
S.N. Hlophe
Aquaculture Research Unit, Faculty of Science and Agriculture, School of Agricultural and Environmental Sciences, University of Limpopo (Turfloop Campus), Private Bag X1106, Sovenga, 0727, South Africa
N.A.G. Moyo
Aquaculture Research Unit, Faculty of Science and Agriculture, School of Agricultural and Environmental Sciences, University of Limpopo (Turfloop Campus), Private Bag X1106, Sovenga, 0727, South Africa
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 10 | Page No.: 1001-1009







ABSTRACT
The culture of macrophagous fish that naturally feed on plant diets may be the solution to reduce the current dependence on fishmeal. Fishmeal is not only expensive but its supply is not always guaranteed. This study focuses on the growth performance, gastric evacuation rate, gastric transit time and carcass composition of Tilapia rendalli fed fresh plants, to determine the extent to which T. rendalli can utilise fresh plants. Kikuyu grass (Pennisetum clandestinum), cabbage (Brassica oleracea), duckweed (Lemna minor), vallisneria (Vallisneria aethiopica) and fishmeal pellets (control) were offered ad libitum to duplicate groups of T. rendalli for 224 days. Specific Grow Rate (SGR), Protein Efficiency Ratio (PER) and Food Conversion Ratio (FCR) were used to determine the growth performance. Fish fed kikuyu grass attained a significantly (p<0.05) higher SGR and a better FCR than those fed the other plant diets. Fish fed vallisneria lost weight. The serial slaughter method showed that vallisneria was evacuated significantly (p<0.05, ANCOVA) faster and was eaten in significantly (p<0.05) higher quantities than the other diets. Kikuyu grass was evacuated much more slowly and eaten in lesser amounts. The low energy content (14.74 MJ kg-1) of vallisneria may explain its faster evacuation and high consumption levels. Fish fed kikuyu grass had higher protein levels, higher omega-3 fatty acids (25.13%) and higher mineral content than those fed the other experimental diets. Fishmeal fed fish had the lowest content of the omega-3 fatty acids (2.52%). T. rendalli performed better when fed plant diets with higher protein and energy contents.
PDF Abstract XML References Citation
Received: June 01, 2011;
Accepted: August 06, 2011;
Published: September 05, 2011
How to cite this article
S.N. Hlophe and N.A.G. Moyo, 2011. The Effect of Different Plant Diets on the Growth Performance, Gastric Evacuation Rate and Carcass Composition of Tilapia rendalli. Asian Journal of Animal and Veterinary Advances, 6: 1001-1009.
DOI: 10.3923/ajava.2011.1001.1009
URL: https://scialert.net/abstract/?doi=ajava.2011.1001.1009
DOI: 10.3923/ajava.2011.1001.1009
URL: https://scialert.net/abstract/?doi=ajava.2011.1001.1009
INTRODUCTION
Traditionally, fishmeal has been used as the major protein source in fish feeds because of its nutritional value and palatability (Nguyen et al., 2009). However, due to the limited world supplies and increasing price of fishmeal, the majority of research on fishmeal replacement with alternative proteins in fish diets has focused on the use of protein derived from plant sources (Enami, 2011). Alternative plant protein sources are generally cheaper compared to animal protein sources. High protein levels are a prerequisite in selecting alternative plant sources, as dietary protein affects the growth performance in tilapia (Musuka et al., 2009). Soybean Meal (SBM) is generally considered to be one of the best readily available plant protein sources in terms of its protein quality and amino acid profile (El-Sayed, 2006). Studies by Kang’ombe and Brown (2008) have shown that SBM can be used in tilapia feeds.
