
Yan-Ping Wang
Laboratory of Animal Embryonic Biotechnology, College of Animal Science and Technology and State Key Laboratories for Agrobiotechnology, China Agricultural University, Beijing, 100193, Peoples� Republic of China
Guang-Bin Zhou
Institute of Animal Genetics and Breeding, College of Animal Science and Technology, Sichuan Agricultural University (Chengdu Campus), Wenjiang, 611130, Peoples� Republic of China
Yan Zeng
Laboratory of Animal Embryonic Biotechnology, College of Animal Science and Technology and State Key Laboratories for Agrobiotechnology, China Agricultural University, Beijing, 100193, Peoples� Republic of China
Jun-Jie Li
Laboratory of Animal Embryonic Biotechnology, College of Animal Science and Technology and State Key Laboratories for Agrobiotechnology, China Agricultural University, Beijing, 100193, Peoples� Republic of China
Qing-Jing Zhang
Laboratory of Animal Embryonic Biotechnology, College of Animal Science and Technology and State Key Laboratories for Agrobiotechnology, China Agricultural University, Beijing, 100193, Peoples� Republic of China
Yun-Peng Hou
Laboratory of Animal Embryonic Biotechnology, College of Animal Science and Technology and State Key Laboratories for Agrobiotechnology, China Agricultural University, Beijing, 100193, Peoples� Republic of China
Shi-En Zhu
Laboratory of Animal Embryonic Biotechnology, College of Animal Science and Technology and State Key Laboratories for Agrobiotechnology, China Agricultural University, Beijing, 100193, Peoples� Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 11 | Page No.: 1052-1060







ABSTRACT
Hyperactivated motility of sperm cells is essential to successful fertilization. This study was performed to observe the effect of Thimerosal treatment of cryopreserved mouse sperm on their hyperactivated motility and subsequent development to two-cell embryos after in vitro fertilization. Hyperactivated motility was evaluated by curvilinear velocity (VCL, the rate of travel of the sperm head) and Amplitude of Lateral Head Displacement (ALH, degree of side-to-side head movement measured as the mean width of head oscillations) of cells using Computer-assisted Sperm Analysis (CASA). The fresh sperm had higher VCL and ALH than frozen-thawed sperm (p<0.05). Following In Vitro Fertilization (IVF), fertilization rate from frozen-thawed sperm was significantly lower than that from fresh sperm (71.6, 94.4%) (p<0.05). Neither VCL nor ALH differed significantly between fresh sperm and frozen-thawed sperm not treated with Thimerosal. The decreased hyperactivated motility of mouse sperm due to cryopreservation may contribute to reduced fertilization rate compared to that achieved with fresh sperm. In addition because Thimerosal treatment of sperm after cryopreservation enhances hyperactivated motility, its use may improve the development of cryopreserved mouse oocytes from cryopreserved sperm.
PDF Abstract XML References Citation
Received: April 14, 2011;
Accepted: August 15, 2011;
Published: September 28, 2011
How to cite this article
Yan-Ping Wang, Guang-Bin Zhou, Yan Zeng, Jun-Jie Li, Qing-Jing Zhang, Yun-Peng Hou and Shi-En Zhu, 2011. Impact on Hyperactivated Motility of Cryopreserved Mouse Sperm from Pretreatment with Thimerosal. Asian Journal of Animal and Veterinary Advances, 6: 1052-1060.
DOI: 10.3923/ajava.2011.1052.1060
URL: https://scialert.net/abstract/?doi=ajava.2011.1052.1060
DOI: 10.3923/ajava.2011.1052.1060
URL: https://scialert.net/abstract/?doi=ajava.2011.1052.1060
INTRODUCTION
Hyperactivated motility is a movement pattern observed in spermatozoa at the site and time of fertilization in mammals (Ho and Suarez, 2001). It is characterized by high-amplitude and asymmetrical flagellar beating of sperm cells which assist them in penetrating the oocyte zona pellucid (Yanagimachi, 1994). In the mouse, hyperactivated sperm can be observed through the wall of the oviduct (Suarez and Pacey, 2006; Abou-Haila and Tulsiani, 2009), pulling off attachments to the mucosal epithelium and rapidly escaping from peripheral storage sites into the central lumen (Suarez and Ho, 2003; Rubinstein et al., 2006; Eisenbach and Giojalas, 2006).
