
A. Dehghan
Department of Clinical Sciences, School of Veterinary Medicine, Islamic Azad University-Kazerun Branch, Kazerun, Iran
LiveDNA: 98.33091
ORCID: 0000-0002-4090-0711
M. Arabi
Department of Clinical Sciences, School of Veterinary Medicine, Islamic Azad University-Kazerun Branch, Kazerun, Iran
LiveDNA: 98.5116
S. Nahid
Department of Basic Sciences, Islamic Azad University-Darab Branch, Darab, Iran
LiveDNA: 98.14032
M. Aminlari
Department of Biochemistry, School of Veterinary Medicine, University of Shiraz, Shiraz, Iran
LiveDNA: 98.12114
Asian Journal of Animal and Veterinary Advances
Year: 2010 | Volume: 5 | Issue: 7 | Page No.: 472-477







ABSTRACT
Oxidative stress commonly occurs following heat stress in tropical regions and affects dairy animals. Glutathione protects cells from oxidative damages. This study was carried out to investigate the serum glutathione level in the ram with a fluorometric method and to determine its changes during heat stress condition. Eight mature rams were selected and kept in the same conditions. The rams were maintained during temperate and warm seasons to compare serum glutathione levels during normal and heat stress conditions, respectively. Heat stress was considered when temperature-humidity index was above 84. Serum samples were obtained at 0, 14 and 28 days after beginning of the study during the seasons. Reduced and oxidized glutathione concentrations were determined using a fluorometric assay. The serum concentrations of reduced glutathione in the normal and heat stress conditions were lower than oxidized glutathione. The reduced and oxidized glutathione levels and their ratios were not different between seasons and at different sampling times, although they were significantly changed during sampling times in the normal and heat stress conditions. Present results represent that glutathione levels change during different environmental conditions. It seems that antioxidant defense system was changed to adapt and prevent oxidative stress effects, although needs further researches.
PDF Abstract XML References Citation
How to cite this article
A. Dehghan, M. Arabi, S. Nahid and M. Aminlari, 2010. Changes of Serum Reduced and Oxidized Glutathione in Heat Stressed Ram. Asian Journal of Animal and Veterinary Advances, 5: 472-477.
DOI: 10.3923/ajava.2010.472.477
URL: https://scialert.net/abstract/?doi=ajava.2010.472.477
DOI: 10.3923/ajava.2010.472.477
URL: https://scialert.net/abstract/?doi=ajava.2010.472.477
INTRODUCTION
Heat stress can limit dairy production in hot climates (Finocchiaro et al., 2005). High ambient temperature and humidity are the major constraint on sheep productivity in tropical and subtropical areas (Marai et al., 2007). The spermatogenesis and testicular function decrease during summer in sheep which is related to photoperiod (Rhim et al., 1993; Rasooli et al., 2010). High ambient temperature during summer months may increase the suppressive effect of photoperiod on testicular function in ram (Rasooli et al., 2010).
The oxidative balance is affected during heat stress periods. Fast production of free radicals and reactive oxygen species and/or a decrease in antioxidant defense mechanisms result in oxidative stress (Sies, 1997; Sies et al., 2005; Trevisan et al., 2001; Bernabucci et al., 2002).
Glutathione (γ-glutamyl-cysteinyl-glycine) is the principal tripeptide involved in the antioxidant cellular defense mechanisms. Glutathione is produced in all mammalian organs, especially in the liver (Pastore et al., 2003). Free glutathione is mainly present in its reduced form (GSH), which can be converted to the oxidized form (GSSG) during oxidative stress. Glutathione reductase can convert GSSG to GSH.
Reduction and conjugation reactions are the most important functions of GSH (Pastore et al., 2003; Forman et al., 2009). The GSH protects cell membranes from oxidative damage by removing of produced reactive species (Pastore et al., 2003; Forman et al., 2009).
Plasma glutathione levels are normally in the micro-molar range. Liver supplies most of the plasma glutathione. It is removed from plasma by transpeptidase action which is mostly located in the kidney (Pastore et al., 2003). Liver and kidney are metabolically active tissues; therefore, glutathione levels may be influenced by different physiological and pathological conditions.
