et al., 2000; Ben-Jonathan et al., 2006; Eltayeb et al., 2010). In fact, in view of the many roles played by prolactin, researchers have proposed the hormone might be renamed versatilin or omnipotin" (Bern and Nicoll, 1968). Further, some research findings point to a possible connection between prolactin and core body temperature during heat exposure. Therefore, the purpose of this presentation was to survey current findings that indicate the possible modulation of prolactin of some thermoregulatory mechanisms during hot climates in domestic ruminants.
Seasonal effect on prolactin secretion: Studies recognize that a number of stimuli, including environmental factors such as ambient temperature and photoperiod exert some influence on the levels of circulating prolactin in several species (Tucker, 1982; Archawaranon, 2006; Roy and Prakash, 2007). It is also well documented that in several mammals exposure to normal alterations of day length and ambient temperature, influences prolactin levels. Therefore, the circulating prolactin displays a clear Circannual rhythm (Karsch et al., 1989) with the highest circulating prolactin levels occurring during summer whitest the lowest concentrations occurring during winter months (Sergent et al., 1988; Brunet and Sebastian, 1991; Curlewis, 1992). Table 1 summarizes data from a number of studies on ruminants that compare prolactin response to environmental temperatures. Smith et al. (1977) revealed that the prolactin value during summer can rise six to seven fold over the lowest value of winter. In addition to showing sensitivity to seasonal variations, prolactin concentration in the plasma shows patterned fluctuations associated with rapid or progressive changes in air temperature. It has been reported that prolactin values in Holstein heifers increase by more than 3 fold if ambient temperature is raised from 18 to 32°C (Ronchi et al., 2001). Also, a rise in ambient temperature from 21 to 31°C increased prolactin values by about 44% in Holstein heifers. However, Smith et al. (1977) reported a 4 fold increase of prolactin in Hereford steers maintained on 40°C compared to that at thermoneutral zone. In addition, lowering ambient temperature from 20-21°C to 4-7°C in cattle decreased serum prolactin by 55-80% (Smith et al., 1977; Wettemann et al., 1982). These studies clearly indicate there is a correlation between seasonal changes in ambient temperature and serum prolactin concentration in the ruminant animal and this variation in prolactin levels appears to be involved in acclimating to seasonal changes.
Effect of heat stress on prolactin release: Heat stress develops when the effective temperature exerted by the surrounding environment surpasses that of the animal’s thermoneutral zone. Some physiological processes are modified during heat exposure in an attempt to support heat balance and hence homeothermy is less disrupted. Therefore, the secretion pattern of various hormones has been known to be modified in heat-stressed animals. Roman-Ponce et al. (1981) demonstrated that sun exposure for a continuous 20 weeks increased plasma prolactin values from 38 to 86 ng mL-1 in lactating cows (Table 1). Also, induced heat stress by solar radiation exposure increased serum prolactin by about 90% in goats (Sergent et al., 1985). Therefore, these results have led some investigators to propose that plasma prolactin level can be used as index of thermal stress tolerance (Chemineau and Ravault, 1984; Barb et al., 1991). Scharf et al. (2010) concluded that prolactin may be combined with rectal temperature in the identification of breed differences in heat sensitivity. Furthermore, research shows that lactotrophs sensitivity to prolactin stimulation is also affected by the prevailing air temperature (Table 2). Some studies have demonstrated that prolactin response to TRH challenge in vivo was significantly higher at higher temperatures compared to lower temperatures but no stimulatory response could be detected at lower temperatures in Holstein heifers (Wettemann and Tucker, 1976; Wettemann et al., 1982). Prolactin values following TRH increased by about 6.8 fold when maintained at 30°C compared to 3.5 fold at 22°C (Barb et al., 1991). Also, when tested under in vitro conditions, lactotrophs harvested from pigs under heat stress were significantly more responsive to TRH than those from animals at thermoneutrality (Matteri et al., 1994; Matteri and Becker, 1996). This research indicates that heat stress affects the synthesis and secretion of prolactin which is associated with higher cellular content of prolactin available for secretion at the level of the lactotrophs (Matteri et al., 1994).
| Table 1: | Effect of environmental temperature on plasma/serum prolactin concentrations in ruminants |
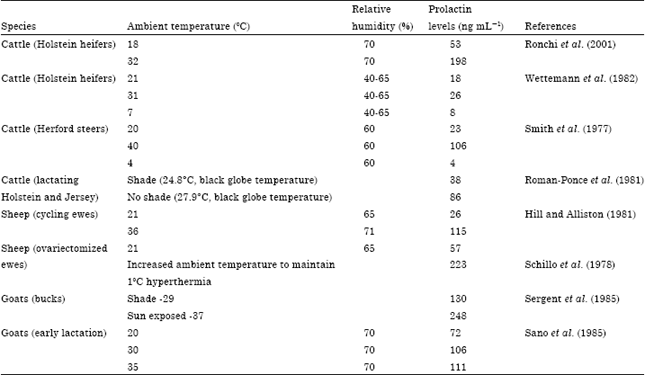 | |
| Table 2: | Effect of environmental temperature on lactotrophs sensitivity |
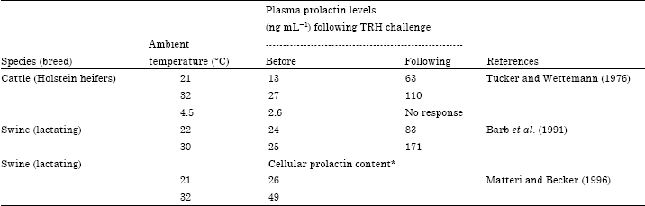 | |
| *ng per 250,000 cells | |
In summary, the positive correlation between ambient temperature and circulating prolactin values indicate that environmental temperature is a powerful stimulus to prolactin release.
The mechanism underlying prolactin stimulation under heat stress has been investigated in a number of species. Thermoregulatory responses to heat stress can be activated by peripheral or central thermoreceptors. However, in studies conducted on humans, it has been shown that peripheral thermoreceptors may not be involved in this process (Koska et al., 2003). Apparently, a substantial rise in core body temperature is a precondition for prolactin secretion (Mills and Robertshaw, 1981) pointing to the involvement of central thermoreceptors in prolactin response to heat exposure in humans. The modulation of circulating prolactin may trigger a suppression of prolactin inhibiting factor neurons and/or elicit of prolactin releasing factor neurons in the hypothalamus. A fall in the dopaminergic neurons activity with exposure to acute heat in calves was noted (Tucker et al., 1991) which was associated with greater prolactin release. Furthermore, it has been established in the ovine that dopamine Dl receptor antagonist substantially attenuated prolactin reaction to high ambient temperature (Colthorpe et al., 1998). This indicates that ventromedial hypothalamic nucleus Dl receptors are implicated in prolactin stimulatory pathway in response to high environmental temperature in the ovine.
