
M. Habtamu Taddele
Mekelle University College of Veterinary Medicine, P.O. Box 1436, Mekelle, Tigray, Ethiopia
R. Rathore
Centre for Animal Disease Research and Diagnosis (CADRAD), Indian Veterinary Research Institute, Izatnagar, Bareilly (U.P.)-243 122, India
K. Dhama
Avian Diseases Section, Division of Veterinary Pathology, Indian Veterinary Research Institute, Izatnagar (U.P.)-243 122, India
R.K. Agarwal
Division of Veterinary Bacteriology and Mycology, Indian Veterinary Research Institute, Izatnagar (U.P.)-243 122, India
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 11 | Page No.: 1037-1051







ABSTRACT
The present study was conducted to evaluate the discriminating power of RAPD-PCR and PCR-RFLP to characterize Salmonella gallinarum (S. gallinarum) isolates. Fifteen S. gallinarum isolates of poultry origin recovered from various sources in different regions of India including three reference strains were characterized using RAPD-PCR and PCR-RFLP. RAPD-PCR, using 5 primers (NSC I, NSC II, NSC III, URP-6 and primer 1290), grouped S. gallinarum isolates into 4-6 RAPD types. The discrimination power (D) of NSC II, URP-6 and primer 1290 was 0.848, 0.81 and 0.695, respectively and 0.62 for NSC I and NSC III. The phylogenetic dendrogram classified the isolates based on their geographic location. However, most S. gallinarum isolates were clustered closely together. In comparative analysis NSC II primer was placed in a separate cluster. The PCR amplification of the 16s rRNA and fimH genes produced approximately 572 bp and 1008 bp product, respectively, for all S. gallinarum isolates. Restriction Endonuclease (RE) enzymes viz., EcoRI, SmaI, AluI, BgiII, MspI and HaeIII and SalI, AluI, MspI and HaeIII were used to digest PCR products of 16s rRNA and fimH genes, respectively. These restriction enzymes generated 2-4 bands of varying molecular size ranging from 40 to 410 bp and 50 to 620 bp for 16s rRNA and fimH genes, respectively. AluI targeted against the fimH gene was the only enzyme which classified S. gallinarum isolates into three groups. In conclusion, the RAPD-PCR profile proved to be used as an epidemiological typing tool since it classified the S. gallinarum isolates according to their geographic origin.
PDF Abstract XML References Citation
Received: March 21, 2011;
Accepted: July 26, 2011;
Published: September 28, 2011
How to cite this article
M. Habtamu Taddele, R. Rathore, K. Dhama and R.K. Agarwal, 2011. Epidemiological Characterization of Salmonella gallinarum Isolates of Poultry Origin in India, Employing Two PCR Based Typing Methods of RAPD-PCR and PCR-RFLP. Asian Journal of Animal and Veterinary Advances, 6: 1037-1051.
DOI: 10.3923/ajava.2011.1037.1051
URL: https://scialert.net/abstract/?doi=ajava.2011.1037.1051
DOI: 10.3923/ajava.2011.1037.1051
URL: https://scialert.net/abstract/?doi=ajava.2011.1037.1051
INTRODUCTION
Indian poultry industry is emerging as the world’s 2nd largest market with a compounded annual growth rate of 12 and 15% (http://ipema.co.in/Company20/Default.aspx,Indian poultry equipment manufacturer’sassociation). Although, the poultry sector contributes its great share to the country’s economy, the production is hampered by several disease conditions. Among diseases of poultry Salmonellosis is of great concern and has been responsible for serious economic losses to the poultry producers (Gast and Shivprasad, 2003; Kabir, 2010). In recent years problems related to Salmonella have increased significantly due to emergence of multi-drug resistant Salmonella (Kabir, 2010). More than 2500 serologically distinct Salmonella variants are widely distributed worldwide (Popoff et al., 2004).
Fowl typhoid caused by Salmonella enterica subspecies enterica serovar Gallinarum biovar Gallinarum prevails to be one of the major Salmonella serotype isolated from poultry in India (Agrawal et al., 2005; Prakash et al., 2005). Fowl typhoid is responsible for immense economic losses due to high morbidity and mortality in all age groups of birds. Although, fowl typhoid has been eradicated from most of the developed countries through improved surveillance and slaughter practices (Baumler et al., 2000; Jin et al., 2006), the disease is still of major economic importance to the developing world (Kabir, 2010).
Various molecular typing techniques have been used in epidemiological studies to differentiate isolates of Salmonella serovars. However, some of these molecular techniques are of little value for epidemiological typing due to limitations such as cost effectiveness, ease of use and availability of facilities. The introduction of PCR, the most widely used molecular tool for detection of several pathogens, has strongly increased the number of various PCR-based typing methods for characterization of important pathogens.
Random Amplified Polymorphic DNA (RAPD) or Arbitrarily Primed PCR (AP-PCR) has been widely used for studying genetic diversity and phylogenetic relationships of bacteria (Jin et al., 2006; Lin et al., 1996). The method generates distinctive DNA fingerprints by using a single, short (10-25 mer) synthetic primer in PCR. RAPD is faster, simpler and more economical than other genetic typing assays (Gomes et al., 2005; Hassanien, 2008). A further advantage of RAPD is that knowledge of the DNA sequence of the target organism is not required (Maripandi et al., 2007; Mollah et al., 2005). However, reproducibility can be a problem under certain RAPD conditions due to low annealing temperature required for PCRs that use arbitrary primers of very short length. For this reason both short and long oligonucleotide primers were used in this study.