There is little information about serum indicators of oxidative stress and antioxidant status of the rams in normal and heat stress conditions. The aims of the present study were to determine serum GSH and GSSG concentrations in rams by a fluorometric method and to evaluate their changes during heat stress conditions.
MATERIALS AND METHODS
The experiment was conducted during February to July 2007 in Nurabad (30°7' N latitude, 51°31' E longitude), Fars province, Iran.
Materials
All chemicals were purchased from Sigma Chemical Co. unless otherwise specified.
Animals and Managements
Eight mature rams (3 to 4-year-old) with the same weight were selected. The rams were fed on pasture regularly with allowed free access to water.
The study was conducted during temperate and warm seasons of the year to compare parameters in normal and heat stress conditions. The rams were separated from females during the study to prevent the effects of reproductive activities.
Estimation of Heat Stress Severity
Temperature-Humidity Index (THI) was used to assess the levels of heat stress condition and calculated by the following formula (Marai et al., 2007):
where, db °F is the dry bulb temperature in °F and RH is the relative humidity (RH%)/100.
Environmental temperature and humidity were measured 3 times daily with 8 h interval at 6:00, 14:00 and 22:00. The THI values obtained above 84 were considered heat stress condition (Finocchiaro et al., 2005; Marai et al., 2007).
Blood Sampling
Jugular blood samples were collected days 0, 14 and 28 after beginning of the study in each season. During summer, sampling was initiated while THI value was obtained above 84. The blood samples were kept ice cold until serum was harvested. Serum samples were kept at -80°C until measurements.
Measurement of GSH and GSSG Concentrations
Concentration of oxidized (GSSG) and reduced (GSH) forms of glutathione were determined using a fluorometric method described by Hissin and Hilf (1976). Briefly, serum samples were deproteinized by adding equal volume of 5% trichloroacetic acid and centrifugation at 10000x g for 10 min. The GSH and GSSG concentrations were measured in μmol L-1.
For GSH measurement, 200 μL of GSH standard solution or supernatant was added to 2.6 mL of phosphate-EDTA buffer (0.1 M sodium phosphate, 5 mM EDTA, pH = 8) and 200 μL of 1 mg mL-1 O-phthaldialdehyde (OPA). The mixture was remained at room temperature for 45 min and then fluorescence detection was done with excitation and emission at 350 and 418 nm, respectively by Perkin-Elmer LS50 fluorescence spectrophotometer (USA) (Hissin and Hilf, 1976).
For GSSG measurement, 200 μL of supernatant was added to 100 μL of 40 mM N-ethylmaleimide (NEM) and remained 30 min at room temperature and then 2.5 mL of NaOH (0.1 N) was added. The NEM react with GSH and remained GSSG reduced to GSH at these steps. Two hundred microliter of this solution was used to react with OPA by described procedure.
Statistical Analysis
The Statistical analysis were done using SPSS software, version 16. Results are presented as Mean±SD. Statistical analysis carried out by the GLM Univariate procedure in the different seasons and sampling times. The seasons were used as fixed effect and the times of sampling were used as random effect. The Paired-Samples t-test procedure was used to compare the variables in the same seasons (Petrie and Watson, 2006). The p<0.05 was considered as statistically significant.
RESULTS
The averages of THI value during temperate and warm season were 64.4±2.7 and 86.4±1.2, respectively.
The concentrations of GSH and GSSG in normal and heat stress conditions were shown in the Table 1. From 48 blood samples collected, the serum concentrations of reduced and oxidized glutathione were not different in normal conditions, although, oxidized glutathione were lower than reduced glutathione in heat stress condition (p<0.001).
The comparison of serum values between different sampling days during Summer and Winter shows that GSH value in 0 sampling day of Summer is higher than 0, 14 and 28 days of Winter (p<0.05). Also, this value in 14 sampling day is higher than 14 and 28 days of winter (p<0.05); while, GSH value in 28 sampling day of summer is significantly lower than 0 days of Winter (p<0.05).
Serum GSSG values in 0 sampling day of summer is significantly higher than 14 day of Winter; while, this value in 28 day of Winter is higher than 0, 14, 28 of Summer (p<0.05).