Aquatic macrophytes are important as a food source, providing shelter as well and nutrient recycling (Uka and Chukwuka, 2011). Tilapia rendalli utilizes macrophytes in the wild and it is well adapted to handling plant diets because of the presence of pharyngeal mill to break down plant matter (El-Sayed, 2006). The use of fresh plants as fish feeds has been investigated by several researchers. When fed only on fresh plants in closed systems, growth performance is usually poor; Setlikova and Adamek (2004) reported slow growth of O. niloticus fed Potamogeton pectinatus and Elodea canadensis and negative growth when offered Spirodela polyrhiza and Myriophyllum spectrum. In another study, Okeyo and Montgomery (1992) reported negative growth in O. aureus when fed three aquatic macrophytes (Elodea canadensis, Myriophylum spicatum and Potamogeton gramineous). However, improved growth performance was reported by Chowdhury et al. (2008) in pond reared Oreochromis niloticus when supplemented with duckweed. Similarly, favourable growth rates were reported when T. rendalli was fed Napier grass (Pennisetum purpureum) (Chikafumbwa, 1996).
One of the primary objectives of this study was to evaluate the capacity of T. rendalli to utilise readily available plants. The plants under investigation are a common lawn grass; kikuyu (Pennisetum clandestinum), an abundant vegetable, cabbage (Brassica oleracea), a common floating weed; duckweed (Lemna minor) and vallisneria a submerged macrophyte (Vallisneria aethiopica).
Feeding rate and gastric evacuation rate of T. rendalli grazing on these plants was also determined. There is paucity of information on the interactions between feeding rate, gastric evacuation rate and plant energy content in T. rendalli. An understanding of these interactions is important in the development of a feeding strategy for T. rendalli when it is cultured in extensive low technology culture systems.
MATERIALS AND METHODS
An experiment to determine the utilisation of kikuyu, cabbage, duckweed and vallisneria by T. rendalli was conducted for 224 days between 22 February and 13 October 2010 at the Aquaculture Research Unit of the University of Limpopo, South Africa. A completely randomised design experiment with two replications per diet was set up (total of 10 tanks). Each tank was stocked with 25 mixed-sex sub-adult T. rendalli of mean weight 64±2 g. Kikuyu grass, vallisneria and duckweed were harvested fresh each morning and cabbage was bought fresh from a local market. All plant feeds with the exception of duckweed were chopped into 1 cm pieces. Diets were offered ad libitum each morning. The amount of food consumed was recorded daily for each tank. All fish from each tank were weighed once every four weeks. Daily measurements were taken for temperature (°C), dissolved oxygen (mg L-1) and pH, using a handheld YSI (556 MPS) multi-meter. Growth performance was determined using: Specific Growth Rate (SGR) was calculated according to Winberg (1965):
All diets were analysed for crude protein, crude fat, crude fibre, carbohydrates, energy, ash, moisture and minerals, following the procedures stipulated by the Association of Official Analytical Chemists (AOAC International, 2003). Amino acid determination was done using a Beckman Amino Acid Analyser System 6300.
Gastric evacuation rate of the different experimental plant diets, kikuyu grass, vallisneria, cabbage, duckweed and fishmeal pellets (control) were determined in T. rendalli. Fish (50±5 g) were stocked in 1 m3 fibre glass tanks at 40 fish per tank. This was duplicated for each diet. The serial slaughter method (Windell, 1968) was used to determine gastric evacuation rate. Two fish per tank (4 fish in total per diet) were sampled and the amount of food remaining in the stomach (dry weight) was determined at 4 h intervals.
The gastric evacuation rate for the different plant diets was expressed by the exponential equation:
where, St is weight of stomach content; So is weight of meal eaten; t is time in h and b is constant.
At the end of the feeding experiment, a sample of four fish was taken from each diet, gutted, deboned and de-scaled. Fish samples were then frozen at -20°C until they were analysed for proximate composition of the carcass. Gas Chromatography was used for fat analysis.
Statistical analysis: One-way Analysis of Variance (ANOVA) on the Statistical Package and Service Solutions (SPSS version 17.0) was used to determine significant differences (p<0.05) on SGR, FCR, PER, standing crop, yield increment, total production and gastric transit time in T. rendalli. Tukey’s test was used to separate means where significant differences (p<0.05) were found. The Analysis of Covariance (ANCOVA) was used to determine significant (p<0.05) differences in the gastric evacuation rate of T. rendalli fed the experimental diets.
