
Z.M. Kalaba
Department of Poultry Production, Faculty of Agriculture, Mansoura University, Mansoura, Egypt
A.E. Abdel-Khalek
Department of Animal Production, Faculty of Agriculture, Mansoura University, Mansoura, Egypt
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 10 | Page No.: 1010-1018







ABSTRACT
Various hormonal treatments were used for induction of ovulation to increase fertility of rabbits. Aim of the present study was to investigate the effect of repeated GnRH injections or Intra-Vaginal Mechanical Stimulation (IVMS) at mating for six consecutive litters, on fertility of naturally mated New Zealand White rabbit does. Does failed to kid after one service were mated in the next parity. Kindling Rate (KR), Number of Total (NTB) and Live Borns (NLB), Viability Rate (VR) and average Kit Weight (AKW) at birth as well as Gestation Period Length (GPL) were recorded after kidding for six consecutive parities. Also, concentrations of Follicle Stimulating Hormone (FSH) and Luteinzing Hormone (LH) were determined after mating of the 2nd litter. Results reveled that GnRH treatment showed highest (p<0.05) KR (71.4%), NTB (6.16/litter), NLB (5.53/litter) and VR (89.8%) and shortest (p<0.05) GPL (30.93 d). However, IVMS showed highest (p<0.05) AKW (66.59 g). Highest (p<0.05) VR was at the 1st and 2nd parity (87.5 and 84.5%), lowest at the 3rd parity (73.8%) and modest between 4th-6th parity. At 2, 3 and 4 h post-mating, FSH concentration was 12.04, 12.90 and 9.58 ng mL-1, while LH concentration was 16.0, 10.38 and 9.63 ng mL-1, respectively. The current study indicated that repeated injection with 0.2 mL GnRH analogue/doe (Receptal) at mating for 6 consecutive litters resulted in the best reproductive performance of rabbit does, leading to improving fertility of NZW does.
PDF Abstract XML References Citation
Received: May 25, 2011;
Accepted: July 02, 2011;
Published: September 05, 2011
How to cite this article
Z.M. Kalaba and A.E. Abdel-Khalek, 2011. Effect of Repeated GnRH Injection and Intra-vaginal Mechanical Stimulation on Reproductive Performance of Doe Rabbits. Asian Journal of Animal and Veterinary Advances, 6: 1010-1018.
DOI: 10.3923/ajava.2011.1010.1018
URL: https://scialert.net/abstract/?doi=ajava.2011.1010.1018
DOI: 10.3923/ajava.2011.1010.1018
URL: https://scialert.net/abstract/?doi=ajava.2011.1010.1018
INTRODUCTION
In rabbits, ovulation is a neuroendocrine reflex that is physiologically induced at the condition of natural breeding. The mechanisms whereby rabbit does become spontaneous ovulators are still unclear, but are likely associated to factors interfering with the control of the gonadal axis involving the hypothalamic centers responsible for GnRH release (Theau-Clement et al., 2008). The application of some of the exogenous hormonal substances may cause fertility disorders in does. For example, equine Chorionic Gonadotropin (eCG) is known to reduce fertility due to immunogenicity if used routinely (Rebollar et al., 2006).
Ovulation has to be induced artificially in does by intramuscular injection of either Gonadotropin-releasing Hormone (GnRH), or its synthetic analogues (Moce et al., 2003; Rommers et al., 2004; Quintela et al., 2004). Highly potent, long-acting synthetic analogue of GnRH have been produced and is available commercially (e.g., gonadoreline, buserelin, fertirelin acetate, desloreline and fertagyl. These compounds proved a new tool for inducing endogenous Follicle-Stimulating Hormone (FSH) and Luteinzing Hormone (LH) release and induce ovulation in all mammalian species (Taponen et al., 2003). In this concern, Bonanno et al. (2008) induced ovulation in rabbits at Artificial Insemination (AI) by intramuscular (i.m) injection of 0.8 μg of synthetic GnRH (buserelin).