It has been known that longer photoperiod stimulates prolactin secretion in several ruminants species (Mabjeesh et al., 2007; Mikolayunas et al., 2008; Garcia-Ispierto et al., 2009). However, a significant interaction between photoperiod and ambient temperature in the control of prolactin levels does exist. Several lines of evidence that indicate a marked fall in prolactin secretion with severe cold conditions in cattle when maintained on longer photoperiod (Zinn et al., 1986; Rius et al., 2005). Apparently, low ambient temperature may mask the stimulatory effect of extended photoperiod on prolactin profiles. Also, the prolactin response to temperature alteration is rapid compared to photoperiod which requires several weeks to reach a maximal prolactin concentrations (Tucker, 1982). This can lead to the conclusion that ambient temperature appears to be more important exteroceptive stimulus in prolactin release compared to photoperiod (Berardinelli et al., 1992).
Although various factors are known to influence the concentration of prolactin hormone measured in plasma, there is a paucity of findings to propose that the measured plasma prolactin is a function of the secretion rate of the pituitary and the clearance rate of the hormone from the blood. In cattle, it has been reported that the increase in plasma levels of prolactin associated with high ambient temperature exposure was a consequence of a reduction in metabolic clearance rate and an increase in secretion rate of this peptide hormone (Smith et al., 1977). Results from the same investigators indicate that the rate of prolactin secretion in the bovine is more important than the clearance rate in maintaining hyperprolactinemia at higher temperatures.
In summary, these findings indicate that prolactin is sensitive to the prevailing environmental temperature and this rise in prolactin values is likely to be implicated in the acclimation responses to thermal load. This homeorhetic process might be associated with adaptation of several physiological body functions that are connected with seasonal variation and heat stress such as thermoregulation and fluid balance (Beede and Collier, 1986; Bernabucci et al., 2010).
Prolactin as a stress hormone: Available data confirmed that prolactin release rises in response to various forms of physical and psychological stressors such as restraint or transport as well as heat stress (Van Vugt et al., 1978). Prolactin has been reported to be increased in response to stressful situations in various species such as camels (Kataria and Kataria, 2010b), donkeys (Kataria and Kataria, 2010a), cattle (Yayou et al., 2010) and sheep (Kataria and Kataria, 2011). It appears that the magnitude of prolactin response to stress depends on the type of stress. Correspondently, it can be argued that an observable prolactin response during heat exposure may be related to a non-specific stress response and may not be associated with thermoregulatory mechanisms. However, this argument is flawed for several reasons. First, not all stress conditions induce a rise in prolactin release; findings indicate that various stress conditions provoke a rather lessening in prolactin concentrations (Morehead and Gala, 1987, 1989). Second, employment of prolactin response as an index of stress has been questioned because it is practically impossible to measure and track the severity of stress by tracing prolactin profiles in the plasma (Natelson et al., 1988). Finally, a research evidence indicates that higher circulating prolactin during heat exposure cannot be interpreted by a nonspecific stress; but rather to specific to thermal stress (Mills and Robertshaw, 1981).
The modulation of thermoregulation by prolactin: The above survey of findings delineates a clear association between thermal stress and the hyper-secretion of prolactin. It is therefore reasonable to suggest that a rise in prolactin levels may be associated in some way to the regulation of body temperature. Furthermore, changes in prolactin secretion during high ambient temperature are positively associated with changes in body temperature. Comprehensive support for this concept comes from several studies that indicate a significant correlation between rectal temperature (RT)and peripheral prolactin concentration during thermal load in several species like goats (Chemineau and Ravault, 1984; Sergent et al., 1985), sheep (Schillo et al., 1978; Hill and Alliston, 1981) and humans (Melin et al., 1988; Low et al., 2005). It follows then that the secretion of prolactin represents part of the physiological response to alterations in ambient temperature.
The most common approach to evaluating the role of prolactin is to reduce it to minimal values by utilizing prolactin suppressants. Therefore, a further suggestion of the possible involvement of prolactin in thermoregulatory mechanisms is based on the observation that heat exposure combined with 2-bromo-α-ergocrytine treatment to suppress circulating prolactin in ovine result in an impairment of the thermoregulatory ability under mild (Faichney and Barry, 1986) or severe (Salah et al., 1995) heat stress conditions (Table 3). In these studies, an increase of 0.4-0.8°C in rectal temperature has been detected with prolactin suppression. Similar results have also been obtained in sun-exposed goats when prolactin suppression produced a thermoregulatory failure that resulted in hyperthermia during the hottest part of the day (Sergent et al., 1988). These findings might indicate that disruption of the prolactin rise in response to heat stress could indirectly affect some mechanisms that might be important in heat defense. Hence, higher prolactin levels during hot conditions stimulate physiological adjustments that enable an animal to tolerate the stress caused by a hot environment. In contrast, in study conducted on cattle, blocking the prolactin hyper-secretion following thermal burden did not result in a thermoregulatory dysfunction (Schams et al., 1980b). This inconsistency is likely the result of different procedures used for prolactin suppression. In the cattle study, only 4 injections of higher doses (0.38 mg/b.wt.) were administered over two week period, while in the other reported studies lower and more frequent doses at rates of 0.047-0.18 mg/bwt/d were given. The treated cattle also exhibited a rising trend in RT during prolactin suppression; however, the study was limited to only 3 treated bulls.
It can be concluded that thermoregulation failure associated with circulating prolactin decline suggests that higher prolactin values might support some thermoregulatory mechanisms during higher ambient temperature. However, the cause of this heat intolerance is not fully elucidated. In this context, cooled heat-stressed dairy cows maintained lower prolactin concentrations along with lower RT and respiration rates during heat stress compared to those without cooling relief (Igono et al., 1987; Do Amaral et al., 2011). Similarly, it has been observed in men that significantly lower prolactin levels in the plasma was induced by face cooling during exercise in hot environment (Brisson et al., 1989; Mundel et al., 2006). This might suggest a possible association of prolactin increase and the activation of heat defense mechanisms so that heat alleviation of heat stressed animals will decrease the needs for the enhancement of thermolysis mechanisms. Furthermore, Scharf et al. (2010) reported that heat exposure to heat sensitive steers, Angus, exhibited heat intolerance signs along with an elevation of serum prolactin value.
| Table 3: | Effect of prolactin suppression on thermoregulation in different ruminants' species |
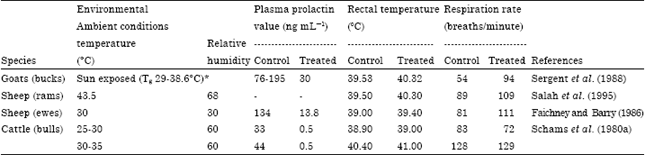 | |
| *Tg is black globe temperature | |
The same investigators, however, observed an absence of prolactin response to heat stress in the heat tolerant breed, Romosinuano. These results clearly demonstrate the association between the boost to dissipate excessive heat load and the rise in prolactin during heat exposure. The above discussion may further support the hypothesis of a potential role to play by prolactin in the thermoregulatory processes during heat exposure.