RESULTS
The SGR, FCR and yield (g m-3) varied significantly (p<0.05, ANOVA) among the diets offered to T. rendalli. Fish fed fishmeal pellets had the highest SGR, FCR and yield (g m-3), while those fed vallisneria exhibited negative growth. Tukey’s test showed that there were no significant differences (p>0.05) in the PER and production (g m-3 day-1) of fish fed duckweed and cabbage. However, significant differences (p<0.05, ANOVA) were observed in the PER and production (g m-3 day-1) of fish fed the other diets. Similarly, fish fed fishmeal pellets had the highest PER and production (g m-3 day-1). Fish fed vallisneria lost weight and hence the SGR, FCR, PER, yield and production were negative (Table 1). T. rendalli fed vallisneria consistently lost weight and its feeding was terminated after 60 days. The mean water temperature was 25±2°C, dissolved oxygen 6.24-7.1 mg L-1, ammonia ranged between 0.62-0.87 ppm.
The proximate composition of the experimental diets showed that among the plant diets, duckweed had the highest protein content followed by kikuyu grass, vallisneria and cabbage had the lowest protein level (Table 2). The control (fishmeal pellets) had the highest protein content when compared to all the plant diets. Kikuyu grass had the highest energy content, followed by the fishmeal pellets (control) then duckweed and cabbage. Vallisneria had the lowest energy content.
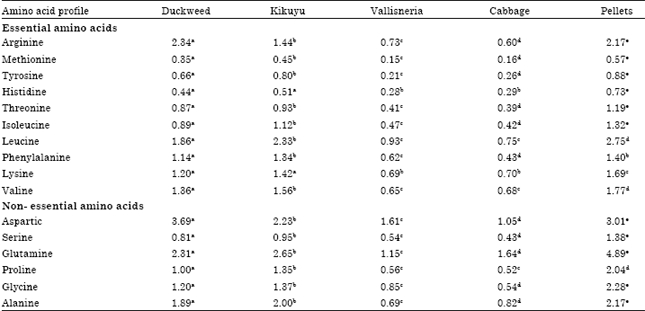
The sulphur containing amino acid methionine was the limiting amino acid in all the diets (Table 3). Methionine levels were significantly (p<0.05) different in all the test diets.
| Table 1: | Growth performance of Tilapia rendalli fed on different experimental diets. Data are Mean values±SE |
 | |
| Values in the same column with different superscripts are significantly different (p<0.05, ANOVA) | |
| Table 2: | Proximate composition of the experimental diets expressed as percent dry matter |
 | |
| Table 3: | Amino acid profile of the experimental diets expressed as percent protein |
 | |
| Values in the same row with different superscripts are significantly different (p<0.05) | |
The lowest (0.15%) levels were recorded in vallisneria whilst kikuyu grass had the highest (1.44%) levels among the plant diets. The lysine content was also significantly different in the diets. Kikuyu grass had the highest (1.42%) levels, followed by duckweed, cabbage and vallisneria had the lowest (0.69%) levels. Arginine was only limiting in cabbage and not limiting in the other diets. Kikuyu grass had a better array of all the essential amino acids than the other plant diets.
There was a significant difference (p<0.05, ANOVA) in the feeding rate of T. rendalli when fed the test diets (kikuyu, duckweed, cabbage, vallisneria or fishmeal pellets). Vallisneria was eaten in higher quantities (210 g day-1) than the other test diets, whilst fishmeal pellets were eaten in smallest quantities (44.8 g day-1). However, post-hoc analysis (Tukey’s test) showed that the mean feeding rate was not significantly different (p>0.05) between kikuyu grass and the commercial fishmeal pellets, as well as between kikuyu grass and duckweed. Similarly, the feeding rate of T. rendalli between duckweed, cabbage and vallisneria was not significantly different (Table 4).