PCR-Restriction Fragment Length Polymorphism (RFLP) is one of the amplification-based techniques that have been developed to determine genetic variability which is based on amplification of specific targets and subsequent digestion with restriction enzymes (Yan et al., 2003). Kwon et al. (2000) reported that PCR-RFLP was applied to differentiate the fliC (iroB) amplicons with the restriction enzyme, Hinp1I that recognizes the region of biotype Gallinarum but not in biotype Pullorum. Using PCR-RFLP they could accurately and rapidly differentiate biotype Pullorum and biotype Gallinarum. Similarly, Kisiela et al. (2005) digested PCR amplicons of fimH gene with SacI enzyme to successfully differentiate between these two biovars. More recently, Cheah et al. (2008) reported for excellent type ability of PCR-RFLP but with low discrimination power in characterizing S. weltevreden isolates. Therefore, the aim of the present study was to evaluate the discriminating power of two of PCR based typing methods, RAPD-PCR and PCR-RFLP analysis on S. gallinarum isolates from different geographic regions of India.
MATERIALS AND METHODS
Bacterial strains: A total of 12 S. gallinarum isolated from various sources in different regions of India and 3 reference S. gallinarum strains were procured from the National Salmonella Centre, Indian Veterinary Research Institute, Izatnagar (Table 1).
| Table 1: | Host and epidemiological sources of Salmonella gallinarum isolates |
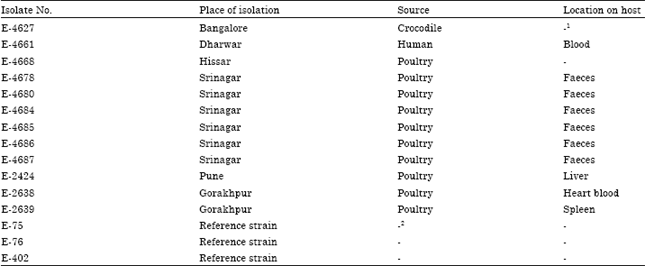 | |
| 1Location on the host not known, 2Source not known | |
| Table 2: | Sequences of different primers used in the study |
 | |
| URP: Universal Rice Primer, RAPD: Randomly amplified polymorphic DNA, 165F Frd: 16s ribosomal RNA forward primer, PCR: Polymerase chain reaction, 16S III Rov: 16s ribosomal RNA reverse primer, FinH Frd: Fimbrial gene forward primer, FinH Rev: Fimbrial gene reverse primer | |
Primers: Five RAPD-PCR primers and primers for PCR amplification of 16s rRNA and fimH genes of Salmonella used in the present study are shown in Table 2. Oligonucleotide primers for PCR amplification of a 572 bp product of Salmonella 16s rRNA gene and 1008 bp product of Salmonella fimH gene were used in the present study.
Maintenance of strains: S. gallinarum strains obtained from the National Salmonella Centre, IVRI, Izatnagar were checked for their biochemical profiles and inoculated on nutrient agar slants and kept at 4°C. Sub-culturing was done periodically to test the purity, morphological and biochemical characteristics of the strains.
Isolation of genomic DNA: Total DNA isolation was carried out using Forensic DNA isolation kit according to the manufacturers’ instruction (Omega, USA.). Isolated DNA was stored at -20°C till use.
Randomly Amplified Polymorphic DNA (RAPD) PCR assay: Randomly amplified polymorphic DNA PCR (RAPD-PCR) was performed using genomic DNA of the test S. gallinarum strains as template with the random primers mentioned in Table 2. Each polymerase chain reaction (PCR) mixture consisted of 20 ng of template DNA, 2.5 μL 10xPCR buffer with (NH4)2SO4, 1 μL of 10 mM dNTPs, 2.5 μL of 25 mM MgCl2, 1 unit of Taq DNA polymerase (MBI Fermentas, USA) and 15 picomolar of each primer (SBS Genetech, India) in 25 μL PCR reaction mix. The PCR programme for URP-6 included initial denaturation at 94°C for 5 min followed by 42 cycles of denaturation (94°C for 1 min), annealing (50°C for 45 sec) and extension (72°C for 1.5 min). Final extension was carried out at 72°C for 5 min. For primers 1290, NSC I, NSC II and NSC III the PCR reaction mix and cycling conditions were the same with the exception of their annealing temperature which was adjusted at 25, 25, 27 and 21°C, respectively. The PCR products were characterized by submarine gel electrophoresis on 1.2% agarose gel. Gene RulerTM 100 bp (MBI Fermentas, USA) and low range DNA ruler (Bangalore Genei) were used as molecular weight marker in first and last lanes.