Maintenance of body temperature depends on the balance between heat input and output. During heat stress exposure, ruminants try to balance the excessive heat load by enhancing the different means of heat dissipation (Elnageeb et al., 2008; Umpapol et al., 2010). Physiological mechanisms for coping with heat stress include higher vasodilatation and increased blood flow to the skin surface along with enhancement in evaporative cooling. Accordingly, the thermoregulatory dysfunction associated with prolactin suppression during heat exposure may be related to a rise in heat storage and/or a down regulation in heat dissipation mechanisms. This cannot exclude the possibility of a disruption of the favorable internal environment for heat dissipation such as blood redistribution. In addition, whether higher prolactin levels influence the ability of ruminants to metabolically adapt during thermal stress is currently unknown. In ruminants there is an over-reliance on evaporative cooling for heat loss when environmental temperature reaches or exceeds skin temperature (Berman et al., 1985). Therefore, a disturbance in heat balance could be related to a possible diminution of evaporative cooling efficiency. However, in the above noted studies, the observed thermoregulatory failure resulting from prolactin suppression was not associated with an attenuation of respiratory evaporative heat loss (Sergent et al., 1988; Salah et al., 1995). Hence, this may point out to a possible down regulation in sweating. Furthermore, prolactin has been shown to stimulate the expression of heat shock protein-60 in rats (Stocco et al., 2001) and heat shock proteins are known to be involved in cytoprotection during heat stress and protect against hyperthermia (Ahmed, 2006; Venkatraman et al., 2006; Collier et al., 2008; Faisal et al., 2008).
The endocrine reaction is conditional on signal transduction through specific receptors and hence, the responsiveness to prolactin is regarded as a function of prolactin receptor expression in target tissues. It has been found that lower prolactin values induced by short day photoperiod (Auchtung et al., 2003) was shown to be associated with a greater expression of prolactin receptors mRNA in mammary gland of dairy cows. Furthermore, Do Amaral et al. (2011) revealed that a higher plasma levels of prolactin induced by heat stress in cows was associated with lower PRL-receptors mRNA expression by lymphocytes. Also, heat stress was associated with down regulation of prolactin signaling pathways in the liver and consequently hepatic lipid metabolism was impaired together with fat mobilization in heat-stressed cows compared to cooled ones (Do Amaral et al., 2009, 2011). Therefore, the stimulatory effect of thermal stress on prolactin secretion is associated with down-regulation of prolactin signaling pathways in a number of target tissues such the liver and the mammary gland. This could be partially accounted for the depressed milk production in dairy cows during heat stress. This can be regarded as an adaptation measure which increases the potential for survival in response to severe stress such as heat stress (Silanikove et al., 2000). Collectively, the presented findings so far may indicate that prolactin hyper-secretion during heat exposure may be involved in the enhancement of some thermoregulatory mechanisms, possibly by supporting the defense against heat or reducing heat increment.
Effect of prolactin on sweat glands: Since, the above cited studies of prolactin blocking during heat stress report normal or higher respiration activity, it is quite reasonable to assume that an impairment in sweating activity might be implicated in this heat balance failure. Prolactin has been proposed to be involved in the control of electrolyte and water flux in various fluid compartments in mammals such as gut, kidney and the mammary gland (Collier et al., 1982; Shennan, 1994). The involvement of prolactin in the regulation of ion transport in the sweat glands has been suggested which was based on the observation of gene expression of prolactin receptors in human sweat glands (Walker et al., 1989; Soos et al., 1993). In this context, evidence for the presence of specific prolactin binding sites in the ovine apocrine sweat glands (Choy et al., 1995) is therefore consistent with the role of prolactin in the modulation of sweat gland secretions.
Furthermore, some evidence in different species may suggest that higher prolactin values during heat exposure may modulate sweat gland activity. Bromocriptine treatment in men which abolishes the hyper-secretion of prolactin in response to exercise in warm conditions, significantly attenuates sweat secretion (Kaufman et al., 1988) with a concurrent increase in Na concentration in the sweat (Boisvert et al., 1993). Also, tall fescue toxicosis that is associated with the consumption of endophyte fungus Acremonium coenophialum is linked with signs of heat intolerance, summer toxicosis, such as elevated rectal temperature and respiration rate in ruminants (Hemken et al., 1981). Lower serum prolactin level is a common observation with fescue toxicosis which is found to be associated with a marked decrease in sweating rate in the sheep (Aldrich et al., 1993). Moreover, utilization of dopamine receptor antagonists in cattle with fescue toxicosis improves body temperature (Lipham et al., 1989) which might be related to increasing prolactin secretion. Therefore, it is possible to speculate that heat intolerance associated with fescue toxicosis could be partly ascribed to the negative effect of lower prolactin values on sweating activity. It is worth noting that in prolactin-suppressant ruminants, the respiratory frequency during thermal load increases compared to controls (Sergent et al., 1988; Salah et al., 1995). This can be viewed as an attempt by these animals to maintain homeothermy by dissipating surplus heat via enhancing respiration rate when the other avenue of evaporative cooling became insufficient. However, this apparent compensatory mechanism in heat elimination by pulmonary evaporation is not effective in preventing hyperthermia.
The role of prolactin in water balance: Heat stress induces marked alterations in water turnover rate and metabolism which boost the requirements for water during heat stress (Alamer, 2003, 2011). Hence, it has been suggested that prolactin response to heat exposure might be involved in meeting the expanding water demands of animals suffering from heat stress (Collier et al., 1982). Furthermore, due to the observed association between environmental temperature and circulating prolactin, it is assumed that prolactin might be associated with the control of water consumption. With the increased evaporative water loss during heat stress, animals increase their water intake. Water intake was found to be unchanged by lowering prolactin levels during heat exposure in small ruminants species (Sergent et al., 1988; Salah et al., 1995). However, in the study of Schams et al. (1980a) a lower water consumption has been detected with prolactin suppression in the bovine. In light of such conflicting findings, it may be suggested that no definite conclusion can be drawn regarding the involvement of prolactin in water intake regulation in ruminants. These inconsistent findings, however, might be related to the severity of heat load; greater thermal load would likely mask any deleterious effects on water intake brought about by prolactin suppression.