Ca2+ signaling in sperm is critical for successful hyperactivated sperm motility. On the one hand, Ca2+ acts through calmodulin on the axoneme to modulate flagellar curvature (Ho et al., 2002); whereas on the other hand, Ca2+ influx may regulate ATP homeostasis (Xia et al., 2007). Two mechanisms trigger the rise of intracellular Ca2+ to initiate and maintain hyperactivation: increased Ca2+ entry via four plasma membrane CatSper ion channels (Qi et al., 2007) and release of Ca2+ from an inositol 1,4,5-trisphosphate (IP3)-gated internal store at the base of the flagellum (Ho and Suarez, 2001). These CATSPER proteins are from alkaline-sensitive voltage-gated and Ca2+ channels are located on the principal piece of the flagellum (Ren et al., 2001). Four CATSPER (Caspter1, Caspter2, Caspter3, Caspter4) channels have clearly been shown to be required for male fertility in that male mice deficient in any one of the four CATSPERs are completely sterile but exhibit no other apparent abnormalities (Ren et al., 2001; Quill et al., 2001, 2003; Kirichok et al., 2006; Jin et al., 2007). IP3 receptors localized to a portion of the redundant nuclear envelope on the side of the flagellum have been shown to increase bend amplitude in response to Ca2+ (Ho and Suarez, 2003). It is evident that release of internal Ca2+ stores was sufficient to initiate hyperactivation in wild-type sperm, similar to results from bull sperm (Ho and Suarez, 2001).
Successful cryopreservation of semen is important not only for the preservation of genetic resources but also for artificial insemination (Salamon and Maxwell, 2000; Dumpala et al., 2006; Sundararaman and Edwin, 2008; Shakeri et al., 2008). Although much progress has been made since this technique was introduced in 1949 (Polge et al., 1949), cryopreservation of mammalian sperm remains more difficult than cryopreservation of diploid cells. Previous studies have shown that cryopreservation induces ultrastructural damage to plasma (Gillan et al., 1995) and to acrosomal membranes (Purdy, 2006; Saad et al., 2011) and that it lowers mitochondrial membrane potential (Pena et al., 2009; Nauk, 1991). However, systematic investigations of hyperactivated sperm motility following cryopreservation have not been reported.
This study has been designed to observe: (1) hyperactivation motility and fertilization rate from frozen-thawed sperm and (2) the effect of Thimerosal, Ca2+ or procaine pretreatment on hyperactivation motility.
MATERIALS AND METHODS
This research project was conducted from February 2009 to March 2010 in Beijing. All chemicals and media were purchased from Sigma Chemical Co. (St. Louis, MO) unless otherwise indicated.
Animals: Kunming (KM) mice (Academy of Military Medical Sciences, Beijing, China) were used in the present study. Mice were housed under temperature- (20-22°C) and light-controlled conditions (lights-on, 06:00-20:00) with free access to food and water. Semen samples were collected from 50 mature males weighing between 35 and 40 g and ranging between 10 and 15 week of age.
Solutions for freezing sperm: Cryopreservation medium contained 18% (w/v) raffinose and 3% (w/v) skim milk (Nakagata, 2000).
