
F. Liu
State Key Laboratory of Agrobiotechnology, China Agricultural University, Beijing, 100193, China
J. Fei
State Key Laboratory of Agrobiotechnology, China Agricultural University, Beijing, 100193, China
N. Li
State Key Laboratory of Agrobiotechnology, China Agricultural University, Beijing, 100193, China
Q. Meng
State Key Laboratory of Agrobiotechnology, China Agricultural University, Beijing, 100193, China
Asian Journal of Animal and Veterinary Advances
Year: 2010 | Volume: 5 | Issue: 8 | Page No.: 557-565







ABSTRACT
Muscle regeneration recapitulates myogenesis. HGF (hepatocyte growth factor) plays an important role in muscle development, as the only secreted growth factor, which can activates myoblasts proliferation. The expression units of wild HGF promoter-luciferase vector exceeded the expression units of HGF promoter-luciferase vector with mutated binding sequence of MyoD, also the expression trend of HGF was in accordance with the expression of MyoD. The expression of MyoD directly affects the expression of HGF during muscle development. In C2C12 culture, the expression of HGF is up-regulated in the phase of proliferation process, but down-related in the phase of differentiation. We identified the proliferation and differentiation stage of C2C12 culture by the definition of expression of myogenic regulator family (MyoD, yf5, yogenin, Myf6) and detected the HGF level during this two stages. MyoD bind site was found in the 5’-UTR of HGF promoter. MyoD is b HLH (basic helix-loop-helix) transcriptional activator which has a critical role during myogenesis. We verified that MyoD can bind the active b HLH site, as shown by EMSA. Transfection of wild HGF promoter-luciferase eukaryotic expression vector with MyoD binding site and HGF promoter-luciferase eukaryotic expression vector with mutated MyoD binding site into C2C12 myoblasts was done.
PDF Abstract XML References Citation
How to cite this article
F. Liu, J. Fei, N. Li and Q. Meng, 2010. The Muscle Transcription Factor MyoD Regulates the Expression of Hepatocyte Growth Factor during the Process of Muscle Development. Asian Journal of Animal and Veterinary Advances, 5: 557-565.
DOI: 10.3923/ajava.2010.557.565
URL: https://scialert.net/abstract/?doi=ajava.2010.557.565
DOI: 10.3923/ajava.2010.557.565
URL: https://scialert.net/abstract/?doi=ajava.2010.557.565
INTRODUCTION
Muscle requires adequate numbers of myoblasts (MB) for the formation of muscle fibers during muscle development, renewal and injured repair. MB are derived from pluripotent precursors that have the capacity to differentiate toward osteocyte, adipocyte, chondrocyte,or fibroblast phenotypes. The commitment of mesenchymal precursor cells along any one of these pathways is regulated by transcription factors that direct tissue-specific gene expression. Transcription factors organize target genes within the nuclear matrix to support a specific differentiation procedure (Cossu et al., 1996). Transcription factors perform their function as the form of dipolymer, which belong to bHLH family. bHLH transcription factors can activate the expression of muscle-correlating factor (Cossu et al.,1996). For MB, important transcription factors for phenotype commitment include Pax3, C-met (receptor of HGF) and HGF (hepatocyte growth factor) (Birchmeier and Gherardi, 1998). The HGF was originally know as a mitogen, which has been widely accepted that HGF plays an important role in organ and tissue development and regeneration (Grounds and Yablonka-Reuveni, 1993). The HGF can stimulate differentiation, propagation, regeneration and morphogenesis of numerous kinds of cells. The expression of HGF and its receptor c-met was detected in undifferentiated mouse C2 myoblasts and show up-regulated tendency. With withdraw of cell cycle into differentiation, the transcription level of HGF and its receptor is down-regulated. It was generally accepted that the transduction signal of combination of HGF and its receptor is a negative regulation to myoblasts’ differentiation (Anastasi et al., 1997). The