Heat stress induces a significant alteration in the body water balance and distribution. It has been reported that heat stress in ruminants results in the expansion of extracellular fluid compartment (El-Nouty et al., 1980; Koga et al., 1999; Alamer, 2011). Also, studies in rats (Horowitz et al., 1988; Meiri et al., 1991) and humans (Senay et al., 1976) demonstrate that plasma volume increases markedly under heat conditions. This is likely to be in proportion to the thermoregulatory necessity of the heat-stressed ruminant (Silanikove, 1987; Chaiyabtur et al., 1990; Silanikove, 1992) by creating advantageous condition for heat dissipation and, hence, buffer any rise in core body temperature.
An increase in plasma osmolality (POSM) and Na concentration have been observed in heat-stressed lambs that have been treated with bromocriptine (Salah et al., 1995). These increases could be regarded as signs of hypovolemia. Hyper-osmolality and hypovolemia are known to exert suppressive effects on evaporative cooling that might lead to hyperthermia (Senay, 1979; Moriomoto, 1990; Alamer and Al-hozab, 2004; Abdalla and Abdelatif, 2008). Therefore, this may offer a partially explanation for the effects of prolactin inhibition on thermoregulatory dysfunction. Salah et al. (1995) suggested that this inability to maintain plasma volume can be ascribed to a possible failure to move water from the gastrointestinal tract towards the blood compartment. In this context, ewes exposed to mild heat with prolactin suppression results in an inability to divert water from the gastrointestinal tract to the vascular system in response heat stress (Faichney and Barry, 1986). As a consequences this may alter the ability of the animals to control the extracellular fluid status. Such findings may indicate that prolactin plays an important role in the maintenance of fluid supply to the vascular system. Furthermore, there is evidence from some mammalian species that indicates the involvement of prolactin in fluid absorption in the intestinal epithelium (Mainoya et al., 1974; Shennan, 1994). Mainoya (1975) has demonstrated that prolactin increases fluid and electrolyte absorption in the duodenum and jejunum in rats. An earlier study (Mainoya et al., 1974) also demonstrated that prolactin has a stimulatory effect on in vitro fluid absorption in various parts of the intestine in rats, guinea-pigs and hamsters. In a later study, Mainoya (1979) showed that prolactin stimulates fluid and salt absorption in the proximal colon but not the distal colon, in rats. In this context, Mainoya (1981) subsequently found that prolactin suppression decreased colonic absorption of water and electrolyte in rats. The presented findings may suggest the influence of prolactin in water movement in the gut and therefore increasing fluid stores ensuring adequate fluid supply to the vascular compartment (Kaufman and Mackay, 1983).
Another role of prolactin in meeting the demand for water is that it may affect renal water loss and hence water conservation. Prolactin appears to reduce renal fluid and electrolyte excretion (Horrobin, 1980). However, very few studies have addressed the effect of prolactin on renal handling of water in ruminants. On the other hand, several studies in non-ruminant species have demonstrated that prolactin can influence renal excretion of water. Prolactin appears to reduce fluid, Na and K excretion in rats (Horrobin, 1980) while suppression of prolactin has been shown to increase urine volume and electrolyte excretion in rats (Richardson, 1973) and in humans (Cole et al., 1975). In rabbits, prolactin induces renal retention of water and salt without affecting water intake (Burstyn et al., 1975). One study in sheep suggests that prolactin can induce renal retention of water and Na but does so by stimulating the aldosterone effect (Burstyn et al., 1972). Collectively, these results suggest that elevated prolactin in heat stressed ruminant maybe involved in meeting the electrolyte and water requirements during thermal stress.
Heat stress induces a marked alteration in cardiac output and blood redistribution. Ruminants maintain their heat balance via vasomotor control by adjusting the amount of blood flowing through the cutaneous vessels by vasodilatation. Consequently, heat in the body core is transported to the body surfaces resulting in the increase of skin temperature (Hales, 1973; Al-Tamimi, 2005; Umpapol et al., 2010). Heat is then dissipated from the skin surface by means of evaporative water loss. The question that arises, then, is what role prolactin plays in this process? The relationship between prolactin levels during heat exposure and the rise in skin temperature (Low et al., 2005) may suggest an involvement of prolactin in blood redistribution or vasodilatation in peripheral tissues. Support for the contention that prolactin plays a role in blood redistribution to the periphery also arises from observed peripheral vasoconstriction when prolactin hypersecretion associated with exercise in heat is suppressed in humans (Brisson et al., 1989). In the case of fescue toxicosis, a decrease in blood flow to peripheral tissues associated with lower circulating prolactin has been noted (Rhodes et al., 1991), however, this cannot be ascribed solely to alteration in prolactin.
Based on observed prolactin modulation during dehydration, it appears that the hormone is also likely involved in body fluid regulation. Reduction of prolactin concentration in plasma was observed during dehydration caused by deprivation or restriction of water in cattle (Doris and Bell, 1984; Becker et al., 1985). However, contradicting results have been obtained in non-ruminant species. These results indicate that water deprivation has a stimulatory effect on prolactin secretion in rats (Kaufman and Mackay, 1983), chickens (Harvey et al., 1979) and in humans (Melin et al., 1988). Probably, such variation in prolactin response to dehydration resembles an example of species variation. In the case of ruminants, lower prolactin levels may be induced by a reduction of the volume of the extracellular fluid that developed during dehydration (Alamer, 2005, 2006). This also may indicates that prolactin suppression may modulate water preservation or body water redistribution mechanisms during periods of water deficiency.
Prolactin role in the control of pelage coat growth: Sleek and thinner hair coat contribute to preventing hyperthermia during heat exposure by facilitating better convective and conductive thermolysis together with a significant reduction in heat absorption by solar radiation (Finch et al., 1984). Consequently, cattle with slick hair coats experience lower body temperature during heat stress conditions (Dikmen et al., 2008). Seasonal pelage moult in ruminants, normally occurring during spring, in order to renew and modify the structure and composition of the pelage that is believed to be one component of adaptation to seasonal climatic changes. In this context, prolactin is believed to be involved in the control of seasonal pelage cycles in several species of mammals including domestic animals (Coffey et al., 2001; Foizik et al., 2009). This is also supported by the notation that the seasonal increase in prolactin levels is required for the growth of summer pelage (Martinet et al., 1984) and lower prolactin values can result in the delay of pelage shedding in ruminants (Gray et al., 2011).
Choy et al. (1995) provided evidence for the existence of prolactin receptors in a variety of cell types in the ovine skin and also the report of the abundance of high expressed and regulated prolactin receptors in follicle cell population known to play a key role in the control of pelage cycles (Nixon et al., 2002). Prolactin has been proposed to have a direct role in mediating pelage growth cycle and prolactin may act directly on the skin via cell components that have been known to exert some effects on the activity of fiber producing epithelium. In this regards, rough hair coat is one of the common symptoms exhibited by cattle with fescue toxicosis which is associated with a marked suppression in serum prolactin levels (Lipham et al., 1989; Coffey et al., 2001; Nihsen et al., 2004) instead of slick and smooth hair coats. Therefore, it has been proposed that reduced prolactin levels could be responsible for the deleterious effects of endophytic toxins on animals (Boling et al., 1989). This assumption gained support from reports indicating that utilization of dopamine receptors antagonists may ameliorate the negative effects of the endophytic toxins on prolactin related physiological functions such as hair coat characteristics (Boling et al., 1989; Lipham et al., 1989).