In addition, ovulation can be induced by the sight of a sexually active male, by mating with a vasectomized male, a female mounting another female (Laborda et al., 2008). Also, mechanical stimulation of rabbit vagina could be used as a tool of increasing fertility as a routine work in rabbit farms in Egypt. Unfortunately, there is no information in the literature on the effect of intravaginal mechanical stimulation on reproductive performance of rabbits.
Therefore, the objective of this study was to evaluate the reproductive performance of NZW rabbit does at 6 consecutive litters as affected by the repeated stimulation of ovulation by GnRH injection as a tool to mimic hypothalamic stimulation for multiple asynchronous ovulations, or intra-vaginal mechanical stimulation at mating.
MATERIALS AND METHODS
Animals and management: The experimental work was carried out in a private Rabbit Production Farm in Zian location, Dakahlyia governorate, Egypt, during the period from August 2009 to March 2010. Total of 63 virgin New Zealand White (NZW) rabbit does (7-8 months of age and 3.5-3.85 kg body weight) as well as 18 fertile NZW rabbit bucks (10-13 months of age and 4-4.25 kg body weight) were used in the present study. All animals were healthy and clinically free of external parasites.
All does and bucks were housed in a naturally ventilated building and kept individually in flat deck cages (50x60x40 cm) only for does equipped with automatic water dispensers and internal nest boxes. Animals received a commercial diet containing 10.5 MJ digestible energy per kg, 18.5% crude protein and 12.5% crude fiber according to their physiological condition. Water was provided ad libitum all over the experimental period. Average of ambient temperature during the experimental period ranged from 15.4°C in December to 30.0°C in August, while relative humidity ranged from 65% in November to 81% in August.
Does and bucks were kept under the same management system. Cages and nest boxes were cleaned and disinfected regularly before each kindling. Light in the rabbit farms was controlled to allow 14-16 h per day during the period of the study. Urine and feces dropped from the cages on the floor were cleaned every day in the morning.
Experimental design: The experimental rabbit does were divided into similar three treatment groups (21 in each) according to their live body weight. Does in the 1st group (control) were injected with 0.2 mL saline solution at mating (without treatment). Does in the 2nd group (GnRH) were intramuscularly injected at mating with 0.2 ml/doe of GnRH analogue (Receptal, Intervet International B.V. Boxmeer-Holland, each ml of Receptal contained 0.0042 mg Buserelin acetate equivalent to 0.004 mg Buserelin). In the 3rd group, vagina of each doe was stimulated mechanically immediately before mating by thermometer (IVMS). All treatments were applied at mating for six consecutive litters.
Reproductive management: On day of mating, receptive doe was transferred to the cage of buck and returned back to her own cage after mating. On day 27 of mating, the nest boxes were supplied with wooden straw to help the doe in preparing a worm comfortable nest for the kits of her litter. Does were mated 10-12 days after parturition of each litter. At the same time, does failed to kid from the previous mating in each treatment group were mated. After parturition, the size of the litter (total and live borns at birth) was recorded and rabbit kits were weaned up to the age of 35 days.
Blood samples and hormonal assay: Blood samples were taken from does in each group at 2, 3 and 4 h after mating. Blood samples were collected by puncture of the ear vein in heparinized clean test tubes and immediately centrifuged (4000 g) for 15 min. Then, plasma was separated and stored at -20°C until assay. Plasma concentrations of FSH and LH were determined in duplicate by radioimmunoassay (RIA), using AFP- 472176 and AFP-3120489 antibodies for FSH and LH, respectively, provided by the National Institute of Diabetes and Digestive and Kidney Diseases (Harbor UCLA Medical Center, CA, USA). The dilutions antibody used were 1:45,000 and 1:1,500,000 for FSH and LH, respectively. Hormones were iodinated with 125I, using the chloramine-T- method (Greenwood et al., 1963). The assay sensitivity was 1.0 and 0.05 ng mL-1 for FSH and LH, respectively. The intra-assay coefficients of variation, estimated from plasma pool measured 8 times in the same assay, were <8% for both hormones.
Data recorded: Within 12 h after kindling of each litter, total number of kits (live and dead) was calculated, dead kits were removed and then size and weight of live kits were recorded. Kindling rate (number of kindled does per number of mated doesx100), viability rate (number of live per total number of kitsx100) and length of gestation period (day) were calculated.