Gastric evacuation rates of the experimental diets varied significantly (p<0.05, ANCOVA). Vallisneria was evacuated faster (b’ = -0.0033) followed by cabbage (b’ = -0.0029), duckweed (b’ = -0.0022), kikuyu grass (b’ = -0.0021) and fish fed fishmeal pellets were evacuated more slowly (b’ = -0.0016). Gastric transit time was significantly different (p<0.05, ANOVA) between the diets. However, the GTT was not significantly different (p>0.05, Tukey) between kikuyu grass and duckweed. Gastric transit time was shortest in fish fed vallisneria (2.21 h), followed by those fed cabbage, duckweed, then kikuyu grass. Fishmeal pellets were in the gastric tract for the longest period (3.51 h) (Table 4).
Carcass composition analysis revealed the amount of protein and calcium was not significantly different (p<0.05, ANOVA) among fish fed the experimental diets (Table 5). It is important to note that fish fed kikuyu grass had the higher protein (71.55%) and calcium (6.76%) levels. There were significant differences (p<0.05, ANOVA) in the energy, fat, moisture and phosphorous levels of the fillet of fish fed the different experimental diets (Table 5). Duckweed fed fish had the second highest amount of these nutrients followed by fish fed cabbage. The highest energy and fat content in the carcass was recorded for fish fed cabbage, duckweed and kikuyu grass, respectively.
Fish fed the experimental plant diets had higher levels of the healthy omega-3 fatty acids than fish fed fishmeal pellets (Table 6). Kikuyu grass produced the highest amount of omega-3 fatty acids in the fillet, followed by fish fed cabbage, duckweed and then vallisneria.
| Table 4: | Feeding rates, Gastric Evacuation Rate (GER), Gastric Transit Times (GTT) and time for complete evacuation (±SE) in Tilapia rendalli fed the experimental diets |
 | |
| Values in the same column with different superscripts are significantly different (p<0.05) | |
| Table 5: | Percentage carcass composition of fish fed the different plant diets |
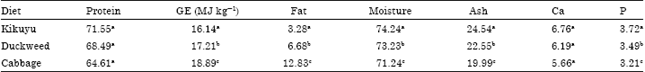 | |
| Values in the same column with different superscripts are significantly different (p<0.05) | |
| Table 6: | Percent fatty acids in the fillet of fish fed the different experimental diets |
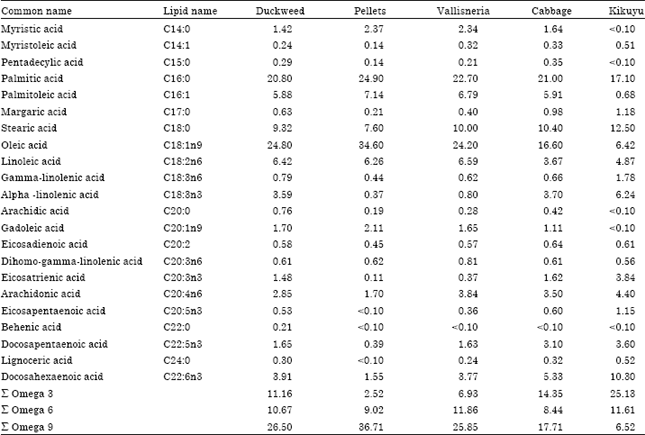 | |
Fish fed fishmeal pellets had the lowest amount of omega-3 fatty acids. The ratio of omega 3:6 fatty acids was highest in kikuyu, followed by cabbage, duckweed, vallisneria and fishmeal had a lower ratio of omega 3:6 fatty acids.
DISCUSSION
T. rendalli fed kikuyu grass performed better than fish fed the other plant diets. This was followed by fish fed duckweed and cabbage, respectively. Fish given vallisneria lost weight. Kikuyu grass fed fish attained higher SGR, PER and lower FCR than fish fed the other plants. There are several dietary factors that affect the growth performance of fish, these include the protein, energy, lipid and fibre content as well as the presence of anti-nutritional factors.