CONCLUSIONS
A mounting body of research indicates an association between prolactin and ambient temperature may reflect a possible influence of prolactin in thermoregulation. This connection has been further supported by the observed thermoregulatory failure that occurs when prolactin increases are inhibited during periods of thermal load. This indicates that prolactin elevation is regarded as an acclimatory response to heat stress. Several modulatory effects have been proposed which are related to thermoregulatory response. One is that prolactin may be involved in the maintenance of an advantageous internal environment to facilitate heat dissipation during heat exposure. Prolactin may also affects the maintenance of sustained fluid flow to the vascular system by facilitating fluid absorption from the gastrointestinal tract. Furthermore, changes in the expression of genes associated with prolactin signaling pathway in some tissues that may be consistent with mechanisms to down-regulate some metabolic processes and therefore support homeothemy. Finally, prolactin might exert some control on the appearance of the summer type hair coat to facilitate heat loss during hot conditions. Therefore, higher prolactin values during thermal load might influence the thermoregulatory mechanisms by facilitating heat loss and reducing heat increment directed to support homeothemy.
REFERENCES
- Abdalla, S.E. and A.M. Abdelatif, 2008. Effects of haemorrhage on thermoregulation, heart rate and blood constituents in goats (Capra hircus). Pak. J. Biol. Sci., 11: 1194-1203.
CrossRefPubMedDirect Link - Ahmed, R.G., 2006. The relation between biological consequences and high temperature in mammals. Int. J. Zool. Res., 2: 48-59.
CrossRefDirect Link - Al-Tamimi, H.J., 2005. Effects of solar radiation on thermophysiological and growth parameters of indigenous black Bedwin Goat kids in Southern Jordan. J. Biol. Sci., 5: 724-728.
CrossRefDirect Link - Alamer, M.A., 2003. Heat tolerance of local goat breed in Saudi Arabia. Arab Gulf J. Sci. Res., 21: 210-216.
Direct Link - Alamer, M. and A. Al-Hozab, 2004. Effect of water deprivation and season on feed intake, body weight and thermoregulation in Awassi and Najdi sheep breeds in Saudi Arabia. J. Arid Environ., 59: 71-84.
CrossRefDirect Link - Alamer, M., 2005. Effects of water deprivation and season on some biochemical constituents of blood in awassi and najdi sheep breeds in Saudi Arabia. J. Anim. Vet. Adv., 4: 107-117.
Direct Link - Alamer, M., 2011. Water requirements and body water distribution in Awassi sheep and Aardi goats during winter and summer seasons. J. Agric. Sci., 149: 227-234.
CrossRef - Aldrich, C.G., M.T. Rhodes, J.L. Miner, M.S. Kerley and J.A. Paterson, 1993. The effects of endophyte-infected tall fescue consumption and use of a dopamine antagonist on intake, digestability, body temperature and blood constituents in sheep. J. Anim. Sci., 71: 158-163.
Direct Link - Archawaranon, M., 2006. Seasonal changes in gonadotropins, prolactin and sex steroid hormones in captive hill mynah Gracula religiosa. J. Boil. Sci., 6: 375-381.
CrossRefDirect Link - Auchtung, T.L., P.E. Kendall, J.L. Salak-Johnson, T.B. McFadden and G.E. Dahl, 2003. Photoperiod and bromocriptine treatment effects on expression of prolactin receptor mRNA in bovine liver, mammary gland and peripheral blood lymphocytes. J. Endocrinol., 179: 347-356.
PubMed - Barb, C.R., M.J. Estienne, R.R. Kraeling, D.N. Marple, G.B. Rampacek, C.H. Rahe and J.L. Sartin, 1991. Endocrine changes in sows exposed to elevated ambient temperature during lactation. Domest. Anim. Endocrinol., 8: 117-127.
PubMed - Beede, D.K. and R.J. Collier, 1986. Potential nutritional strategies for intensively managed cattle during thermal stress. J. Anim. Sci., 62: 543-554.
Direct Link - Ben-Jonathan, N., E.R. Hugo, T.D. Brandebourq and C.R. LaPensee, 2006. Focus on prolactin as a metabolic hormone. Trends Endocrinol. Metab., 17: 110-116.
CrossRef - Bernabucci, U., N. Lacetera, L.H. Baumgard, R.P. Rhoads, B. Ronchi and A. Nardone, 2010. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal, 4: 1167-1183.
CrossRefDirect Link - Boisvert, P., G.R. Brisson and F. Peronnet, 1993. Effect of plasma prolactin on sweat rate and sweat composition during exercise in men. Am. J. Physiol., 264: F816-F820.
PubMed - Bole-Feysot, C., V. Goffin, M. Edery, N. Binart and P.A. Kelly, 1998. Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr. Rev., 19: 225-268.
PubMed - Boling, J.A., L.D. Bunting, G.M. Davenport, J.L. Van der Veen, K.M. Meekins, N.W. Bradley and R.E. Kohls, 1989. Physiological responses of cattle consuming tall fescue to environmental temperature and supplemental phenothiazine. J. Anim. Sci., 67: 2377-2385.
Direct Link - Brunet, A.G. and A.L. Sebastian, 1991. Effect of season on plasma concentrations of prolactin and cortisol in pregnant, non-pregnant and lactating ewes. Anim. Reprod. Sci., 26: 251-268.
Direct Link - Burstyn, P.G., D.F. Horrobin and M.S. Manku, 1972. Saluretic action of aldosterone in the presence of increased salt intake and restoration of normal action by prolactin or by oxytocin. J. Endocrinol., 55: 369-376.
PubMed - Burstyn, P.G., N. Gaynes, N. Golightly and A. Scott, 1975. Proceedings: The effects of prolactin on renal excretion and on voluntary intake of water and sodium in rabbits. J. Physiol., 251: 30P-31P.
PubMed - Chemineau, P. and J.P. Ravault, 1984. Hourly variations in rectal temperature and prolactinemia in the creole goat kept outside in a tropical environment. Reprod. Nutr. Dev., 24: 71-80.
PubMed - Choy, V.J., A.J. Nixon and A.J. Pearson, 1995. Localisation of receptors for prolactin in ovine skin. J. Endocrinol., 144:: 143-151.
PubMed - Coffey, K.P., W.K. Coblentz, J.B. Humphry, E.L. Piper and C.F. Rosenkrans et al., 2001. Growth performance and serum prolactin concentrations of stocker steers implanted with trenbolone acetate while grazing endophyte-infected fescue in the spring. Prof. Anim. Sci., 17: 166-173.