HGF is a key regulator and the only known growth factor in muscle regeneration (Bladt et al., 1995). HGF’s pleiotropic role is most important during the early phase of muscle regeneration by the mean of mitogenic and chemotactic activities, resulting in an optimal myoblasts density to impair muscle fibers or form new muscle fibers (Buckingham, 2000). The MRF (include MyoD, Myf5, Myogenin, Myf6/Myf4), basic helix-loop-helix transcriptional activators of the myogenic regulator family, which play an indispensable role in muscle development (Tapscott and Weintraub, 1991). During the phase of proliferation and differentiation, the pattern of HGF expression was consistent with the pattern of MyoD expression and Myf5 expression. Additionally, myoblasts are widely termed proliferative MyoD and/or Myf5 positive myogenic cells (Grounds and Yablonka-Reuveni, 1993). Maybe some connections exist between HGF and MyoD/Myf5, which is poorly proved. Through HGF promoter sequence analysis we found that the proximal HGF promoter contains a binding sequence for MyoD. MyoD can bind to canonical E-box sequence CANNTG (Di Renzo et al., 1991), which was done by the experiment of EMSA. This report mainly elucidated that HGF can bind to MyoD and be regulated by MyoD. During proliferation phase and differentiation phase of myoblasts, the expression of HGF has similar pattern with MRF expression. The expression rate of HGF promoter-luciferase vector with binding sequence for MyoD exceeds the expression rate of HGF promoter-luciferase vector with mutated binding sequence for MyoD. Together, MyoD can not only bind to HGF, but also regulates the expression of HGF.
MATERIALS AND METHODS
Cell Culture
The C2C12 mouse pre-myoblast cells were purchased from china XieHe cell center. Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; neuronbc-biotech, Beijing) supplemented with 10% fetal bovine serum (FBS, Hyclone) and 1% antibiotics (penicillin, 100,000 units L-1 and streptomycin,100,000 units L-1; Gibco) and grown in a humidified atmosphere of 10% CO2 in air at 37°C. During differentiation, cells were cultured in DMEM with 2% FBS and 1% antibiotics (penicillin,100,000 units L-1 and streptomycin, 100,000 units L-1; Gibco) and grown in a humidified atmosphere of 10% CO2 in air at 37°C (Defrances et al., 1992). Cells were cultured with 104 mL-1 in 6-well plates. Cells were counted each day until they reached confluent. Later, cells were examined 3, 5, 7 days.
Real-Time PCR
Primers were made from mouse HGF genome, which are used for RT-PCR are shown in Table 1. Conditions for Real time PCR condition are 95°C, 10 min, then 95°C, 15 sec, 60°C, 1 min (40 cycles). Cells total RNA were extracted by All prep DNA/RNA mini kit (QIANGEN) 3, 5, 7 days. Total mRNA were reverse transcribed to cDNA, Cells RNA were examined 3, 5, 7 days.
| Table 1: | Primers used in real time PCR |
 | |
EMSA
Probes were made from the proximal 300 bp HGF promoter. The mutated probe was synthesized with a Not I restriction enzyme site. DNA oligonucleotides were annealed with annealing buffer (BIYUNTIAN).Nuclear protein were extracted from C2C12 cells cultured 3 days. C2C12 cells were cultured with the density of 104 mL-1. 32P-labeled double-stranded oligonucleotides (0.04 pmol) spanning 30/24 bp of the indicated sequence wad added to 10 μL EMSA buffer (100 mM Tris-HCl (pH 6.8), 10 mM EDTA, 500 mM Kcl, 100 mM β-mercaptoehanol, 10 mM DTT, 40% glycerol) and 2-4 μg poly (deoxyinosine-deoxycytosine). For preparation of each γ-[32P]-labeled probe, the oligonucleotides were annealed, radiolabeled with T4 polynucleotide kinase and γ-[32P]ATP and separated from unreacted nucleotides by purification with QIAquick Nucleotide removal Kit (QIAGEN). The reaction mix included nuclear protein (2 μg). After 30 min of incubation, the reaction mixture was loaded onto a standard 5% denatured polyacrylamide gel in 1xTris glycine/EDTA buffer. After electrophoresis, the gel was dried with DEAE paper and exposed to autoradiographic film for the appropriate length of time (24 h to overnight).