Generally, diets with high protein content produce better growth rates. Duckweed had the highest level of proteins among the plant diets, however fish fed kikuyu grass performed better than those fed duckweed. The nutritional value of protein is based on the amino acid composition of the protein source and their biological availability from the protein (El-Sayed, 2006). This was confirmed in this study as kikuyu grass had superior levels of the essential and often limiting amino acids methionine and lysine and fish fed the grass performed better. This confirms that the amino acid levels of the diet are more important than the total protein value. Fishmeal pellets had higher protein content and higher proportion amino acid than all the plant diets and this accounts for the high growth rates obtained in fish fed the pellets. Similarly, Adewolu and Adoti (2010) reported better growth performance in fish fed a higher protein diet.
The specific growth rates (on wet weight basis) obtained in T. rendalli feeding on the plant diets in this study ranged from 0.13%/day in fish fed cabbage to 0.17%/day in fish fed kikuyu. These results show that T. rendalli can utilise plant based protein. The negative values obtained for SGR, FCR and PER in T. rendalli given vallisneria may be due to its low nutritive value. Vallisneria is an aquatic macrophyte that has a higher moisture content than the other plants. Furthermore this macrophyte has low protein and energy content. These results support the findings of several researchers who found that Tilapias sometimes lose weight when fed exclusively on aquatic macrophytes; Saeed and Ziebell (1986) reported negative growth for T. zillii when fed Elodea densa a submerged macrophyte. Similarly, Setlikova and Adamek (2004) reported that O. niloticus lost weight when fed Spirodela polyrhiza and Myriophylum specatum.
The energy content in the feed is another factor affecting growth performance, fish feed to satisfy their energy requirements and if the diet does not contain sufficient energy levels, protein is used for energy rather than for growth. An inadequate dietary protein to energy ratio may result in lower growth performance (Ali et al., 2008). Kikuyu grass had the highest (17.99%) energy levels, followed by fishmeal pellets (17%), duckweed (16.43%), then cabbage (16.09%) and vallisneria had the lowest levels (14.74%). The high energy levels coupled with a good amino acid balance in kikuyu grass may have resulted in more protein spared for growth than in the other plant diets. This may suggest that diets with high energy levels yield higher better growth performance than low energy diets. This is in agreement with studies conducted by Ali et al. (2008) who reported poor FCR values in O. niloticus fed diets when the energy content of the diet was reduced. In aquaculture, diets with sufficient energy levels are desired so that the energy demand for the fish should be met by none protein sources.
Another important factor known to affect the utilisation of plant diets is the fibre content of the diet. A major reason why the feeding efficiency of herbivorous fish is usually low is that fish have limited cellulose activity (Saha et al., 2006) However, fibrous compounds (except lignin) are subject to partial digestion by hydrochloric acid in the stomach especially in species which produce low HCl such as T. rendalli. This was confirmed in this study because kikuyu grass had the highest fibre (18.58%) and yet it was effectively utilised by T. rendalli. High levels of fibre in the diet also affect apparent digestibility and gastrointestinal transit rate in tilapias (Lanna et al., 2004). Diets with high levels of fibre are not desirable in practical fish feeds as they hinder nutrient digestibility, resulting in poor utilisation of the feed and low growth rates (Azaza et al., 2008). However, Dioundick and Stom (1990) reported that some amount of fibre is required for optimal utilisation of the diet and overall growth. T.rendalli being pre-adapted to utilising higher plants may be able to breakdown some of the fibre/cell wall and expose the cell contents to digestive processes. The low stomach pH also helps in the release of energy from plant cell walls (Abdel-Tawwab, 2003). Plenty surface area for digestion and absorption is also provided by the long intestines. Therefore, the amount of fibre in the diets appears to play a minor role in hindering the growth performance in T. rendalli.