Analysis of RAPD data: Each isolate was scored for the presence or absence (1 or 0) of each band on agarose gel. The index of similarity (F) between samples was calculated using the formula of Nei and Li (1979) Fxy = 2nxy/nx+ny. Where nxy is the number of RAPD bands shared by the two samples and nx and ny are the numbers of RAPD bands scored in each sample. The distance matrices, mean and standard deviation were calculated using the STATISTICA software and this result was directly used to construct the phylogenetic relationships among the isolates by the neighbour-joining method using STATISTICA software. The numerical index of discrimination (D) was calculated using the Simpson’s index of diversity (Hunter and Gaston, 1988) using the following formula:
where, N is the total number of strains in the sample population, s is the total number of types described and nj is the number of strains belonging to the jth type.
PCR-restriction Fragment Length Polymorphism (PCR-RFLP) analysis
PCR amplification: PCR reaction mix was prepared in 25 μL which contains 50 ng of template DNA, 1x PCR assay buffer with (NH4)2SO4 and 2.5 mM MgCl2. The dNTPs at a concentration of 200 μM, 1 U of Taq DNA polymerase (MBI Fermentas, USA) and 15 picomol solution of each primer (SBS Genetech, India) were also added to the mixture. In this study, PCR assay was performed in Gene Amp PCR System 9700 (Applied Biosystems, India). The cycling conditions for amplification of 16s rRNA include 94°C for 5 min (predenaturation), 35 cycles of 94°C for 1 min (denaturation), 55°C for 45 sec (annealing) and 72°C for 1 min (polymerization) followed by 72°C for 10 min (final extension). The cycling conditions for the amplification of fimH gene were as above except annealing temperature (57°C for 45 sec). The above PCR cycling conditions were chosen on the basis of preliminary experiments performed to optimise amplification of Salmonella target sequences. All amplifications included a blank which contained all reagents but not the target DNA. DNA markers of 100 bp (MBI Fermentas, USA) were used as a molecular weight marker. The amplification products were checked for the presence of the desired bands on 1.2% agarose gel.
| Table 3: | RE enzymes and their respective sites for 16s rRNA and fimH genes |
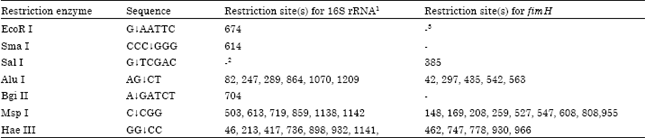 | |
| 10 Checked with full 16s rRNA gene sequence (1234 bp) available at GenBank (NCBI), 21 Has no restriction site on fimH gene, 32 Has no restriction site on 16s rRNA | |
RE digestion of PCR product and RFLP profile by gel electrophoresis: Both genes, 16s rRNA and fimH, were checked for their restriction sites using Gene Tool software and accordingly the following restriction enzymes were selected. The restriction enzymes EcoRI, SmaI, AluI, BgiII, MspI and HaeIII (Bangalore Genei, India and Fermentas, USA) were used for digesting the 16s rRNA gene PCR product and SalI, AluI, MspI and HaeIII for fimH gene (Table 3).
RESULTS
RAPD PCR profile: The profiles generated by RAPD PCR with the five primers were composed of two to nine major bands. Each fingerprint contained varying size bands ranging from 0.2 kb to approximately 3 kb. The RAPD PCR was performed in triplicate to ensure for reproducibility and banding patterns for identical samples were indistinguishable.
URP-6 has revealed approximately a consistent 0.8 and 1.25 kb bands in all the S. gallinarum isolates including reference strains. Three major bands of molecular weight around 2.5, 1.8 and 0.2 kb were produced in 12, 11 and 10 isolates including reference strains, respectively (Fig. 1a).
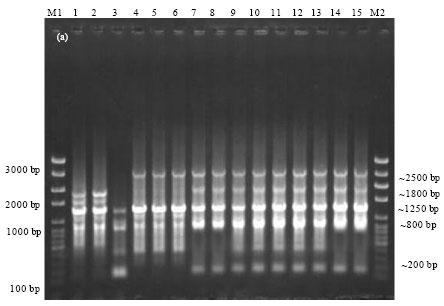 | |
| Fig. 1a: | RAPD-PCR profiles of S. gallinarum isolates with URP-6 primer. Lane M1 and M2: Molecular weight marker; Lane 1: E4627; Lane 2: E4661; Lane 3: E4668; Lane 4: E4678; Lane 5: E4680; Lane 6: E4684; Lane 7: E4685; Lane 8: E4686; Lane 9: E4687; Lane 10: E75; Lane 11: E76; Lane 12: E2424; Lane 13: E2638; Lane 14: E2639; Lane 15: E402 |
 | |
| Fig. 1b: | RAPD-PCR profiles of S. gallinarum isolates with primer 1290. Lane M1 and M2: Molecular weight marker; Lane 1: E4627; Lane 2: E4661; Lane 3: E4668; Lane 4: E4678; Lane 5: E4680; Lane 6: E4684; Lane 7: E4685; Lane 8: E4686; Lane 9: E4687; Lane 10: E75; Lane 11: E76; Lane 12: E2424; Lane 13: E2638; Lane 14: E2639; Lane 15: E402 |
Approximately 1.0 and 1.2 kb bands were produced uniformly in all isolates and reference strains with the primer 1290 (Fig. 1b). Additional bands of approximate molecular weight 1.8 and 1.5 kb were noticed in 9 isolates including two reference strains.