Cell Transfection
The HGF promoter fragment and mutated HGF promoter fragment, respectively was inserted into pGL3-Basic luciferase eukaryote expression vector. C2C12 cells were cultured in 6-well plates with 104 mL-1. Until cells reached 50-60% confluence, transfect promoter reporter constructs into cells, after 24 h transfection, we substitute original medium for fresh growth substrate. The cells were collected to detect the luciferase activities in 3, 5, 7 days.
Statistics
Each experiment was performed 2 or 3 times, with 4 or 5 replicates per treatment. Data of experiments were showed at the forms of Mean±SEM, t-test was used between two groups and single factor variance analysis among several groups, statistics difference was estimated by p<0.05.
RESULTS
Cells Growth Curve and the Identification of Proliferation and Differentiation (RT-PCR)
Cells were placed in 6-well plates with 103,104,105 mL-1. Finally, from Fig. 1 of cells growth curve (cell numbers and cells morphology) can show a best tendency change with the density of 104 mL-1. Figur 2a-f show C2C12 cell shapes of proliferation and differentiation phases on 3th, 5th, 7th day. MRF is the key family of regulating myogenesis, so we defined the proliferation and differentiation phase of myogenesis through the expression pattern of MRF in cultured myoblasts. We imitated myogenesis in the model of myoblasts culture in vitro. MRF (MyoD, Myogenin, Myf5, Myf6) expression and HGF expression were assayed 3, 5, 7 days. Figure 3a-d showed that the expression of HGF was up-regulated in proliferation condition and down-regulated in differentiation condition. Its receptor C-MET expressed constitutively at different conditions.
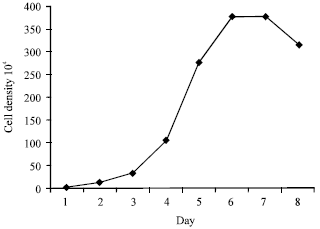 | |
| Fig. 1: | The growth curve of mouse C2C12 myoblasts cultured in growth medium. C2C12 myoblasts were cultured with the density of 104 mL-1 |
 | |
| Fig. 2: | Observation of cell C2C12 cultured in growth (a, b, c) and differentiation (d, e, f) medium, 10x/0.25 (a) C2C12 myoblasts grow fast with double polarized spread (b) Cell grew long and thin, running paralleling, fused each other and formed multinuclear pre-myotube and (c) more and more myotubes and the volume grew larger, (d, e, f) adjacent myotubes fused each other with muscle fibers trend |
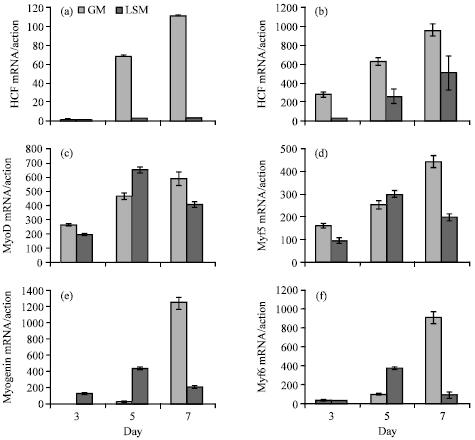 | |
| Fig. 3: | (a-f) Expression of HGF, MyoD, Myogenin, Myf5, Myf6 was measured by quantitative real-time PCR. mRNA extracted from C2C12 cells cultured under growth medium and differentiation medium. Results are Mean±SEM *p<0.05 |
From Fig. 3c, MyoD expression showed upward trend from 3 to 7 days under growth medium, but under differentiation medium MyoD expression went down from 7th day, This phenomenon was in accordance with MyoD expression in muscle development. MyoD is known to maintain myoblasts linage and keep myoblast cells proliferation. Myf5 expression pattern was consistent in MyoD expression pattern, also Myf5 played the same role in muscle development as MyoD (Fig. 3e). Myogenin expression show a high expression at 7th day, which shows myoblast cells began to differentiate, Myogenin originates differentiation in muscle development, which show the function in myoblast cells culture (Fig. 3d). Myf 6 regulates terminal differentiation and muscle fiber maintenance (Grounds and Yablonka-Reuveni, 1993). From Fig. 3f, Myf6 expression keeps a steady tendency in the final phase of differentiation. The HGF expression showed rising trend in proliferation phase and represented declining trend in differentiation phase, also this tendency was in accordance with MyoD expression and Myf5 expression.