The gastric evacuation rate in T. rendalli was probably influenced by the energy content in the feed. Pellets with the high energy content were evacuated slowly and the evacuation rate decreased with decreasing energy content in the feed. Similarly, vallisneria with the lowest energy content had the shortest evacuation time. These results are in agreement with De Silva and Anderson (1995) and Targett and Targett (1990) who indicated that gastric evacuation is regulated by the energy content of the food; the greater the energy content, the slower the evacuation rate. Vallisneria was eaten in larger quantities than the other experimental diets because it has a low energy content. Similarly, Riche et al. (2004) reported that the return of appetite in fish is linked gastric emptying time. This indicates that rapid evacuation leads to a more rapid increase in fish appetite leading to more consumption of the particular diet. In the case of vallisneria, where the fish lost weight, it appears that the increased rate of evacuation was not sufficient to allow for the lower energy content.
Carcass composition showed that T. rendalli fed kikuyu grass had the highest protein content of all the plant diets. This is consistent with the PER values that were also higher in kikuyu. Omega 3-fatty acids were also very high in kikuyu. On the contrary, fish fed fishmeal pellets had low omega 3-fatty acid levels. Perschbacher et al. (2010) also obtained similar results reporting high omega 3-fatty acids in fish fed a plant diet. Plants are the major source of omega 3-fatty acids. The implications of these results are that the consumption of fish fed kikuyu grass may reduce the chances of developing heart related diseases.
CONCLUSION
This study has shown that T. rendalli can effectively utilise kikuyu grass and duckweed. T.rendalli is pre-adapted to feeding on plants since it feeds on macrophytes in the wild. Kikuyu grass may be a good substitute for fishmeal in T. rendalli diets. Further studies are recommended to evaluate the extent to which kikuyu can replace fishmeal in T. rendalli practical diets. These studies must also focus on the identification and destruction of antinutritional factors in kikuyu.
ACKNOWLEDGMENTS
Authors are grateful to Dr. Joseph Sara and Mr. Johan. Theron for assistance with many aspects of this study. We extend our utmost appreciation to Mr. Gavin Geldenhuys for all the technical support. The National Research Foundation is acknowledged for funding this study.
REFERENCES
- Adewolu, M.A. and A.J. Adoti, 2010. Effect of mixed feeding schedules with varying dietary crude protein levels on the growth and feed utilization of Clarias gariepinus (Burchell, 1822) fingerlings. J. Fish. Aquat. Sci., 5: 304-310.
CrossRefDirect Link - Ali, A., S.M. Al-Ogaily, N.A. Al-Asgah, J.S. Goddard and S.I. Ahmed, 2008. Effect of feeding different protein to energy (P/E) ratios on the growth performance and body composition of Oreochromis niloticus fingerlings. J. Applied Ichthyol., 24: 31-37.
CrossRef - Azaza, M.S., F. Mensi, J. Ksouri, M.N. Dhraief, B. Brini, A. Abdelmouleh and M.M. Kraiem, 2008. Growth of Nile tilapia (Oreochromis niloticus L.) fed with diets containing graded levels of green algae ulva meal (Ulva rigida) reared in geothermal waters of southern Tunisia. J. Applied Ichthyol., 24: 202-207.
CrossRefDirect Link - Chikafumbwa, F.J.K., 1996. The use of napier grass (Pennisetum purpureum) and maize (Zea mays) bran as low-cost tilapia aquaculture inputs. Aquaculture, 146: 101-107.
CrossRef - Chowdhury, M.M.R., M. Shahjahan, M.S. Rahman and M.S. Islam, 2008. Duckweed (Lemna minor) as supplementary feed in monoculture of nile tilapia, Oreochromis niloticus. J. Fish. Aqua. Sci., 3: 54-59.
CrossRefDirect Link - Dioundick, O.B. and D.I. Stom, 1990. Effect of dietary a-cellulose levels on the juvenile tilapia Oreochromis mossambicus (Peters). Aquacult., 91: 311-315.