CrossRefDirect Link - Cole, E.N., D. Evered, D.F. Horrobin, M.S. Manku, J.P. Mtabaji and B.A. Nassar, 1975. Proceedings: Is prolactin a fluid and electrolyte regulating hormone in man? J. Physiol., 252: 54P-55P.
PubMed - Collier, R.J., D.K. Beede, W.W. Thatcher, L.A. Israel and C.J. Wilcox, 1982. Influences of environment and its modification on dairy animal health and production. J. Dairy Sci., 65: 2213-2227.
CrossRefDirect Link - Collier, R.J., J.L. Collier, R.P. Rhoads and L.H. Baumgard, 2008. Invited review: Genes involved in the bovine heat stress response. J. Dairy Sci., 91: 445-454.
CrossRefDirect Link - Colthorpe, K.L., S.T. Anderson, G.B. Martin and J.D. Curlewis, 1998. Hypothalamic dopamine D1 receptors are involved in the stimulation of prolactin secretion by high environmental temperature in the female sheep. J. Neuroendocrinol., 10: 503-509.
PubMed - Curlewis, J.D., 1992. Seasonal prolactin secretion and its role in seasonal reproduction: A review. Reprod. Fertil. Dev., 4: 1-23.
PubMedDirect Link - Dikmen, S., E. Alava, E. Pontes, J.M. Fear, B.Y. Dikmen, T.A. Olson and P.J. Hansen, 2008. Differences in thermoregulatory ability between slick-haired and wild-type lactating Holstein cows in response to acute heat stress. J. Dairy Sci., 91: 3395-3402.
CrossRef - Do Amaral, B.C., E.E. Connor, S. Tao, J. Hayen, J. Bubolz and G.E. Dahl, 2009. Heat-stress abatement during the dry period: Does cooling improve transition into lactation? J. Dairy Sci., 92: 5988-5999.
CrossRef - Do Amaral, B.C., E.E. Connor, S. Tao, M.J. Hayen, J.W. Bubolz and G.E. Dahl, 2011. Heat stress abatement during the dry period influence metabolic gene expression and improves immune status in the transition period of dairy cows. J. Dairy Sci., 95: 86-96.
CrossRefPubMedDirect Link - Doris, P.A. and F.R. Bell, 1984. Plasma prolactin during the body fluid and electrolyte changes of dehydration and sodium depletion in steers. Life Sci., 34: 1683-1689.
PubMed - El-Nouty, F.D., I.M. Elbanna, T.P. Davis and H.D. Johnson, 1980. Aldosterone and ADH response to heat and dehydration in cattle. J. Applied Physiol., 48: 249-255.
Direct Link - Elnageeb, M.E., A.M. Abdelatif and S.E.A. Makawi, 2008. Thermoregulation and reproductive performance of grazing desert ewes (Ovis aries) as influenced by concentrate supplementation. Pak. J. Biol. Sci., 11: 2209-2216.
CrossRefPubMedDirect Link - Eltayeb, N.M., C.E. Wani and I.A. Yousif, 2010. Assessment of broodiness and its influence on production performance and plasma prolactin level in native chicken of the Sudan. Asian J. Poult. Sci., 4: 1-6.
CrossRefDirect Link - Faichney, G.J. and T.N. Barry, 1986. Effects of mild heat exposure and suppression of prolactin secretion on gastro-intestinal tract function and temperature regulation in sheep. Aust. Biol. Sci., 39: 85-97.
PubMed - Faisal, B.A., S.A. Abdel-Fattah, Y.M. El-Hommosany, M.F.M. Ali and N.M. Abdel-Gawad, 2008. Immunocompetence, hepatic heat shock protein 70 and physiological responses to feed restriction and heat stress in two body weight lines of Japanese quail. Int. J. Poult. Sci., 7: 174-183.
CrossRefDirect Link - Finch, V.A., I.L. Bennett and C.R. Holmes, 1984. Coat colour in cattle: Effect on thermal balance, behaviour and growth and relationship with coat type. J. Agric. Sci., 102: 141-147.
CrossRef - Foizik, K., E.A. Langan and R. Paus, 2009. Prolactin and the skin: A dermatological perspective on an ancient pleiotropic peptide hormone. J. Invest. Dermatol., 129: 1071-1087.
CrossRef - Freeman, M.E., B. Kanyicska, A. Lerant and G. Nagy, 2000. Prolactin: Structure, function and regulation of secretion. Physiol. Rev., 80: 1523-1631.
PubMed - Garcia-Ispierto, I., F. Lopez-Gatius, S. Almeria, J. Yaniz and P. Santolaria et al., 2009. Factors affecting plasma prolactin concentrations throughout gestation in high producing dairy cows. Domestic Anim. Endocrinol., 36: 57-66.
CrossRefDirect Link - Hales, J.R., 1973. Effects of exposure to hot environments on the regional distribution of blood flow and on cardiorespiratory function in sheep. Pflugers Arch., 344: 133-148.
PubMed - Harvey, S., C.G. Scanes, A. Chadwick and N.J. Bolton, 1979. Growth hormone and prolactin secretion in growing domestic fowl: Influence of sex and breed. Br. Poult. Sci., 20: 9-17.
PubMed - Hemken, R.W., J.A. Boling, L.S. Bull, R.H. Hatton, R.C. Buckner and L.P. Bush, 1981. Interaction of environmental temperature and anti-quality factors on the severity of summer fescue toxicosis. J. Anim. Sci., 52: 710-714.
PubMed - Hill, T.G. and C.V. Alliston, 1981. Effects of thermal stress on plasma concentrations of luteinizing hormone, progesterone, prolactin and testosterone in the cycling ewe. Theriogenology, 15: 201-209.
PubMed - Horowitz, M., E. Sugimoto, T. Okuno and T. Morimoto, 1988. Changes in blood volume and vascular compliance during body heating in rats. Pflugers Archiv. Eur. J. Physiol., 412: 354-358.
CrossRef - Horrobin, D.F., 1980. Prolactin as a regulator of fluid and electrolyte metabolism in mammals. Fed. Proc., 39: 2567-2570.
PubMed - Igono, M.O., H.D. Johnson, B.J. Steevens, G.F. Krause and M.D. Shanklin, 1987. Physiological, productive and economic benefits of shade, spry and fan system versus shade for Holstein cows during summer heat. J. Dairy Sci., 70: 1069-1079.
CrossRef - Karsch, F.J., J.E. Robinson, C.J.I. Woodfill and M.B. Brown, 1989. Circannual cycles of luteinizing hormone and prolactin secretion in ewes during prolonged exposure to a fixed photoperiod: Evidence for an endogenous reproductive rhythm. Biol. Reprod., 41: 1034-1046.