The GSH to GSSG ratios in 0 and 14 sampling days of Summer are significantly higher than all sampling days of Winter (p<0.05); although, this value in 14 day of Winter is higher than 28 day of Summer (p<0.05).
The data showed that the GSH and GSSG levels and also GSH to GSSG ratios were not different between seasons and at different sampling times (p>0.05), although, they significantly changed during different sampling times in the normal and heat stress conditions (Table 1).
| Table 1: | The effects of heat stress on oxidative status of the rams |
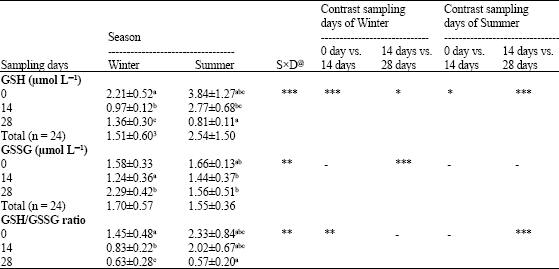 | |
| Normal condition is mentioned by winter samples. Means of each factor within winter column are different to rows in Summer column with same superscript (p<0.05). *p<0.05; **p<0.01; ***p<0.001. @SxD: SeasonxDays interaction | |
DISCUSSION
Measuring of blood glutathione (GSH and GSSG) is a useful way for detecting glutathione status in whole-body (Pastore et al., 2003). It was found that the GSH and GSH/GSSG ratio is low in the blood of patients with various diseases (Lyons et al., 2000; Pastore et al., 2003).
The cellular level of GSH is predominating over the GSSG in normal condition (Pastore et al., 2003). The tissue and plasma levels of glutathione change during different situations. The plasma level of glutathione decreases when its synthesis reduces in tissues (Griffith and Meister, 1979; Pastore et al., 2003). Also, glutathione can be metabolized by tissues, such as kidney and mammary tissue (Pocius et al., 1981). In addition, nutritional factors strongly influence hepatic and plasma GSH content (Chao and Allen, 1992; Hum et al., 1992; Kirn et al., 1992).
During sampling times of the present study, the serum GSH levels and GSH/GSSG ratios decreased in normal and heat stress conditions (p<0.05). Although, GSSG levels increased in normal condition (p<0.05) and did not change in heat stress condition. Sgorlon et al. (2008) also found that total glutathione concentrations significantly decreased with the time of sampling during normal condition.
The presented studies (Griffith and Meister, 1979; Pocius et al., 1981; Chao and Allen, 1992; Hum et al., 1992; Kirn et al., 1992; Pastore et al., 2003; Sgorlon et al., 2008) and present results suggested that glutathione levels may change during different physiological, pathological, nutritional and stress conditions in blood and tissues. It may be predictable that body needs to change antioxidant defense system regularly to be protected from oxidative stress.
Heat stress and oxidative stress are 2 types of damaging agents which have been reported to induce an adaptive response (Crawford and Davies, 1994). The main physiologic benefit of adaptive response is protection of cells from high doses of a toxic agent (Crawford and Davies, 1994; Leeuwenburgh et al., 1997).
The present data showed that the GSH and GSSG levels and their ratios were not different seasonally (p>0.05). No specific study in the literature identifies which has separately measured blood GSH and GSSG levels during heat stress condition in sheep or any other animals. Thiobarbituric (TBARS) assay, as an oxidative stress index, was not performed in the present study. Therefore, it is not obvious that oxidative stress has affected the presented rams. It is possible that the differences during sampling times were relatable to nutritional and metabolic factors or adaptation to environmental changes.
CONCLUSION
The sheep which rears in tropical regions adapts itself to encountered heat stress and glutathione levels change to prevent disturbing in antioxidant defense system and to defend against oxidative stress. It is suggested that the effects of heat stress on serum glutathione levels of sheep will be determined along with TBARS assay.
ACKNOWLEDGMENT
The authors wish to thank Dr. Hossein Tavallali from Shiraz Payame Noor University who had great helps in fluorometric assays.
REFERENCES
- Bernabucci, U., B. Ronchi, N. Lacetera and A. Nardone, 2002. Markers of oxidative status in plasma and erythrocytes of transition dairy cows during hot season. J. Dairy Sci., 85: 2173-2179.