EMSA To determine whether the proximal MyoD binding site bound MyoD, EMSA was done. Nuclear protein was incubated with two 32P-labeled probes spanning the active MyoD binding site and mutated MyoD binding site. Figure 4a shows the location of the probes. Bond of nuclear protein with probe1 was observed. From Fig. 4b, we can know the probe was MyoD binding sequence in the proximal 300 bp HGF promoter. These results were concordant with the mutational studies identifying the proximal 300 bp MyoD binding site of HGF promoter start site as conferring the majority of MyoD transcriptional activity.
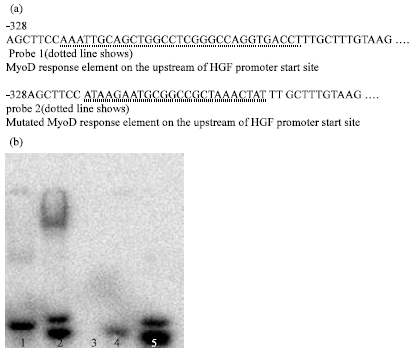 | |
| Fig. 4: | (a, b) Probe 1: MyoD binding fragment on the upstream of HGF promoter. Probe 2: mutated MyoD binding fragment. Lane 1: nuclear protein bind to probe 2 but not shown band; lane 2 nuclear protein bound to probe 1; lane 3, nuclear protein without any probe; lane 4, free probe 2; lane 5, free probe 1. EMSA showed probe binding to nuclear protein. The highest affinity binding was presented in probe 1, which spanned the region expected to confer MyoD activity |
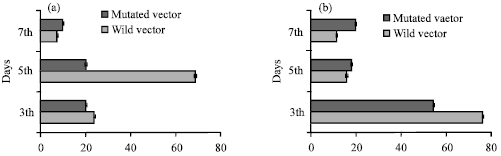 | |
| Fig. 5: | Luciferase activity assay. (a) Wild HGF promoter-luciferase expression vector transfected into C2C12 cells (cultured in growth medium), expression activity was assayed at 3th day and 5th day (b) Wild HGF promoter-luciferase expression vector transfected into C2C12 cells (cultured in differentiation medium), expression activity was assayed at 3th day and 5th day. Results show arbitrary luciferase units corrected for transfection efficiency. All transfection experiments were repeated four times p>0.05 |
Cell Transfection
To determine the proximal MyoD binding site of HGF promoter start site conferred the majority of MyoD transcriptional activity. Transfection of 3.1 kb HGF promoter (with MyoD binding site)-luciferase and 3.1 kb HGF promoter with the mutated binding site into C2C12 cells cultured with different conditions. The expression activity were assayed 3, 5 days. The result was concordant with the pattern of the regulation of MyoD to HGF expression. The expression units of wild construct exceeded the expression rate of mutated construct. MyoD can bind the HGF promoter to regulate its expression in muscle development (Fig. 5a, b).