PubMed - Kataria, N. and A.K. Kataria, 2010. Assessment of stress due to hot ambience in donkeys from arid tracts in India. J. Stress Physiol. Biochem., 6: 12-17.
Direct Link - Kataria, N. and A.K. Kataria, 2011. Use of prolactin as an indicator of stress in Marwari sheep from arid tracts in India. Comp. Clin. Pathol., 20: 333-336.
CrossRefDirect Link - Kaufman, F.L., D.E. Mills, R.L. Hughson and G.T. Peake, 1988. Effects of bromocriptine on sweat gland function during heat acclimatization. Horm. Res., 29: 31-38.
PubMed - Kaufman, S. and B.J. Mackay, 1983. Plasma prolactin levels and body fluid deficits in the rat: Causal interactions and control of water intake. J. Physiol., 336: 73-81.
Direct Link - Koga, A., K. Kurrata, K. Ohata, M. Nakajima and H. Hirose et al. 1999. Internal changes of blood compartment and heat distribution in swamp buffaloes under hot conditions: Comparative study of thermo-regulation in buffaloes and Friesian cows. Asian-Aus. J. Anim. Sci., 12: 886-890.
Direct Link - Koska, J., J. Rovensky, T. Zimanova and M. Vigas, 2003. Growth hormone and prolactin responses during partial and whole body warm-water immersions. Acta Physiol. Scand., 178: 19-23.
CrossRef - Lipham, L.B., F.N. Thompson, J.A. Stuedmann and J.L. Strin, 1989. Effects of metoclopramide on steers grazing endophyte-infected fescue. J. Anim. Sci., 67: 1090-1097.
PubMed - Low, D., T. Cable and A. Purvis, 2005. Exercise thermoregulation and hyperprolactinaemia. Ergonomics, 48: 1547-1557.
PubMed - Mabjeesh, S.J., O.G. Garber and A. Shamay, 2007. Effect of photoperiod in the third trimester of gestation on milk production and circulating hormones in dairy goats. J. Dairy Sci., 90: 699-705.
Direct Link - Mainoya, J.R., 1975. Effects of bovine growth hormone, human placental lactogen and ovineprolactin on intestinal fluid and ion transport in the rat. Endocrinology, 96: 1165-1170.
PubMed - Mainoya, J.R., 1979. Effect of prolactin on fluid and NaCl absorption by the rat proximal and distal colon. Experientia, 35: 1060-1061.
PubMed - Mainoya, J.R., 1981. Colon absorption of water and NaCl in the rat during lactation and the possible involvement of prolactin. Experientia, 37: 1083-1084.
CrossRef - Mainoya, J.R., H.A. Bern and J.W. Regan, 1974. Influence of ovine prolactin on transport of fluid and sodium chloride by the mammalian intestine and gall bladder. J. Endocrinol., 63: 311-317.
PubMed - Matteri, R.L., B.A. Becker and W.R. Lamberson, 1994. Somatotroph and lactotroph function in relation to growth in sex-week-old pigs reared in a hot or cool environment. Domest. Anim. Endocrinol., 11: 101-114.
PubMed - Matteri, R.L. and B.A. Becker, 1996. Lactotroph and somatotroph function in piglets reared in a constant hot environment. Life Sci., 58: 711-717.
PubMed - Martinet, L., D. Allain and C. Weiner, 1984. Role of prolactin in the photoperiodic control of moulting in the mink (Mustela vison). J. Endocrinol., 103: 9-15.
PubMed - Melin, B., M. Cure, J.M. Pequignot and J. Bittel, 1988. Body temperature and plasma prolactin and norepinephrine relationships during exercise in a warm environment: Effect of dehydration. Eur. J. Appl. Physiol. Occup. Physiol., 58: 146-151.
PubMed - Mikolayunas, C.M., D.L. Thomas, G.E. Dahl, T.F. Gressley and Y.M. Berger, 2008. Effect of prepartum photoperiod on milk production and prolactin concentration of dairy ewes. J. Dairy Sci., 91: 85-90.
CrossRef - Mills, D.E. and D. Robertshaw, 1981. Response of plasma prolactin to changes in ambient temperature and humidity in man. J. Clin. Endocrinol. Metab., 52: 279-283.
PubMed - Morehead, M.H. and R.R. Gala, 1987. Restraint stress depresses prolactin surges in pseudopregnant rats and adrenalectomy does not alter the response. Life Sci., 41: 1491-1498.
CrossRef - Morehead, M.H. and R.R. Gala, 1989. The restraint stress-induced decrease of the nocturnal prolactin surge and the physiology of pseudopregnancy and pregnancy in the rat. Life Sci., 45: 207-215.
PubMed - Moriomoto, T., 1990. Thermoregulation and body fluids: Role of blood volume and central venous pressure. Jap. J. Physiol., 40: 165-179.
PubMed - Mundel, T., P. Hooper, S.J. Bunn and D.A. Jones, 2006. The effects of face cooling on the prolactin response and subjective comfort during moderate passive heating in humans. Exp. Physiol., 91: 1007-1014.
CrossRef - Natelson, B.H., H.E. Ottenweller, D. Pitman and W.N. Tapp, 1988. An assessment of prolactin's value as an index of stress. Life Sci., 42: 1597-1602.
PubMedDirect Link - Nihsen, M.E., E.L. Piper, C.P. West, R.J. Jr. Crawford and T.M. Denard et al., 2004. Growth rate and physiology of steers grazing tall fescue inoculated with novel endophytes. J. Anim. Sci., 82: 878-883.
Direct Link - Nixon, A.J., C.A. Ford, J.E. Wildermoth, A.J. Craven, M.G. Ashby and A.J. Pearson, 2002. Regulation of prolactin receptor expression in ovine skin in relation to circulating prolactin and wool follicle growth status. J. Endocrinol., 172: 605-614.
PubMedDirect Link - Rhodes, M.T., J.A. Paterson, M.S. Kerley, H.E. Garner and M.H. Laughlin, 1991. Reduced blood flow to peripheral and core body tissues in sheep and cattle induced by endophyte-infected tall fescue. J. Anim. Sci., 69: 2033-2043.
Direct Link - Richardson, B.P., 1973. Evidence for a physiological role of prolactin in osmoregulation in the rat after its inhibition by 2-bromo-α-ergokrytine. Br. J. Pharmacol., 47: 623P-624P.
Direct Link - Rius, A.G., E.E. Connor, A.V. Capuco, P.E. Kendall, T.L. Auchtung-Montgomery and G.E. Dahl, 2005. Long-day photoperiod that enhances puberty does not limit body growth in holstein heifers. J. Dairy Sci., 88: 4356-4365.
Direct Link - Roman-Ponce, H., W.W. Thatcher, R.J. Collier and C.J. Wilcox, 1981. Hormonal responses of lactating dairy cattle to TRH and ACTH in a shade management system within subtropical environment. Theriogenology, 16: 131-138.