Statistical analysis: Data obtained in this study were statistically analyzed using computer program of SAS (2004). Factorial ANOVA (GLM) was used to perform the effect of treatment, litter number and their interaction different traits studied. Also, the effect of treatment, time post-mating and their interaction only for the 2nd litter on concentration of FSH and LH in blood plasma was performed. Multiple rang test (Duncan, 1955) was employed to test statistical differences among means at (p<0.05). Only the significant interactions were included in the discussion.
RESULTS AND DISCUSSION
Reproductive and litter size of doe rabbits
Effect of treatment: Results presented in Table 1 reveal that stimulation of ovulation by GnRH showed significantly (p<0.05) the higher kindling rate (71.4%), litter size per doe as a total (6.16/litter) and live kits (5.53/litter), viability rate (89.8%), kit weight (64.11 g) and litter weight (345.5 g as compared to 48.4%, 5.67 and 3.91/litter, 70.0%, 60.83 g and 1.76/litter in the controls, respectively). The differences between GnRH and IVMS groups in all traits weight were insignificant, but all values were higher for GnRH than IVMS treatment. In general, reproductive index was the highest (3.95) for GnRH-treated does, moderate in IVMS (3.46) and the lowest in the control (1.89).
Localization of GnRH within the hypothalamus has been well documented in rabbits (Arimura et al., 1973) and many investigators reported improvement in kindling of rabbit does as affected by exogenous GnRH.
| Table 1: | Least square means and standard errors of means (SEM) of productive and reproductive traits of doe rabbits as affected by stimulation treatment and does parity |
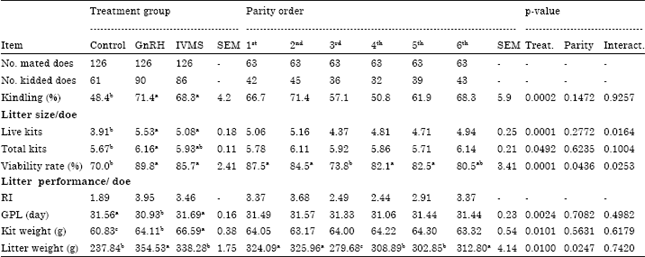 | |
| a, b and c: Means denoted within the same raw for group or parity having different superscripts are significantly different at p≤0.05. IVMS: Intravaginal mechanical stimulation, RI: Reproductive index. GPL: Gestation period length | |
In nearly similarity with the present results, Zapletal et al. (1990) reported that the total annual fertility rate was higher (73.02%, p<0.05) in rabbit group treated with lecirelin (GnRH) but with higher litter size as a total and live borns (9.82 and 9.70, respectively) than those obtained in the present study. It has been shown that GnRH mediates the hypothalamic control of pituitary gonadotropin secretion and biosynthesis (Dekel et al., 1988) to induce ovulation. However, the obtained improvement in kindling rate and litter size of rabbit does treated with GnRH as compared to the controls in this study was also due to the effects of GnRH on the ovary include stimulation of oocyte maturation (Dekel et al., 1988; Yang et al., 1995).
In addition, GnRH may interfere with the stimulatory effects of exogenous gonadotropin on follicular development, corpus luteum establishment and oocyte maturation (Zanagnolo et al., 1996). Some studies have shown that GnRH and its analogues also exert a direct effect on gonadal function, influencing rabbit oocyte maturation both in vivo and in vitro (Yang et al., 1995; Yoshimura et al., 1992). It has been suggested that GnRH induces oocyte maturation via activation of specific GnRH receptors on granulosa cells (Koves et al., 1989). Exposure to GnRH stimulates prostaglandin synthesis in preovulatory follicles (Wong and Richards, 1992). Increasing concentration of prostaglandin plays an important role in oocyte maturation (Calder et al., 2001). According to these studies, GnRH treatment increased number of kits per litter and decreased kit weight at birth (Table 1), which is in good agreement with the finding of Zapletal and Pavlik (2008), who reported that the conception rates ranged from 10.0 to 89.5% for different GnRH doses.