Sperm freezing and thawing: Fresh sperm: Caudal epididymides were excised and rinsed with HS medium (Wennemuth et al., 2000):
| • | Freezing sperm: Sperm were loaded into 0.25 mL straws (I.V.M., L’Aigle, France) with cryopreservation solution. After equilibrating for 30 min, straws were placed 2 cm above liquid nitrogen (LN2) for 10 min, then plunged into LN2 and stored for at least 1 week |
| • | Thawing: Frozen straws were removed from liquid nitrogen and put into a water bath maintained at 37°C for no longer than 15 min. The spermatozoa suspensions were then added to 75 μL of HTF medium with a final concentration of approximately 5x105 cells mL-1 and incubated for 1-2 h at 37°C |
Partial zona pellucida incised by a piezo manipulator (ZIP) of oocytes: Procedures for ZIP were performed as described by Kawase et al. (2002). After ZIP, the oocytes were cultured in HTF medium for at least 1 h before IVF.
In vitro fertilization (IVF): Oocytes were placed into 75 μL drops of HTF medium under mineral oil, in which a small drop of spermatozoa suspension was added for insemination. Five hours after IVF, eggs were removed from the fertilization drop, washed in HTF medium and cultured in 50 μL drops of HTF medium.
About 24 h after fertilization, oocytes were examined and those that had cleaved into two equal blastomeres were considered to be fertilized.
Analysis of sperm motility: Sperm motility was also evaluated using Computer-assisted Sperm Analysis (CASA) (Sundararaman et al., 2007). Sperm movement was imaged using a 4xOlympus negative phase objective (Hi-Tech Instruments, Philadelphia, PA) and recorded using a Panasonic AG-7300 Super VHS video recorder. The video images were digitized (30 frames at 60 Hz) and analyzed using HTM-IVOS (Version 12.3, Hamilton Thorne Research, Beverly, MA). Motion parameters measured were curvilinear velocity (VCL, the rate of travel of the sperm head), amplitude of lateral head displacement (ALH, degree of side-to-side head movement measured as the mean width of head oscillations) and beat/cross frequency (BCF, number of times the head crossed the averaged path per second). Because movement of the sperm head is dependent on flagellar activity, it can be used as an indicator of flagellar bending patterns. Increased VCL and ALH are indicative of hyperactivation (Mortimer and Mortimer, 1990). For each treatment sample, 400-450 motile sperm were analyzed and for each experiment, replicate tests were performed.
Ca2+ imaging: Sperm were loaded with 10 μM Fluo-3 AM and 0.05% pluronic F-127 for 30 min at room temperature in the dark, followed by 2 washes in HS medium, each with a 5 min spin at 400 g. Washed sperm were resuspended in HS medium, plated onto cover slips coated with Cell Tak and allowed to attach for approximately 10 min.
Fluorescence intensities were quantified using EZ-C1 Free Viewer software (Nikon), as described previously (Aoki et al., 1997) with some modifications.
The treatment of sperm: Fresh and cryopreservation sperm were exposed to 50 μM Thimerosal, 2.4 mM Ca2+, 5 mM Procaine or 10 mM 1,2-bis (2-aminophenoxy) ethane-N,N, N9, N9-tetraacetic acid (BAPTA), after which hyperactivated motility was measured using Computer-assisted Sperm Analysis (CASA) as previously described.
Statistic analyses: Data were analyzed using Minitab statistical software (Minitab Inc., State College, PA). Treatment effects were detected using analysis of variance, followed by Tukey's test for individual post-hoc comparisons. Differences at p<0.05 were considered statistically significant.
RESULTS
Effect of ZIP treatment on the fertilization rates of mouse oocytes after IVF with cryoprserved spermatozoa: According to Table 1, the cleavage rate (71.6%) of oocytes fertilized by post-thawed spermatozoa was significantly lower than that of fresh oocytes fertilized by fresh sperm (94.4%) and of ZIP (Partial zona pellucida incised by a piezo manipulator)-zona oocytes fertilized with either fresh (94.5%) or frozen sperm (89.3%) (p<0.05).