CrossRef - El-Sayed, A.F.M., 2006. Tilapia Culture. CAB International, Wallingford, England, ISBN: 9781845930165, Pages: 277.
Direct Link - Enami, H.R., 2011. A review of using canola/rapeseed meal in aquaculture feeding. J. Fish. Aquat. Sci., 6: 22-36.
CrossRefDirect Link - Lanna, E.A.T., L.E. Pezzato, P.R. Cecon, W.M. Furuya and M.A.D. Bomfim, 2004. Apparent digestibility and gastrointestinal transit in Nile tilapia (Oreochromis niloticus) in function of the dietary crude fiber. R. Bras. Zootec., 33: 2186-2192.
CrossRef - Nguyen, T.N., D.A. Davis and I.P. Saoud, 2009. Evaluation of alternative protein sources to replace fish meal in practical diets for juvenile tilapia, Oreochromis spp. J. World Aquacult. Soc., 40: 113-121.
CrossRefDirect Link - Okeyo, D.O. and W.L. Montgomery, 1992. Growth and conversion efficiency in the Blue Tilapia, Orechromis aureus, when fed aquatic macrophytes. J. Arizona-Nevada Acad. Sci., 24-25: 1-10.
Direct Link - Perschbacher, P.W., M.A. Lihono and J. Koo, 2010. GMT nile tilapia Orechromis niloticus growth and lipid composition fed a spirulina commercial pellet combination or commercial pellet only.GMT nile tilapia Orechromis niloticus growth and lipid composition fed a spirulina commercial pellet combination or commercial pellet only. Asian Fish Sci., 23: 91-99.
Direct Link - Riche, M., D.I. Haley, M. Oetker, S. Garbrecht and D.L. Garling, 2004. Effect of feeding frequency on gastric evacuation and the return of appetite in tilapia Oreochromis niloticus (L.). Aquaculture, 234: 657-673.
CrossRef - Saha, S., R.N. Roy, S.K. Sen and A.K. Ray, 2006. Characterization of cellulase-producing bacteria from the digestive tract of tilapia, Oreochromis mossambica (Peters) and grass carp, Ctenopharyngodon idella (Valenciennes). Aquacult. Res., 37: 380-388.
CrossRef - Setlikova, I. and Z. Adamek, 2004. Feeding selectivity and growth of the Nile tilapia (Oreochromis niloticus L.) fed on temperate-zone aquatic macrophytes. Czech J. Anim. Sci., 49: 271-278.
Direct Link - Targett, T.E. and N.M. Targett, 1990. Energetics of food selection by the herbivorous parrotfish Sparismosa radians : Roles of assimilation efficiency, gut evacuation rate, and secondary metabolites. Mar. Ecol. Prog. Ser., 66: 13-21.
Direct Link - Winberg, G.G., 1965. Rate of metabolism and food requirements of fishes. Fish. Res. Bd. Can. Trans. Ser., 194: 1-202.
Direct Link - Musuka, C.G., J.S. Likongwe, J. Kang'ombe, W.W.L. Jere and A.H. Mtethiwa, 2009. The effect of dietary protein and water temperatures on performance of T. rendalli juveniles reared in indoor tanks. Pak. J. Nutr., 8: 1526-1531.
CrossRefDirect Link - Uka, U.N. and K.S. Chukwuka, 2011. Utilization of aquatic macrophytes in Nigerian freshwater ecosystem. J. Fish. Aquat. Sci., 6: 490-498.
CrossRefDirect Link
Tshepo Josephat Sebake Reply
Awesome...thats all I can say! No team can beat that of Prof. Moyo and Sam, two inteligent hardworking scientists is what this A-Team is made up of. Supported by Dr. J. Sara, Mr. J. Theron and Mr. G. Geldenhuys! This Paper is gonna open doors on Tilapia farming looking at its quality.