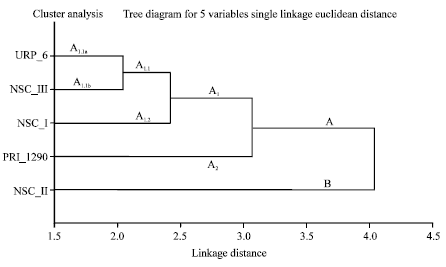
With Primer NSC I, except one strain which lacked 1.75 and 0.4 kb bands, all other isolates and reference strains showed bands of approximate molecular size 1.75, 1.2, 0.7 and 0.4 kb, consistently. With the same primer weak band of approximately 1.4 kb size was detected in all the isolates except one (E-4668). Three isolates, E-4678, E4680 and E-4684, were containing additional 1.0 and 0.9 kb bands (Fig. 1c). NSC II primer produced different RAPD profile for most of the strains compared to other four primers used in the study showing great variation (Fig. 1d) and having better discrimination power, 0.848. However, approximately 1.7 kb band was consistent in occurrence in all the isolates. The RAPD profile produced by NSC III primer contained approximately 1.25 and 0.8 kb fragments uniformly in all the isolates (Fig. 1e). Except two isolates, E-4627 and E-4661, the remaining 13 isolates possessed a band of molecular size around 2.5 kb.
Analysis of RAPD profile and dendrogram construction: Primers 1290 and URP-6 classified S. gallinarum isolates in to 5 RAPD types (Fig. 2a, b); whereas NSC I and NSC III, classified the isolates in to 4 RAPD types (Fig. 2c, d) and NSC II grouped the isolates into 6 RAPD types (Fig. 2e). The highest Discrimination power (D) was found with NSC II (0.848), followed by URP-6 (0.810) and primer 1290 (0.695). The discrimination power of NSC I and NSC III was 0.62.
The comparative statistical analysis classified the five RAPD primers used in this study into two major groups where the maximum dissimilarity distance was around 4.0 units (Fig. 2f). Four primers (URP-6, Primer 1290, NSC I and NSC III) were grouped in one cluster and NSC II in a separate cluster. According to the comparative statistical analysis URP-6 and NSC III are closely related in classifying the S. gallinarum isolates.
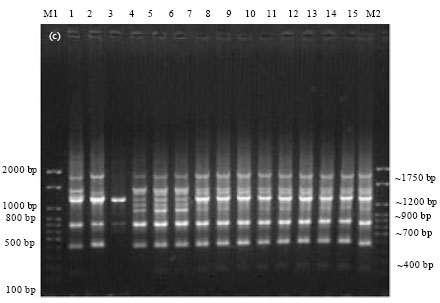 | |
| Fig. 1c: | RAPD-PCR profiles of S. gallinarum isolates with NSC I primer. Lane M1 and M2: Molecular weight marker; Lane 1: E4627; Lane 2: E4661; Lane 3: E4668; Lane 4: E4678; Lane 5: E4680; Lane 6: E4684; Lane 7: E4685; Lane 8: E4686; Lane 9: E4687; Lane 10: E75; Lane 11: E76; Lane 12: E2424; Lane 13: E2638; Lane 14: E2639; Lane 15: E402 |
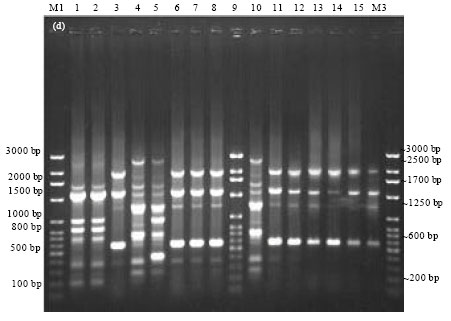 | |
| Fig. 1d: | RAPD-PCR profiles of S. gallinarum isolates with NSC II primer. Lane M1, M2 and M3: Molecular weight marker; Lane 1: E4627; Lane 2: E4661; Lane 3: E4668; Lane 4: E4678; Lane 5: E4680; Lane 6: E4684; Lane 7: E4685; Lane 8: E4686; Lane 9: E4687; Lane 10: E75; Lane 11: E76; Lane 12: E2424; Lane 13: E2638; Lane 14: E2639; Lane 15: E402 |
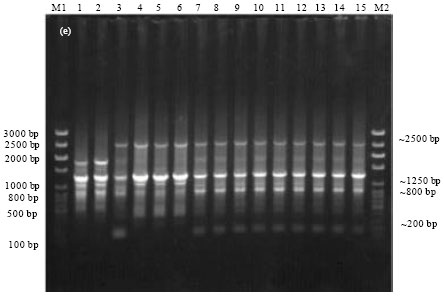
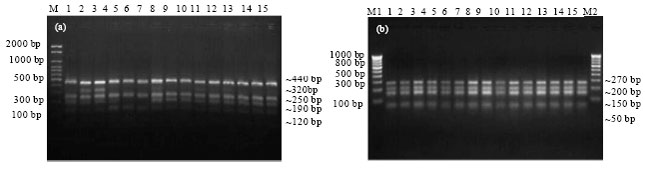
PCR-RFLP analysis of S. gallinarum isolates: PCR amplification of the 16s rRNA gene loci (rrn) and fimH gene of S. gallinarum isolates generated a product of 572 and 1008 bp, respectively. The band size detected in all the S. gallinarum isolates was consistent for both genes as analysed by agarose gel electrophoresis (Fig. 3).