On the other hand, buserelin or triptorelin added to seminal doses (Viudes-de-Castro et al., 2007) have been shown to be valid methods for inducing ovulation in rabbit does submitted to artificial insemination. However, the hormonal doses needed were high and the kindling rates obtained were similar or lower than those for the control group. In agreement with the results of GnRH injection intramuscularly, Ondruska et al. (2008) found that supplementation of insemination dose at AI with GnRH-Lecirelinum (7.5 μg) increased kindling rate (72.09%) as compared to 62.74% in the control group (administered i.m. with 2.5 μg same synthetic GnRH immediately after AI). However, viability rate was not affected significantly by GnRH treatment (Ondruska et al., 2008). The present results are nearly similar for kindling rate with improving viability rate inspite the marked differences in level and location of GnRH treatment. In this respect, Quintela et al. (2004) used GnRH-buserelin in considerable higher concentrations for intravaginal use (10 up to 20 times) compared with intramuscular control (0.8 μg per female). Differences in kindling rate among treated groups were not statistically significant in their experiments. After intravaginally application of 20-times higher concentration of buserelin they obtained significantly higher prolificacy in experimental females compared with the control (11.7 young and 9.4 kits per litter, respectively). In conclusion, buserelin was evaluated to determine potential negative effects on the sperm characteristics and no detrimental effects were found on sperm motility, viability or acrosomal integrity during 24 h of sperm incubation at 16°C. According to the previous results and those obtained in our study, GnRH treatment to induce ovulation in rabbits is depending on type of analogue, does of GnRH and location of treatment.
The obtained results clearly demonstrated that the GnRH analogue (buserelin) i.m. injected was extremely efficient, significantly increasing kindling rate and viability rate of kits at birth for 6 successive litters in comparison with IVMS and control.
Effect of parity: Results in Table 1 show that only viability rate of kits and litter weight at birth significantly (p<0.05) affected by parity. However, kindling rate, litter size (total and live), kit weight and gestation period were not affected significantly by parity. Generally, most of the studied traits were the highest at the 2nd parity and the lowest at the 3rd one.
Inspite the insignificant differences in kindling rate at different parities, rabbit does showed pronounced increase in kindling rate from 66.7 % at the 1st parity to 71.4% at the 2nd one and decreased to 57.1% at the 3rd parity, then insignificantly increased with progress of parity up to 68.3%, at the 6th parity. The noted increase in kindling rate and litter size at the 2nd compared with 1st parity may be due to linkage between pseudo-pregnant does and parity. In this respect, Theau-Clement et al. (2008) recorded increasing the percentage of pseudo-pregnant does in primiparous (31.2%) than in secondiparous does (12.2%, p<0.001).
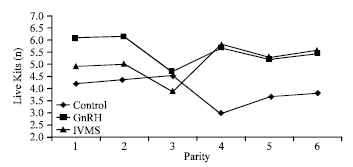 | |
| Fig. 1: | Interaction between treatment and parity for number of live kits at birth |
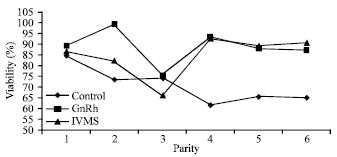 | |
| Fig. 2: | Interaction between treatment and parity for viability rate at birth |
Yet, the components of litter size have been studied by genetic (Argente et al., 1997) and phenotypic (Santacreu et al., 1992) points of view.
Analysis of variance revealed that the effect of interaction between treatment and parity was significant (p<0.05) only on number of live kits/litter and viability rate, reflecting nearly similar trend of change (p<0.05) in GnRH and IVMS groups as compared to control group (Fig. 1, 2).
At the 1st and 2nd parities, number of live kits (Fig. 1) was significantly (p<0.05) greatest (6.09 and 6.14/litter), followed by IVMS (4.90 and 5.00/litter), and the lowest in control group trend (4.19 and 4.33/litter). At the 3rd parity, live kits decreased in GnRH and IVMS groups to the lowest values (4.67 and 3.90/litter, respectively) versus slight increase to the highest values (4.52/litter) in control group, but the differences were not significant. At the 4th parity, live kits showed marked increase to 5.67/litter in GnRH and to 5.81/litter in IVMS groups, being significantly higher than 2.95/litter in control group. At the 5th and 6th parities, live kits were significantly (p<0.05) higher in both GnRH (5.19 and 5.43/litter) and IVMS (5.28 and 5.57/litter) groups than in control group (3.67 and 3.81 /litter).