CrossRefDirect Link - Chao, P.Y. and K.G.D. Allen, 1992. Glutathione production in copper deficient isolated rat hepatocytes. Free Radic. Biol. Med., 12: 145-150.
CrossRef - Crawford, D.R. and K.J. Davies, 1994. Adaptive response and oxidative stress. Environ. Health Perspect., 102: 25-28.
Direct Link - Finocchiaro, R., J.B.C.H.M. van Kaam, B. Portolano and I. Misztal, 2005. Effect of heat stress on production of mediterranean dairy sheep. J. Dairy Sci., 88: 1855-1864.
CrossRefDirect Link - Forman, H.J., H. Zhang and A. Rinna, 2009. Glutathione: Overview of its protective roles, measurement and biosynthesis. Mol. Aspects Med., 30: 1-12.
CrossRefPubMedDirect Link - Griffith, O.W. and A. Meister, 1979. Potent and specific inhibition of glutathione synthesis by buthionine sulfoximine (s-n butyl homocysteine sulfoximine). J. Biol. Chem., 254: 7558-7560.
PubMedDirect Link - Hissin, P.J. and R. Hilf, 1976. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem., 74: 214-226.
CrossRefPubMedDirect Link - Hum, S., K.G. Koski and L.J. Hoffer, 1992. Varied protein intake alters glutathione metabolism in rats. J. Nutr., 122: 2010-2018.
Direct Link - Leeuwenburgh, C., J. Hollander, S. Leichtweis, M. Griffiths, M. Gore and L.L. Ji, 1997. Adaptations of glutathione antioxidant system to endurance training are tissue and muscle fiber specific. Am. J. Physiol. Regul. Integr. Comp. Physiol., 272: R363-R369.
Direct Link - Lyons, J., A. Rauh-Pfeiffer, Y.M. Yu, X.M. Lu and D. Zurakowski et al., 2000. Blood glutathione synthesis rates in healthy adults receiving a sulfur amino acid-free diet. Proc. Natl. Acad. Sci. USA., 97: 5071-5076.
PubMed - Marai, I.F.M., A.A. El-Darawany, A. Fadiel and M.A.M. Abdel-Hafez, 2007. Physiological traits as affected by heat stress in sheep-A review. Small Rumin. Res., 7: 1-12.
CrossRefDirect Link - Pastore, A., G. Federici, E. Bertini and F. Piemonte, 2003. Analysis of glutathione: Implication in redox and detoxification. Clin. Cham. Acta, 33: 19-39.
CrossRefPubMedDirect Link - Pocius, P.A., J.H. Clark and C.R. Baumrucker, 1981. Glutathione in bovine blood: Possible source of amino acids for milk protein synthesis. J. Dairy Sci., 64: 1551-1554.
Direct Link - Rasooli, A., M.T. Jalali, M. Nouria, B. Mohammadian and F. Barati, 2010. Effects of chronic heat stress on testicular structures, serum testosterone and cortisol concentrations in developing lambs. Anim. Reprod. Sci., 117: 55-59.
CrossRefDirect Link - Rhim, T.J., D. Kuehl and G.L. Jackson, 1993. Seasonal changes in the relationships between secretion of gonadotropin-releasing hormone, luteinizing hormone and testosterone in the ram. Biol. Reprod., 48: 197-204.
Direct Link - Sgorlon, S., G. Stradaioli, G. Gabai and B. Stefanon, 2008. Variation of starch and fat in the diet affects metabolic status and oxidative stress in ewes. Small Rum. Res., 74: 123-129.
Direct Link - Sies, H., W. Stahl and A. Sevanian, 2005. Nutritional, dietary and postprandial oxidative stress. J. Nutr., 135: 969-972.
PubMedDirect Link - Sies, H., 1997. Oxidative stress: Oxidants and antioxidants. Exp. Physiol., 82: 291-295.
CrossRefPubMedDirect Link - Trevisan, M., R. Browne, M. Ram, P. Muti, J. Freudenheim, A.M. Carosella and D. Armstrong, 2001. Correlates of markers of oxidative status in the general population. Am. J. Epidemiol., 154: 348-356.
CrossRefDirect Link