 | |
| Fig. 1e: | RAPD-PCR profiles of S. gallinarum isolates with NSC III primer. Lane M1 and M2: Molecular weight marker; Lane 1: E4627; Lane 2: E4661; Lane 3: E4668; Lane 4: E4678; Lane 5: E4680; Lane 6: E4684; Lane 7: E4685; Lane 8: E4686; Lane 9: E4687; Lane 10: E75; Lane 11: E76; Lane 12: E2424; Lane 13: E2638; Lane 14: E2639; Lane 15: E402 |
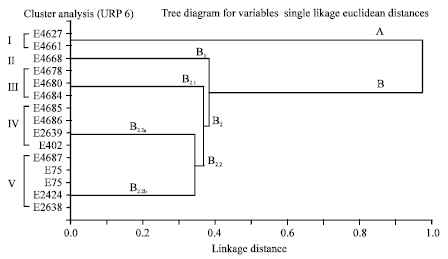 | |
| Fig. 2a: | Dendrogram computed based on RAPD analysis of S. gallinarum isolates using URP 6 |
Restriction Endonuclease (RE) profile of PCR products: The PCR products subjected to selected RE produced two to four bands of varying molecular size. Bands ranging approximately from 40 to 410 bp and 50 to 620 bp for 16s rRNA and fimH genes were produced, respectively. Four of the restriction enzymes (EcoRI, SmaI, BgiII and AluI) produced two fragments in 16s rRNA gene. Whereas, the other two enzymes (HaeIII and MspI) produced three and four major bands, respectively with similar banding pattern for all the S. gallinarum strains.
The enzymes used for digesting the PCR product of fimH gene revealed two bands with SalI enzyme and four bands with AluI, MspI and HaeIII enzymes. Out of the four restriction enzymes used for fimH gene the enzyme AluI showed different patterns of fragments which classified the S. gallinarum isolates into three major groups (Fig. 4). With all the isolates, bands of approximate molecular size 440 and 250 bp were consistent.
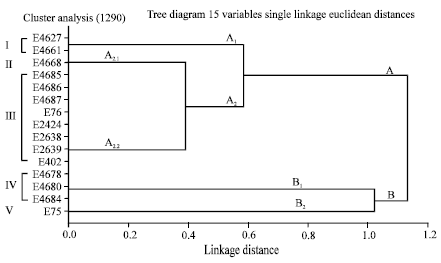 | |
| Fig. 2b: | Dendrogram computed based on RAPD analysis of S. gallinarum isolates using primer 1290 |
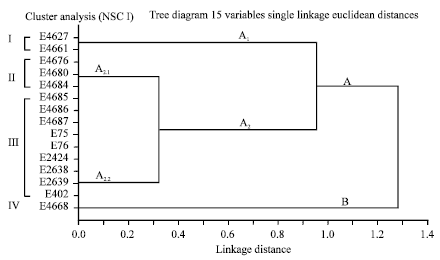 | |
| Fig. 2c: | Dendrogram computed based on RAPD analysis of S. gallinarum isolates using NSC I |
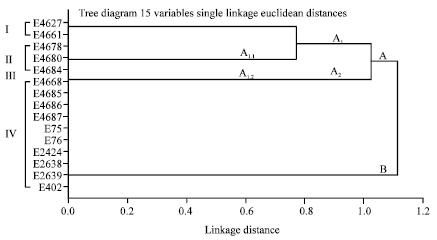 | |
| Fig. 2d: | Dendrogram computed based on RAPD analysis of S. gallinarum isolates with NSC III |
 | |
| Fig. 2e: | Dendrogram computed based on RAPD analysis of S. gallinarum isolates using NSC II |
 | |
| Fig. 2f: | Dendrogram computed based on comparative analysis of the 5 primers |
 | |
| Fig. 3 (a-b): | PCR amplification of 16s rRNA gene (A) and fimH gene (B) of 15 S. gallinarum isolates. Lane M: Molecular weight marker; Lane 1: E4627; Lane 2: E4661; Lane 3: E4668; Lane 4: E4678; Lane 5: E4680; Lane 6: E4684; Lane 7: E4685; Lane 8: E4686; Lane 9: E4687; Lane 10: E75; Lane 11: E76; Lane 12: E2424; Lane 13: E2638; Lane 14: E2639; Lane 15: E402 |
 | |
| Fig. 4 (a-b): | RE digestion of fimH gene of S. gallinarum isolates with AluI (A) and MspI (B). Lane M: Molecular weight marker; Lane 1: E4627; Lane 2: E4661; Lane 3: E4668; Lane 4: E4678; Lane 5: E4680; Lane 6: E4684; Lane 7: E4685; Lane 8: E4686; Lane 9: E4687; Lane 10: E75; Lane 11: E76; Lane 12: E2424; Lane 13: E2638; Lane 14: E2639; Lane 15: E402 |
The discriminating power of this enzyme was around 0.533. RE enzymes SalI, MspI and HaeIII produced same banding pattern with all the isolates indicating highly conserved nature of the fimH gene.