Regarding viability rate of kits, results illustrated in Fig. 2 show that at the 1st parity, viability rate was 84.69, 86.69 and 89.39% in control, GnRH and IVMS groups, respectively and the differences were not significant. Then, it showed similar trend to that of number of live/litter at subsequent parities. Viability rate was significantly (p<0.05) the highest at the 2nd, 3rd and 4th parities in GnRH group (99.21, 75.55 and 93.43%, respectively) and at the 5th and 6th parities in IVMS group (89.32 and 90.43%, respectively). Yet, kits of the control group showed significantly (p<0.05) the lowest viability at all parities, except for the 3rd one.
Based on the previous results of parity effect, GnRH treatment or IVMS at mating for 6 successive litters had beneficial effects on fertility and viability of kits at birth.
Concentration of gonadotrophins: Results in Table 2 show that stimulation of ovulation by GnRH led to significant (p<0.05) increase in concentration of FSH and LH in blood plasma to 16.71 and 16.57 ng mL-1 as compared to 5.79 and 9.23 ng mL-1 in the control, respectively. However, IVMS significantly (p<0.05) increased only FSH concentration as compared to the control (14.02 vs. 5.79 ng mL-1).
| Table 2: | Effect of type of stimulation and sampling time after mating on concentration of FSH and LH in blood plasma of rabbit does |
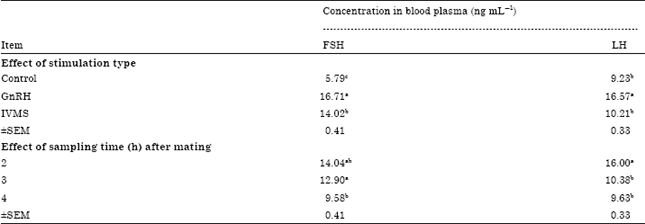 | |
| a, b and c: Means denoted within the same column for each effect having different superscripts are significantly different at p<0.05. IVMS: Intravaginal mechanical stimulation | |
As affected by mating time, FSH concentration significantly decreased 4 h post-mating, however, LH concentration showed significant decrease 3 h post-mating. At 2, 3 and 4 h post-mating, FSH concentration was 14.04, 12.90 and 9.58 ng mL-1, while LH concentration was 16.0, 10.38 and 9.63 ng mL-1, respectively.
The insignificant interaction between treatment and mating time on concentration of both FSH and LH was reflected in the highest concentration in GnRH treatment 2 h post-mating.
In this study the changes occurring in plasma LH and FSH concentrations 2, 3 and 4 h post- mating may explain ovulation rate and litter size. The reproductive hormone profile of FSH and LH in rabbit does have been studied at mating by Ubilla et al. (1992, 2000 a,b). The FSH may influence the follicular growth; meanwhile LH may determine the number of breakage of pre-ovulatory follicles. According our results, there was association between FSH and LH peaks. Similar findings were reported by Muelas et al. (2008) and Rodriguez et al. (1989).
Many authors observed that the highest LH and FSH concentrations reached 60-90 min after exogenous administration of GnRH, decreasing to basal concentrations 4-5 h later (Mills and Gerardot, 1984; Rodriguez et al., 1989; Rebollar et al., 1997).