Computer-assisted sperm analysis (CASA) measurements of post-thawed spermatozoa treated with thimerosal: As shown in Table 2, the VCL and ALH in post-thawed sperm were lower than in fresh sperm (301.5 vs.216.0, 16.2 vs.10.4, p<0.05). There was significantly higher in VCL or ALH between post-thawed sperm exposed to thimerosal and that of those without Thimerosal treatment (277.6 vs. 216.0, 15.9 vs 10.4, p<0.05). It is similar to the situation with fresh sperm (357.2 vs 301.5. 21.3 vs. 16.2, p<0.05). Stimulation by thimerosal of post-thawed sperm was not blocked by lowering extracellular Ca2+ with 10 mM 1,2-bis (2-aminophenoxy) ethane-N,N, N9, N9-tetraacetic acid (BAPTA) to approximately 30 nM.
Computer-assisted sperm analysis (CASA) measurements of post-thawed spermatozoa treated with Ca2+: Addition of 2.4 mM Ca2+ increased the VCL and ALH in post-thawed sperm and the VCL were higher (373.5, p<0.05) than that in non-treated fresh sperm (314.6). But there is a significant difference in ALH between the post-thawed sperm exposed to Ca2+ and the fresh sperm (13.8 vs.15.3, p<0.05) (Table 3).
Casa measurements of post-thawed spermatozoa treated with procaine: Exposure of post-thawed sperm to Procaine visibly increased the fresh sperm hyperactivated motility as measured by VCL (308.7 vs. 358.3, p<0.05) but the VCL and ALH were significantly lower (p<0.05) in post-thawed sperm than in fresh sperm irregardless of whether they were treated with procaine (230.1 vs.308.7, 264.8 vs. 308.7; 11.3 vs. 15.9, 12.4 vs.15.9) (Table 4).
| Table 1: | Effect of ZIP treatment on the fertilization rates of mouse oocytes after IVF with cryopreserved spermatozoa |
 | |
| ZIP: Partial zona pellucida incised by a piezo manipulator, IVF: In vitro fertilization, Values with different superscripts within the same column differ significantly (p<0.05) | |
| Table 2: | CASA measurements of post-thawed spermatozoa treated with thimerosal |
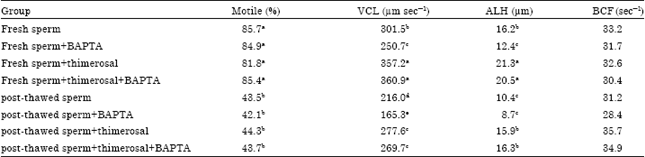 | |
| Values with different superscripts within the same column differ significantly (p<0.05). VCL: The rate of travel of the sperm head, ALH: Amplitude of lateral head displacement, degree of side-to-side head movement measured as the mean width of head oscillations, BCF: Beat/cross frequency, No. of times the head crossed the averaged path per second, CASA: Computer-assisted sperm analysis | |
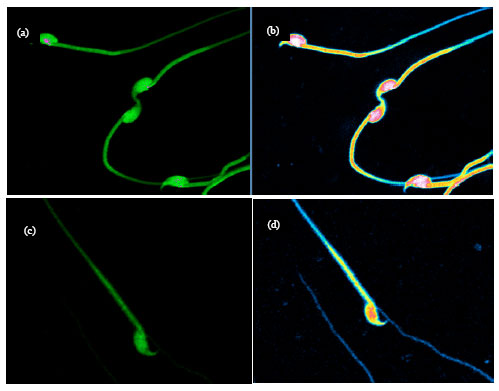 | |
| Fig. 1(a-d): | Ca2+ imaging in the cryopreserved sperm. Ca2+ (green) were found to be distributed at the head and the flagellar in the mouse fresh sperm (Fig. 1a, b) and their color turned cooler (Fig. 1c, d) when the sperm were cryopreserved |
| Table 3: | CASA measurements of post-thawed spermatozoa treated with Ca2+ |
 | |
| Values with different superscripts within the same column differ significantly (p<0.05) | |
| Table 4: | CASA measurements of post-thawed spermatozoa treated with Procaine |
 | |
| Values with different superscripts within the same column differ significantly (p<0.05) | |
Effect of cryopreservation on Ca2+ concentration in mouse sperm: In the present study, the changes of Ca2+ imaging of fluo3-loaded sperm were also examined. As shown in Fig. 1, Ca2+ were found to be distributed at the head and the flagellar (Fig. 1a, d). However, in some cryopreserved sperm, we observed the concentration of the Ca2+ were decreased (Fig. 1b, d) because warmer colors indicate higher fluorescent intensities and increased intracellular Ca2+ (Fig. 1b, d).