Direct Link - Ronchi, B., G. Stradaioli, A.V. Supplizi, U. Bernabucci and N. Lacetera et al., 2001. Influence of heat stress or feed restriction on plasma progesterone, oestradiol-17β, LH, FSH, prolactin and cortisol in Holstein heifers. Livest. Prod. Sci., 68: 231-241.
CrossRefDirect Link - Roy, K.S. and B.S. Prakash, 2007. Seasonal variation and circadian rhythmicity of the prolactin profile during the summer months in repeat-breeding Murrah buffalo heifers. Reprod. Fertil. Dev., 19: 596-605.
PubMed - Salah, M.S., M.A. AlShaikh, M.Y. Al-Saiadi and H.H. Mogawer, 1995. Effect of prolactin inhibition on thermoregulation, water and food intakes in heat-stressed fat-tailed male lambs. Anim. Sci., 60: 87-91.
Direct Link - Schams, D., E. Stephan and R.D. Hooley, 1980. Effect of prolactin inhibition under heat exposure on water intake and excretion of urine, sodium and potassium in bulls. Acta Endocrinol., 94: 315-320.
Direct Link - Schams, D., E. Stephan and R.D. Hooley, 1980. The effect of heat exposure on blood serum levels of anterior pituitary hormones in calves, heifers and bulls. Acta Endocrinol., 94: 309-314.
Direct Link - Scharf, B., J.A. Carroll, D.G. Riley, C.C. Jr. Chase and S.W. Coleman et al., 2010. Evaluation of physiological and blood serum differences in heat-tolerant (Romosinuano) and heat-susceptible (Angus) Bos taurus cattle during controlled heat challenge. J. Anim. Sci., 88: 2321-2336.
CrossRef - Schillo, K.K., C.W. Alliston and P.V. Malven, 1978. Plasma concentrations of luteinizing hormone and prolactin in the ovariectomized ewe during induced hyperthermia. Biol. Reprod., 19: 306-313.
Direct Link - Senay, Jr. L.C., 1979. Temperature regulation and hypohydration: A singular view. J. Applied Physiol., 47: 1-7.
Direct Link - Senay, L.C., D. Mitchell and C.H. Wyndham, 1976. Acclimatization in a hot, humid environment: body fluid adjustments. J. Applied Physiol., 40: 786-796.
Direct Link - Sergent, D., P. Berbigier, G. Kann and J. Fevre, 1985. The effect of sudden solar exposure on thermophysiological parameters and on plasma prolactin and cortisol concentrations in male Creole goats. Reprod. Nutr. Dev., 25: 629-640.
Direct Link - Sergent, D., P. Berbigier and J.P. Ravault, 1988. Effect of prolactin inhibition on thermophysiological parameters, water and feed intake of sun-exposed male Creole goats (Capra hircus) in Guadeloupe (French West Indies). J. Therm. Biol., 13: 53-59.
CrossRef - Shennan, D., 1994. Regulation of water and solute transport across mammalian plasma cell membranes by prolactin. Dairy Res., 61: 155-166.
PubMed - Silanikove, N., 1987. Impact of shelter in hot Mediterranean climate on feed intake, feed utilization and body fluid distribution in sheep. Appetite, 9: 207-215.
CrossRefDirect Link - Silanikove, N., 1992. Effects of water scarcity and hot environment on appetite and digestion in ruminants: A review. Livestock Prod. Sci., 30: 175-194.
CrossRef - Smith, V.G., R.R. Hacker and R.G. Brown, 1977. Effect of alterations in ambient temperature on serum prolactin concentration in steers. J. Anim. Sci., 44: 645-649.
CrossRefPubMedDirect Link - Stocco, C., E. Callegari and G. Gibori, 2001. Opposite effect of prolactin and prostaglandin F2α on the expression of luteal genes as revealed by rat cDNA expression array. Endocrinology, 142: 4158-4161.
Direct Link - Tucker, H.A., L.T. Chapin, K.J. Lookingland, K.E. Moore, G.E. Dahl and J.M. Evers, 1991. Temperature effects on serum prolactin concentrations and activity of dopaminergic neurons in the infundibulum/pituitary stalk of calves. Proc. Soc. Exp. Biol. Med., 197: 74-76.
PubMedDirect Link - Tucker, H.A. and R.P. Wettemann, 1976. Effects of ambient temperature and relative humidity on serum prolactin and growth in heifers. Proc. Soc. Exp. Biol. Med., 151: 623-626.
PubMed - Umpapol, H., C. Vajrabukka, T. Jitrajug, C. Songvicha, S. Tainglum and W. Sirikul, 2010. Application of GnRH administration at post artificial insemination in synchronized estrus heifer and dairy cows by PGF2α induction on conception rate in phupan dairy co-operative, Sakol-Nakon Province, Thailand. Pak. J. Nutr., 9: 594-599.
CrossRefDirect Link - Venkatraman, A., N.S.N. Kumar and B.S. Ramakrishna, 2006. Butyrate cytoprotection of colonic epithelial cells may be mediated through inhibition of heat shock protein 70. Asian J. Cell Biol., 1: 81-92.
CrossRefDirect Link - Walker, A.M., M.T. Robertson and C.J. Christopher, 1989. Distribution of a prolactinlike material in human eccrine sweat glands. J. Invest. Dermatol., 93: 50-53.
CrossRefDirect Link - Wettemann, R.P. and H.A. Tucker, 1976. The influence of low and elevated ambient temperatures on serum prolactin and growth hormone in heifers-A review. Int. J. Biometeorol., 20: 36-41.
CrossRef - Wettermann, R.P., H.A. Tucker, T.W. Beck and D.C. Meyerhoeffer, 1982. Influence of ambient temperature on prolactin concentrations in serum of holstein and brahman x hereford heifers. J. Anim. Sci., 55: 391-394.
PubMedDirect Link - Yayou, K.I., S. Ito, N. Yamamoto, S. Kitagawa and H. Okamura, 2010. Relationships of stress responses with plasma oxytocin and prolactin in heifer calves. Physiol. Behav., 99: 362-369.
PubMed - Zinn, S.A., R.W. Purchas, L.T. Chapin, D. Petitclerc, R.A. Merkel, W.G. Bergen and H.A. Tucker, 1986. Effects of photoperiod on growth, carcass composition, prolactin, growth hormone and cortisol in prepubertal and postpubertal Holstein heifers. J. Anim. Sci., 63: 1804-1815.
PubMedDirect Link - Meiri, U., M. Shochina and M. Horowitz, 1991. Heat acclimated hypohydrated rats: Age-dependent vasomotor and plasma volume responses to heat stress. J. Therm. Biol., 16: 241-247.
CrossRef