In agreement with our results concerning the FSH, Muelas et al. (2008) did not find change in FSH concentration 48 h before or after mating and 2 h after mating. Also, Ubilla et al. (2000b) did not find any difference in FSH concentrations between 48 h before and 7 h after AI. However, the present trend of change in LH was comparable with that observed on rabbits by Muelas et al. (2008), who found significant differences among LH plasma concentrations measured 48 h before or after the mating (5.1 and 5.4 ng mL-1, respectively) vs. 2 h after the mating (29.0 ng mL-1). Similar trend was observed by Ubilla et al. (2000a). In accordance with the marked increase in LH concentration in GnRH-treated does, Rodriguez et al. (1989) showed that LH level sharply increased after 15 min of GnRH challenge, being 200-1000%, depending on sexual receptivity of does and GnRH dose. However, Muelas et al. (2008) found that the LH levels were 4-6 fold higher 2 h after the mating in does with high ovulation rate, which may explain the highest litter size of GnRH-treated does as compared to the control group. In addition, This may explain the higher ovarian stimulation and pituitary responsiveness after GnRH biostimulation. The pronounced reduction in litter size with advancing parity may be related to that the response of circulating FSH to active immunization against GnRH. In several species, active immunization against GnRH consistently decreased LH secretion to undetectable levels (Garza et al., 1986; Ondruska et al., 2008).
CONCLUSION
Based on the foregoing results, stimulation of ovulation in rabbit does by 0.2 mL GnRH (receptal)/doe at mating for 6 consecutive litters can be successfully used to improve fertility of rabbits and their litter weight. Also, intra-vaginal mechanical stimulation at mating could have some advantages on rabbit fertility by avoiding the intramuscular application of GnRH analogues and probably reducing the sanitary risk derived from incorrect use of needles. Further studies are necessary for determining an optimal level of GnRH analogue at different times of mating in relation with the physiological status of rabbit does at the moment of mating.
REFERENCES
- Argente, M.J., M.A. Santacreu, A. Climent, G. Bolet and A. Blasco, 1997. Divergent selection for uterine capacity in rabbits. J. Anim. Sci., 75: 2350-2354.
Direct Link - Calder, M.D., A.N. Caveney, M.E. Westhusin and A.J. Watson, 2001. Cyclooxygenase-2 and prostaglandin E(2) (PGE(2)) receptor messenger RNAs are affected by bovine oocyte maturation time and cumulus-oocyte complex quality and PGE(2) induces moderate expansion of the bovine cumulus in vitro. Biol. Reprod., 65: 135-140.
PubMedDirect Link - Dekel, N., O. Lewysohn, D. Ayalon and E. Hazum, 1988. Receptors for gonadotropin releasing hormone are present in rat oocytes. Endocrinology, 123: 1205-1207.
PubMed - Garza, F. Jr., D.L. Thompson Jr., D.D. French, J.J. Wiest and R.L. St. George et al., 1986. Active immunization of intact mares against gonadotropin-releasing hormone: Differeatid effects on secretion of luteinizing hormone and follicle stimulating hormone. Biol. Reprod., 35: 347-352.
PubMed - Greenwood, F.C., W.M. Hunter and J.S. Glover, 1963. The preparation of 131I-labelled human growth hormone of high specific radioactivity. Biochem. J., 89: 114-123.
PubMed - Koves, K., P.E. Gottschall and A. Arimura, 1989. Gonadotropin-releasing hormone binding sites in ovaries of normal cycling and persistent-estrus rats. Biol. Reprod., 41: 505-511.
PubMed - Mills, T.M. and R.J. Gerardot, 1984. Dissociation of copulation from ovulation in pregnant rabbits. Biol. Rep., 30: 1243-1252.
PubMed - Moce, E., R. Lavara and J.S. Vicente, 2003. Effect of an asynchrony between ovulation and insemination on the results obtained after insemination with fresh or frozen sperm in rabbits. Anim. Reprod. Sci., 75: 107-118.
PubMed - Quintela, L.A, A.I. Pena, M.D. Vega, J. Gullon and M.C. Prieto et al., 2004. Ovulation induction in rabbit does submitted to artificial insemination by adding buserelin to the seminal dose. Reprod. Nutr. Dev., 44: 78-88.
CrossRefDirect Link - Rebollar, P.G., J.M. Alvarino, J.C. Illera and G. Silvan, 1997. Effect of gonadoreline and naloxone on induction of ovulation and plasma LH in rabbit. J. Physiol. Biochem., 53: 205-210.