DISCUSSION
In this study, we compared fertilization rate and hyperactivated motility of fresh vs. frozen and thawed mouse sperm. The sperm had been cryopreserved with a medium containing raffinose and skim milk, followed after thawing by incubation with fresh oocytes. Results showed that cleavage rate following use of frozen-thawed sperm (71.6%) was significantly lower than that from fresh sperm (94.4%) (Table 1). It seems that the frozen-thawed sperm did not have enough power to penetrate the oocyte zona pellucida which is suggested by our following result. When the oocytes with partial zona pellucida removal were in vitro fertilized with frozen-thawed sperm, cleavage rate was improved to 89.3% (Table 1). Such decreased penetration of sperm due to cryopreservation was also observed by Salamon and Maxwell (1995) and may have resulted from reduced hyperactivated motility, a normal level which is necessary for sperm cells to penetrate the cumulus matrix and zona pellucida (Suarez and Pacey, 2006).
Increases in Ca2+ concentration ([Ca2+]i) often are associated with sperm hyperactivated motility (Yanagimachi, 1994). In addition, Ca2+ acts through calmodulin on the axoneme to modulate flagellar curvature (Ho et al., 2002). Hyperactivated motility is characterized by high amplitude, asymmetrical flagellar bends and a circular or erratic swimming trajectory when intracellular Ca2+ is approximately 200B1000 nM in sperm (Ho et al., 2002; Suarez and Dai, 1995). Cytosolic Ca2+ concentration ([Ca2+]i) can be raised by either Ca2+ influx through plasma membrane ion channels (Yanagimachi, 1982) or by Ca2+ release from intracellular stores (Ho and Suarez, 2001). When Thimerosal was added to the medium of fresh sperm, flagellar bend amplitude and VCL were increased (Table 2). From this result, we infer that Thimerosal, a sulfhydryl reagent, can induce Ca2+ release from IP3R (Bindoli and Fleischer, 1983). The VCL and ALH of frozen-thawed sperm treated with Thimerosal, similar to the situation with fresh sperm, were higher than that of those without Thimerosal treatment (Table 2), suggesting that Thimerosal can stimulate release of Ca2+ from internal stores and increase hyperactivated motility of mouse frozen-thawed sperm.
Incubation of fresh or frozen-thawed sperm for 90 min under 2.4 mM Ca2+ condition led to hyperactivation (Table 3). The VCL and ALH in fresh sperm exposed to Ca2+ were significantly higher than those in fresh sperm not exposed to Ca2+ and in frozen-thawed sperm that did have Ca2+ exposure. There was no difference in ALH between the frozen-thawed sperm and the frozen-thawed sperm exposed to Ca2+. At the same time, Procaine treatment did not improve the hyperactivated motility of frozen-thawed sperm (Table 3), possibly because plasma membranes had been damaged during sperm cryopreservation (Gillan et al., 1995). Frozen-thawed sperm showed a swollen flagellar base and had coiled flagella (Nishizono et al., 2004) which likely damaged the Catspers channels, leading to impaired hyperactivated motility. Similarly, Marquez et al. (2007) reported that Procaine (5 mM) did not hyperactivate CATSPER1 or CATSPER2-/- sperm. Ca2+ entry through the principal piece-localized CATSPER channels has been implicated in the activation of hyperactivated motility (Xia et al., 2007). Mice deficient in any of the four CATSPERs are completely sterile but exhibit no other apparent abnormality (Quill et al., 2001). In this study there was no difference in Ca2+ concentration of flagella between untreated cryopreserved sperm and cryopreserved sperm treated with Ca2+ or with Procaine (data not shown).