DISCUSSION
The HGF is known as a mitogenic growth factor to stimulate hepatocyte proliferation and plays an important role in liver regeneration. Gradually, HGF is known to stimulate several kinds of cells to differentiate, propaganda, regenerate, mobile and morphogenesis as a multipotential growth factor, secreted by mesenchymal cells and eliciting its effects on epithelia. HGF can bind its receptor c-met to perform the correlative biological function (Hartmann et al., 1998). The research work of Anastasi showed that HGF and its receptor expressed in the undifferentiated mouse c2 muscle cells and perform their function by autocine to down-regulate differentiation of pre-myoblast (Anastasi et al., 1997). HGF is the only known growth factor that activates quiescent satellite cells in skeletal muscle. The process of satellite cell activation and differentiation during muscle regeneration is reminiscent of embryonic muscle development. The HGF has emerged as an important candidate factors in muscle regeneration for several reasons. 1) The c-met receptor is present on quiescent satellite cells in normal muscle tissue and precedes expression of other myogenic developmental genes. Addition of HGF to either cultured satellite cells or normal muscle tissue promotes entry of quiescent satellite cells into cell cycle. 2) HGF is expressed not only in regenerating muscle but also in normal muscle tissue and it can be released upon injury. 3) HGF promotes migration of muscle cells in vitro (Allen et al., 1995).
Muscle development involves many factors. At molecular level, activation of myogenic precursor cells is characterized by the rapid up-regulation of two MRFs (Weintraub, 1993), Myf5 and MyoD. MyoD and Myf5 are known to maintain pre-myoblast proliferation and keep high expression in proliferation phase of muscle development (Yablonka-Reuveni et al., 1999). Myogenin works to originate muscle differentiated development (Hasty et al., 1993). Myf4 maintain terminal differentiation and muscle fibers quantities. This report is the first to demonstrate the relationship of HGF expression and myogenic determination factor in muscle development. Muscle development procedure is a long-term and complex, so the study show the proliferation and differentiation stage of muscle development by the expression of MRF (MyoD, Myogenin, Myf5, Myf4) in cultured mouse C2C12 pre-myoblast. We use this model to conduct the expression pattern of HGF. The result shows that the expression pattern of HGF was concordant with the expression of MyoD and Myf5.
The mechanisms and factors regulating HGF expression in muscle development are poorly understood. The proximal sequence of HGF promoter start site contains MyoD binding site. MyoD can bind to this site by EMSA assay. The regions conferring stimulation by MyoD were identified by mutational analysis of binding site. We conduct transfection experiment to assay the regulating function of MyoD. The effect on HGF expression rate with transfection of a MyoD binding site HGF- promoter expression vector is higher than the expression rate with transfection of a mutated MyoD binding site HGF promoter expression vector. The expression pattern of a MyoD binding site HGF promoter plasmid was concordant with the expression of MyoD and Myf5.
The overview of HGF regulation in muscle development needs us to conduct the downstream pathway. The regulation of HGF transcription level made it perform its biological function through downstream signaling pathway (Comoglio, 2001). Present study shows the upstream regulation of HGF gene expression in muscle development procedure. In this research, we can know MyoD directly affects the expression of HGF during muscle development or muscle regeneration. Importantly, the expression trend of HGF is unanimous with the expression trend of MyoD. Maybe, the combination of MyoD and HGF could produce an effect on HGF biological function. The HGF can bind its receptor c-met to activate several signal pathways to perform morphogenesis and migration. They facilitate myoblasts migration and branching morphogenesis through the recruitment of downstream related factors, which play very important roles in limb skeletal muscle development. PI3K and Ras/MAPK are the prominent pathways in muscle development and some researches have provided some potent evidences (Zhang et al., 2003). JAK2/STAT2/STA3 effects the expression of HGF and MyoD. Apparently, STAT3 can stimulate the expression of the two genes simultaneously (no data shown). Presumably, STAT3 does so by cooperating with different related factors. The expression patterns of some factors involved in the pathway are not shown (Di Renzo et al., 1991; Wang et al., 2008). Together, these studies imply that HGF plays a prominent role in regulating the early phases of muscle development or muscle regeneration, correlating with expression of MyoD. Furthermore, additional interesting work will be required to conduct the important regulator of HGF gene in muscle development.