PubMed - Rebollar, P.G., A. Milanes, N. Pereda, P. Millan and P. Cano et al., 2006. Oestrus synchronisation of rabbit does at early post-partum by doe-litter separation or ECG injection: Reproductive parameters and endocrine profiles. Anim. Reprod. Sci., 93: 218-230.
PubMed - Rodriguez, J.M., C. Agrasal and A. Esquifino, 1989. Influence of sexual receptivity on LH, FSH and prolactin release after GnRH administration in female rabbits. Anim. Reprod. Sci., 20: 57-65.
Direct Link - Rommers, J.M., R. Meijerhof, J.P. Noordhuizen and B. Kemp, 2004. The effect of level of feeding in early gestation on reproductive success in young rabbit does. Anim. Reprod. Sci., 81: 151-158.
PubMed - Taponen, J., P. Hjerppe, M.E. Kopra, H. Rodriguez, T. Katila and H. Kindahl, 2003. Premature prostaglandin F2α secretion causes luteal regression in GnRH-induced short estrous cycles in cyclic dairy heifers. Theriogenology, 60: 379-393.
CrossRefPubMedDirect Link - Theau-Clement, M., G. Bolet, L. Fortun-Lamothe, G. Brecchia and C. Boiti, 2008. High plasmatic progesterone levels at insemination depress reproductive performance of rabbit does. Proceedings of the 9th World Rabbit Congress, June 10-13, Verona, Italy, pp: 459-463.
Direct Link - Ubilla, E., J.M.R. Alvarino, A. Esquifino and C. Agrasal, 1992. Effects of induction of parturition by administration of a prostaglandin F2α analogue in rabbits: Possible modification of prolactin, LH and FSH secretion patterns. Anim. Reprod. Sci., 27: 13-20.
Direct Link - Ubilla, E., P.G. Rebollar, D. Pazo, A.I. Esquifino and J.M. Alvarino, 2000. Effects of doe-litter separation on endocrinological and productivity variables in lactating rabbits. Livestock Prod. Sci., 67: 67-74.
CrossRefDirect Link - Ubilla, E., P.G. Rebollar, D. Pazo, A. Esquifino and J.M.R. Alvarino, 2000. Pituitary and ovarian response to transient doe-litter separation in nursing rabbits. J. Reprod. Fert., 118: 361-366.
PubMedDirect Link - Viudes-de-Castro, M.P., R. Lavara, F. Marco-Jimenez, C. Cortell and J.S. Vicente, 2007. Ovulation induced by mucosa vaginal absorption of buserelin and triptorelin in rabbit. Theriogenology, 68: 1031-1336.
PubMed - Wong, W.Y. and J.S. Richards, 1992. Induction of prostaglandin H synthase in rat preovulatory follicles by gonadotropin-releasing hormone. Endocrinology, 130: 3512-3521.
PubMed - Yang, B.C., T. Uemura and H. Minaguchi, 1995. Effects of gonadotropin releasing hormone agonist on oocyte maturation, fertilization and embryonal development in mice. J. Assist. Reprod. Genet., 12: 728-732.
PubMed - Yoshimura, Y., Y. Nakamura M. Ando, M. Jinno, T. Nanno and T. Oda, 1992. Protein kinase C mediates gonadotropin-releasing hormone agonist-induced meiotic maturation of follicle-enclosed rabbit oocytes. Biol. Reprod., 47: 118-125.
PubMed - Zanagnolo, V., A.M. Dharmarajan, J. Hesla and E.E. Wallach, 1996. Effects of a gonadotropin-releasing hormone analog on rabbit ovarian function. Endocrinology, 137: 5400-5406.
PubMed - Zapletal, D. and A. Pavlik, 2008. The effect of lecirelin (GnRH) dosage on the reproductive performance of nulliparous and lactating rabbit does. Anim. Reprod. Sci., 104: 306-315.
CrossRefPubMedDirect Link - Zapletal, D., V. Adamec and D. Klecker, 1990. Effect of GnRH hormones application on performance of artificial inseminated rabbits (Oryctolagus cuniculus) during year. Acta Univ. Agric. Silviculturae Mendeleianae Brunensis, 51: 123-132.
Direct Link