Fluorescent imaging of fluo3-loaded sperm showed that frozen-thawed sperm had lower fluorescent intensities ([Ca2+]i) than those of fresh sperm (Fig. 1). The [Ca2+]i decrease may be associated with un-hyperactivated sperm motility which is suggested by the finding that hyperactivated motility of sperm after cryopreservation was significantly lower than that of fresh sperm (Table 2).
CONCLUSION
To sum up, in the present study, the decreased hyperactivated motility of mouse sperm due to cryopreservation may impair fertilization and Thimerosal treatment of mouse sperm after cryopreservation may increase hyperactivated motility.
ACKNOWLEDGMENT
The Authors thank BillHohenboken for revising the English version of this manuscript. The present work was supported by Funded by National Natural Science Foundation of China (No. 30972102) and high technology research and development program of China (No. 2008AA101007).
REFERENCES
- Abou-Haila, A. and D.R. Tulsiani, 2009. Signal transduction pathways that regulate sperm capacitation and the acrosome reaction. Arch. Biochem. Biophys., 485: 72-81.
PubMed - Aoki, F., D.M. Worrad and R.M. Schultz, 1997. Regulation of transcriptional activity during the first and second cell cycles in the preimplantation mouse embryo. Dev. Biol., 181: 296-307.
PubMed - Bindoli, A. and S. Fleischer, 1983. Induced Ca2+ release in skeletal muscle sarcoplasmic reticulum by sulfhydryl reagents and chlorpromazine. Arch. Biochem. Biophys., 221: 458-466.
PubMed - Dumpala, P.R., H.M. Parker and C.D. McDaniel, 2006. The effect of semen storage temperature and diluent type on the sperm quality index of broiler breeder semen. Int. J. Poult. Sci., 5: 838-845.
CrossRefDirect Link - Eisenbach, M. and L.C. Giojalas, 2006. Sperm guidance in mammals-An unpaved road to the egg. Nat. Rev. Mol. Cell Biol., 7: 276-285.
PubMed - Ho, H.C. and S.S. Suarez, 2003. Characterization of the intracellular calcium store at the base of the sperm flagellum that regulates hyperactivated motility. Biol. Reprod., 68: 1590-1596.
CrossRef - Jin, J., N. Jin, H. Zheng, S. Ro, D. Tafolla, K.M. Sanders and W. Yan, 2007. CatSper3 and CatSper4 are essential for sperm hyperactivated motility and male fertility. Biol. Reprod., 77: 37-44.
PubMed - Kawase, Y., T. Iwata, O. Ueda, N. Kamada and T. Tachibe et al., 2002. Effect of partial incision of the zona pellucida by piezo-micromanipulator for in vitro fertilization using frozen-thawed mouse spermatozoa on the developmental rate of embryos transferred at the 2-cell stage. Biol. Reprod., 66: 381-385.
PubMed - Saad, A.E.M., A.R. Alhamidi, S. Alkahtani and M.A.E.K. Sandouka, 2011. Determination of the arabian sand gazelle sperm and acrosomes defects by using the spermac staining technique. Asian J. Biol. Sci., 4: 138-147.
CrossRefDirect Link - Mortimer, S.T. and D. Mortimer, 1990. Kinematics of human spermatozoa incubated under capacitating conditions. J. Androl., 11: 195-203.
PubMed - Nishizono, H., M. Shioda, T. Takeo, T. Irie and N. Nakagata, 2004. Decrease of fertilizing ability of mouse spermatozoa after freezing and thawing is related to cellular injury. Biol. Reprod., 71: 973-978.
CrossRefPubMedDirect Link - Pena, F.J., H.R. Martinez, J.A. Tapia, C.O. Ferrusola, L.G. Fernandez and B.M. Garcia, 2009. Mitochondria in mammalian sperm physiology and pathology: A review. Reprod. Domest. Anim., 44: 345-349.