DISCUSSION
Molecular typing methods have been found essential and useful in epidemiological investigations of closely related bacterial strains (Gillespie and Oliver, 2004). Subtyping of Salmonella serovars are considered to be of practical and clinical importance in tracing the epidemiological status and investigations of outbreaks. In contrast to phenotypic characterization, nucleic acid based typing could provide a more precise strain differentiation of Salmonella (Esteban et al., 1993; Gillespie and Oliver, 2004). However, some of these molecular techniques are of little value for epidemiological typing due to limitations such as cost effectiveness, ease of use and availability of facilities. The introduction of PCR has increased the number of various PCR-based typing methods which are helpful in differentiation and characterization of pathogens of veterinary importance. Hence, in the present study the two PCR based typing techniques, RAPD-PCR and PCR-RFLP were used for characterizing the S. gallinarum isolates of different geographic origin.
The RAPD/PCR used in this study can be an epidemiological tool for the typing of indistinct and related isolates, excluding unrelated ones (Maripandi et al., 2007). The RAPD method using both short and long arbitrary oligonucleotide primers was chosen in this study to evaluate the genomic diversity among S. gallinarum isolates. RAPD-PCR assays are simpler, faster, more convenient and easier to perform than most other molecular typing methods. However, only when used under well defined and optimized conditions RAPD is capable of reproducing the amplification of random fragments of DNA and generating high degrees of polymorphism (Lin et al., 1996; Maripandi et al., 2007; Soto et al., 1999). RAPDs have also been reported to detect higher levels of polymorphism, Hassanien (2008) compared, for example with RFLPs in cases where the two have been applied in the same material. The success of this technique is due to the fact that knowledge of the target genome is not required. Furthermore, only single random and short 10-mer oligonucleotide primers are used in any genome (Maripandi et al., 2007).
Among all the 5 primers used, NSC II produced immense genetic variability to the maximum extent and this may be due to the difference in discriminating ability of the primer. The discriminating power (D), 0.848, of this primer was found to be much better than other primers used in the study. A report by Meenu (2002) showed maximum genetic variability among S. paratyphi B isolates using NSC II primer when compared to NSC I and NSC III. If RAPD is to be useful as a typing method, a sufficient discriminatory number of amplified fragments must be produced which essentially depends on the primer-template interaction (Saxena, 2000). RAPD primers bind at random sites of the genomic DNA and if it binds in variable regions, products with great variability can be produced (Maripandi et al., 2007). However, if the primer binds in a highly conserved region similar types of profiles may be produced with repeatability. The RAPD primers NSC I and NSC III proved to be used as efficient RAPD primers regardless of their short oligonucleotide sequence. Even though the discrimination power of these primers was less compared to others but they were able to discriminate the human origin S. gallinarum into separate RAPD type and also other isolates according to their geographic location.
The result of URP-6 which grouped the isolates into 5 RAPD types is in close correlation with the works of Jin et al. (2006). However, similar result obtained with primer 1290 did not agree with their result in which they observed only 2 RAPD types. This variation may be because of the difference in annealing temperature of the primers and also reaction mixture. Additionally the difference in geographic location of the isolates may be taken into consideration.
With all the primers S. gallinarum isolates from Bangalore and Dharwar of the Karnataka state belong to the same RAPD type and this indicates the close relationship of these isolates at genetic level. The two isolates from Gorakhpur were also placed in same RAPD type in all the primers except URP-6 signifying the close relation of the isolates at their genetic level. The strain, E-4668, from Hissar was placed in a separate RAPD type in four of the primers and in primer NSC II, it was grouped together with the three reference strains and with isolate from Pune. This strain was isolated from human blood and this may be the main reason to be grouped separately.
Interestingly, the isolate from Pune was grouped together with the reference strains with four of the primers (NSC I, NSC II, NSC III and primer 1290) and with URP-6 it was kept in one group (RAPD type V) with the two reference strains, E-75 and E-76 and near to reference strain E-402. This clearly showed very close relatedness of this isolate to the reference strains at the genetic level. With primers NSC I, NSC III, URP-6 and primer 1290, three isolates (E-4678, E-4680 and E-4684) from Srinagar were positioned in one RAPD type indicating for close relation of these isolates. The dendrogram computed for most of the primers classified the S. gallinarum isolates based on their geographic location. These results proved that RAPD can be used as a potent epidemiological tool for typing Salmonella serotypes.
The phylogenetic analysis of S. gallinarum isolates showed that even though they can be grouped in different RAPD types according to their geographic origin, they are closely related to each other since most S. gallinarum isolates were clustered closely together in the phylogenetic dendrogram. The descriptive statistics based on mean and standard deviation computed against each primer indicated for the closely relatedness of the isolates. The largest difference observed among the 15 S. gallinarum isolates tested involved 5 bands and this difference is produced with NSC II primer. The maximum dissimilarity distance observed was in the range of 1.0 to 1.3 units which magnified for the close relation of these gallinarum isolates.