The study first systemically conduct the regulation effect of MyoD on the impact of HGF gene expression from the upstream molecular transcriptional level in muscle development, which links the MRF family with secreted growth factor HGF.
ACKNOWLEDGMENT
This study was supported by The National Key Scientific Program of China (No. 2009CB941600) and The National Natural Science Foundation of China (30800777).
REFERENCES
- Allen, R.E, S.M. Sheehan, R.G. Taylor, T.L. Kendall and G.M. Rice, 1995. Hepatocyte groth factor activates quiescent skeletal muscle satellite cells in vitro. J. Cell. Physiol., 165: 307-312.
PubMed - Anastasi, S., S. Giordano, O. Sthandier, G. Gambarotta, R. Maione, P. Comoglio and P. Amati, 1997. A natural hepatocyte growth factor/scatter factor autocrine loop in myoblast cells and the effect of the constitutive met kinase activation on myogenic differentiation. J. Cell Biol., 137: 1057-1068.
Direct Link - Birchmeier, C. and E. Gherardi, 1998. Development roles of HGF/SF and its receptor, the c-Met tyrosine kinase. Trends Cell Biol., 8: 404-410.
PubMed - Bladt, F., D. Rlethmacher, S. Isenmann, A. Aguzzi and C. Birchmeier, 1995. Essential role for the c-met receptor in the migration of myogenic precursor cells into the limb bud. Nature, 376: 768-771.
Direct Link - Comoglio, P.M., 2001. Pathway specificity for Met signaling. Nat. Cell Biol., 3: E161-E162.
Direct Link - Cossu, G., S. Tajbakhsh and M. Budkinghanm, 1996. How is myogenesis initiated in the embryo. Trends Genet., 12: 218-223.
PubMed - Di Renzo, M.F., R.P. Narsimhan, M. Olivero, S. Bretti and S. Giordano et al., 1991. Expression of the Met/HGF receptor in normal and neoplastic human tissues. Oncogene, 6: 1997-2003.
PubMed - Defrances, M.C., H.K. Wolf, G.K. Michalopoulos and R. Zarnegar, 1992. The presence of hepatocyte growth factor in the developing rat. Development, 116: 387-395.
Direct Link - Grounds, M.D. and Z. Yablonka-Reuveni, 1993. Molecular and cell biology of skeletal muscle regeneration. Mol. Cell Biol., 3: 210-256.
PubMed - Hartmann, G., T. Prospero, V. Brinkmann, C. Ozcelik and G. Winter et al., 1998. Engineered mutants of HGF/SF with reduced binding to heparan sulphate proteoglycans, decreased clearance and enhanced activity in vivo. Curr. Biol., 16: 125-134.
PubMed - Hasty, P., A. Bradley, J.H. Morris, D.G. Edmondson, J.M. Venuti, E.N. Olson and W.H. Klein, 1993. Muscle deficiency and neonatal death in mice with a targeted mutation in the Myogenin gene. Nature, 364: 501-506.
CrossRefDirect Link - Wang, K., C. Wang, F. Xiao, H. Wang and Z. Wu, 2008. JAK2/STAT2/STAT3 are required for myogenic differentiation. J. Biol. Chem., 283: 34029-34036.
PubMed - Tapscott, S.J and H. Weintraub, 1991. MyoD and the regulation of myogenesis by helix-loop-helix proteins. J. Clin. Invest., 87: 1133-1138.
PubMed - Weintraub, H., 1993. The MyoD family and myogenesis: Redundancy, networks and thresholds. Cell, 75: 1241-1244.
PubMed - Yablonka-Reuveni, Z., M.A. Rudnicki, A.J. Rivera, M. Primig, J.E. Anderson and P. Natanson, 1999. The transition from proliferation to differentiation is delayed in satellite cells from mice lacking MyoD. Dev. Biol., 210: 440-455.
PubMed - Zhang, Y.W. and F.V.W. George, 2003. HGF/SF-met signaling in the control of branching morphogenesis and invasion. J. Cell. Biochem., 88: 408-417.
Direct Link