PubMed - Polge, C., A.U. Smith and A.S. Parkes, 1949. Revival of spermatozoa after vitrification and dehydration at low temperatures. Nature, 1: 666-666.
CrossRef - Purdy, P.H., 2006. A review on goat sperm cryopreservation. Small Rumin. Res., 63: 215-225.
CrossRefDirect Link - Qi, H., M.M. Moran, B. Navarro, J.A. Chong and G. Krapivinsky et al., 2007. All four CatSper ion channel proteins are required for male fertility and sperm cell hyperactivated motility. Proc. Natl. Acad. Sci., 104: 1219-1223.
CrossRef - Quill, T.A., S.A. Sugden, K.L. Rossi, L.K. Doolittle, R.E. Hammer and D.L. Garbers, 2003. Hyperactivated sperm motility driven by CatSper2 is required for fertilization. Proc. Nat. Acad. Sci., 100: 14869-14874.
CrossRef - Ren, D., B. Navarro, G. Perez, A.C. Jackson, S. Hsu and Q. Shi et al., 2001. A sperm ion channel required for sperm motility and male fertility. Nature, 413: 603-609.
CrossRefDirect Link - Rubinstein, E., A.Ziyyat, J.P. Wolf, F. Le-Naour and C. Boucheix, 2006. The molecular players of sperm-egg fusion in mammals. Semin. Cell. Dev. Biol., 17: 254-263.
PubMed - Shakeri, M., H. Roshanfekr, M. Mamoei and K. Mirzadeh, 2008. Effect of epididymis handling conditions on the quality of ram spermatozoa recovered post-mortem. Asian J. Anim. Vet. Adv., 3: 400-408.
CrossRefDirect Link - Suarez, S.S. and X. Dai, 1995. Intracellular calcium reaches different levels of elevation in hyperactivated and acrosome-reacted hamster sperm. Mol. Reprod. Dev., 42: 325-333.
CrossRef - Suarez, S.S. and H.C. Ho, 2003. Hyperactivation of mammalian sperm. Cell. Mol. Biol., 49: 351-356.
PubMed - Suarez, S.S. and A.A. Pacey, 2006. Sperm transport in the female reproductive tract. Hum. Reprod. Update, 12: 23-37.
Direct Link - Salamon, S. and W.M.C. Maxwell, 1995. Frozen storage of ram semen II. Causes of low fertility after cervical insemination and methods of improvement. Anim. Reprod. Sci., 38: 1-36.
CrossRef - Salamon, S. and W.M.C. Maxwell, 2000. Storage of ram semen. Anim. Reprod. Sci., 62: 77-111.
PubMedDirect Link - Sundararaman, M.N., J. Kalatharan and K.T.P. Jawahar, 2007. Analyses of morphological and morphometrical deviations of bull spermatozoa by computer assisted semen analysis technique. Asian J. Anim. Vet. Adv., 2: 196-204.
CrossRefDirect Link - Sundararaman, M.N. and M.J. Edwin, 2008. Changes in motility characteristics of goat spermatozoa during glycerol-equilibration and the relevance to cryopreservation. Asian J. Cell Biol., 3: 22-33.
CrossRefDirect Link - Xia, J., D. Reigada, C.H. Mitchell and D. Ren, 2007. Catsper channel-mediated Ca2+entry into mouse sperm triggers a tail-to-head propagation. Biol. Reprod., 77: 551-559.
PubMed - Yanagimachi, R., 1982. Requirement of extracellular calcium ions for various stages of fertilization and fertilization-related phenomena in the hamster. Gamete Res., 5: 323-344.
CrossRef - Wennemuth, G., R.E. Westenbroek, T. Xu, B. Hille and D.F. Babcock, 2000. CaV2.2 and CaV2.3 (N- and R-type) Ca2+ channels in depolarization-evoked entry of Ca2+ into mouse sperm. J. Biol. Chem., 275: 21210-21217.
PubMed