The comparative phylogenetic dendrogram computed against the primers used clearly indicated that primer NSC II is different from the others. URP-6 and NSC III were more closely related in their outputs with maximum dissimilarity distance of 2.0 units. URP-6 was the next primer giving high discrimination (0.810) between isolates which classified the isolates into 5 groups. Primer 1290 though classified the isolates into 5 groups like URP-6 its power of discrimination was less, 0.695. The present study clearly indicated that both long and short oligonucleotide primers can be used in combination or separately but the use of more than one primer is advantageous as it enhanced the discriminatory power of the typing method. Similar primers have also been used by other workers on different Salmonella serotypes and showed good discrimination power (Jin et al., 2006; Meenu, 2002; Quintaes et al., 2004).
All the 5 primers (NSC I, NSC II, NSC III, URP-6 and primer 1290) produced reproducible result. Even though reproducibility was maintained but there were variations between some of the primers used. These observations revealed that RAPD profile can be best used as a complimentary epidemiological tool in combination with other molecular and phenotypic typing techniques. Similar interpretations have been reported by several investigators on the same serotype as well as other serotypes (Jin et al., 2006; Maripandi et al., 2007; Meenu, 2002; Morshed and Peighambari, 2010; Dos Santos et al., 2008; Yaqoob et al., 2007).
PCR amplification of the 16s rRNA (rrn) gene and fimH gene of S. gallinarum isolates gave a uniform product size of approximately 572 bp and 1008 bp, respectively. This indicated the highly conserved nature of these two genes within the genus. This was further proved by the restriction enzyme profiles. The restriction enzymes utilized for this study were selected on the basis of their restriction sites for the target genes and RE enzymes having 1-6 restriction sites were used. Among all the enzymes used against fimH gene only AluI was able to produce different banding patterns among the S. gallinarum isolates. The results of other restriction enzymes used in the study provided uniform banding patterns for all the S. gallinarum isolates. This indicated highly conserved nature of the genes, 16s rRNA and fimH. The result of the restriction enzyme AluI showed its potential to be used in epidemiological investigations of Salmonella.
In restriction digestion the number of fragments should be sufficient to enable good discrimination but not so many that they overlap or are coincidental on the gel, thereby resulting in a crowded fingerprint which is difficult to interpret (Towner and Cockayne, 1993). The result obtained by digestion of the fimH gene with AluI supported this fact. Results obtained exhibited that PCR-RFLP has good repeatability but low discriminatory power due to its inability to produce different banding pattern within S. gallinarum isolates. The discriminatory power of PCR-RFLP is generally poor compared to other amplification-based techniques, like RAPD. This is primarily due to the limited region of the genome that is examined and this finding is in accordance with the interpretation of Cheah et al. (2008).
CONCLUSION
In this study the RAPD-PCR profile proved to be used as an epidemiological typing tool since it classified the S. gallinarum isolates according to their geographic origin. RAPD-PCR fingerprint strategy should be applicable to bacterial typing for its rapidness, simplicity, low cost and potential to generate polymorphisms. Primer NSC II showed better discriminatory power compared to other primers. PCR-RFLP using the restriction enzyme AluI classified the S. gallinarum isolates into three types indicating for the efficiency of this enzyme in epidemiological investigations. The discriminating power of PCR-RFLP can be enhanced further by targeting genes showing great variability between serotypes and within serotype.
ACKNOWLEDGMENTS
Material and financial support provided by the Division of Bacteriology and Mycology, Indian Veterinary Research Institute, Izatnagar, Bareilly (U.P.), India is acknowledged by the authors. The National Salmonella Centre, Indian Veterinary Research Institute, Izatnagar, Bareilly (U.P.), India is also highly acknowledged for providing the Salmonella gallinarum isolates including the reference strains. The authors are also thankful to Prof. SK khar for his contribution in editing this manuscript.
REFERENCES
- Agrawal, R.K., B.R. Singh, N. Babu and M. Chandra, 2005. Novel haemolysins of Salmonella enterica spp. enterica serovar Gallinarum. Indian J. Exp. Biol., 43: 626-630.
PubMed - Cheah, Y.K., N.A. Salleh, L.H. Lee, S. Radu, S. Sukardi and J.H. Sim, 2008. Comparison of PCR fingerprinting techniques for the discrimination of Salmonella enterica subsp. enterica serovar Weltevreden isolated from indigenous vegetables in Malaysia. World J. Microbiol. Biotechnol., 24: 327-335.
CrossRef - Esteban, E., K. Snipes, D. Hird, R. Kasten and H. Kinde, 1993. Use of ribotyping for characterization of Salmonella serotypes. J. Clin. Microbiol., 31: 233-237.
Direct Link - Gillespie, B.E. and S.P. Oliver, 2004. Comparison of an automated ribotyping system, pulsed-field gel electrophoresis and randomly amplified polymorphic DNA fingerprinting for differentiation of Streptococcus uberis strains. Biotechnology, 3: 165-172.
CrossRefDirect Link - Gomes, A.R., L. Muniyappa, G. Krishnappa, V.V.S. Suryanarayana, S. Isloor, B. Prakash and P.G. Hugar, 2005. Genotypic characterization of avian Escherichia coli by random amplification of polymorphic DNA. Int. J. Poult. Sci., 4: 378-381.
CrossRefDirect Link - Hassanien, H.A., 2008. Use of Randomly Amplified Polymorphic DNA (RAPD) analysis to detect genetic variation in sea bass (Dicentrarchus labrax). J. Fish. Aquat. Sci., 3: 39-46.
CrossRefDirect Link - Hunter, P.R. and M.A. Gaston, 1988. Numerical index of the discriminatory ability of typing system: An application of Simpsons index of diversity. J. Clin. Microbiol., 26: 2465-2466.
PubMedDirect Link - Kwon, H.J., K.Y. Park, H.S. Yoo, J.Y. Park, Y.H. Park and S.J. Kim, 2000. Differentiation of Salmonella enterica serotype Gallinarum biotype Pullorum from biotype Gallinarum by analysis of phase 1 flagellin C gene (fliC). J. Microbiol. Methods, 40: 33-38.
CrossRef - Jin, J.D., D.S. Lee, E.K. Shin, S.J. Kim, R. Jung and T.W. Hanh, 2006. Molecular typing by random amplification of polymorphic DNA (RAPD) and detection of virulence genes of S. Enterica subsp enterica serovar Gallinarum biovar Gallinarum. J. Vet. Med. Sci., 68: 1321-1326.
PubMed - Kabir, S.M.L., 2010. Avian colibacillosis and salmonellosis: A closer look at epidemiology, pathogenesis, diagnosis, control and public health concerns. Int. J. Environ. Res. Public Health, 7: 89-114.
CrossRefPubMedDirect Link - Kisiela, D., M. Kuczkowski, L. Kiczak, A. Wieliczko and M. Ugorski, 2005. Differentiation of Salmonella gallinarum biovar Gallinarum from Salmonella gallinarum biovar Pullorum by PCR-RFLP of the fimH gene. J. Vet. Med. Ser. B, 52: 214-218.
CrossRef - Lin, A.W., M.A. Usera, T.J. Barrett and R.A. Goldsby, 1996. Application of random Amplified polymorphic DNA analysis to differentiate strains of Salmonella enteritidis. J. Clin. Microbiol., 34: 870-876.
Direct Link - Maripandi, A., S.S. Suresh Raja, P. Ponmurugan and G. Gurusubramanian, 2007. Random Amplification of Polymorphic DNA (RAPD) of Salmonella enteritidis isolated from chicken samples. Biotechnology, 6: 278-282.
CrossRefDirect Link - Mollah, M.B.R., M.S. Alam, F.B. Islam and M.A. Ali, 2005. Effectiveness of RAPD marker in generating polymorphism in different chicken population. Biotechnology, 4: 73-75.
CrossRefDirect Link - Morshed, R. and S.M. Peighambari, 2010. Drug resistance, plasmid profile and random amplified polymorphic DNA analysis of Iranian isolates of Salmonella enteritidis. New Microbiol., 33: 47-56.
PubMed - Nei, M. and W.H. Li, 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA., 76: 5269-5273.
CrossRefPubMedDirect Link - Popoff, M.Y., J. Bockemuhl and L.L. Gheesling, 2004. Supplement 2002 (No. 46) to the Kauffmann-White scheme. Res. Microbiol., 155: 568-570.
PubMed - Prakash, B., G. Krishnappa, L. Muniyappa and B.S. Kumar, 2005. Epidemiological characterization of avian Salmonella enterica serovar infections in India. Int. J. Poult. Sci., 4: 388-395.
CrossRefDirect Link - Dos Santos, L.R., R.A. Ribeiro, S.D. de Oliveira, L.B. Rodrigues, M.L. Flores, R.F.F. Lopes and V.P. do Nascimento, 2008. RAPD/PCR and phage typing of Salmonella enteritidis isolated from poultry and food poisoning outbreaks. Sao Paulo Arq. Inst. Biol., 75: 91-94.
Direct Link - Soto, S.M., B. Guerra, M.A. Gozalez-Hevia and M.C. Mendoza, 1999. Potential of three-way randomly amplified polymorphic DNA analysis as a typing method for twelve Salmonella serotypes. Applied Environ. Microbiol., 65: 4830-4836.
Direct Link - Yan, S.S., M.L. Pendrak, B. Abela-Ridder, J.W. Punderson, D.P. Fedorko and S.L. Foley, 2003. An overview of Salmonella typing public health perspectives. Clin. Applied Immunol. Rev., 4: 189-204.
CrossRefDirect Link - Yaqoob, E., I. Hussain and S.U. Rahman, 2007. Molecular characterization by using Random Amplified Polymorphic DNA (RAPD) analysis of Salmonella enteritidis isolates recovered from avian and human sources. Pak. Vet. J., 27: 102-104.
Direct Link - Quintaes, B.R., N.C. Leal, E.M. Reis and E. Hofer, 2004. Optimization of randomly amplified polymorphic DNA-polymerase chain reaction for molecular typing of Salmonella enterica serovar Typhi. Rev. Soc. Bras. Med. Trop., 37: 143-147.
PubMed