
F. Rahim
Physiology Research Center, Ahwaz Jondishapour University of Medical Sciences, Ahwaz, Iran
B. Keikhaei
Research Center of Thalassemia and Hemoglobinopathies, Shafa Hospital, Ahwaz Jondishapour University of Medical Sciences, Ahwaz, Iran
A. Sarkaki
Physiology Research Center, Ahwaz Jondishapour University of Medical Sciences, Ahwaz, Iran
A.H. Doulah
Department of Nursing, Ahwaz Azad University, Ahwaz, Iran
Asian Journal of Animal and Veterinary Advances
Year: 2010 | Volume: 5 | Issue: 1 | Page No.: 13-23







ABSTRACT
The present study was conducted in order to investigate the effects of right-unilateral lesion of substantia nigra neurons by means of Ibotenic acid, a cholinergic-selective neurotoxin, on hematological parameters in rats. Thirty male Wistar rats weighing 200±50 g at the start of the experiment were used. The substantia nigra was right-unilateral lesioned by stereotaxic microinjections of ibotonic acid. Seven days after neurosurgery, we assessed the total number of White Blood Cells (WBC), the total number of Red Blood Cells (RBC), Red Cell Distribution (RDW), platelet and hemoglobin level and the erythrocyte indexes (Mean Cell Volume, (MCV), Mean Cell Hemoglobin,(MCH), Mean Cell Hemoglobin Concentration (MCHC)). Ibotenic acid treatment induced a highly significantly decrease of white blood cells, followed by significant decrease in red blood cells and hemoglobin level comparative with sham-operated rats. Also in the ibotenic acid-lesioned rats the erythrocyte indexes (Mean Cell Volume, MCV; Mean Cell Hemoglobin, MCH were significantly decreased comparative with sham-operated rats. By contrast, platelets, mean cell hemoglobin concentration and red blood cell distribution width were significantly increased in the ibotonic-acid lesioned rat versus sham-operated animals. On the whole, the obtained data indicate the important role of the central nerves system in the regulation of erythrocyte dynamics.
PDF Abstract XML References Citation
How to cite this article
F. Rahim, B. Keikhaei, A. Sarkaki and A.H. Doulah, 2010. Ibotenic Acid-Induced Hematological Disorders in Rat Model of Alzheimer's Disease. Asian Journal of Animal and Veterinary Advances, 5: 13-23.
DOI: 10.3923/ajava.2010.13.23
URL: https://scialert.net/abstract/?doi=ajava.2010.13.23
DOI: 10.3923/ajava.2010.13.23
URL: https://scialert.net/abstract/?doi=ajava.2010.13.23
INTRODUCTION
Alzheimer's Disease (AD) is characterized by degenerative changes in the brain (Etienne et al., 1986) and the impairment of learning and memory function (Miranda et al., 2003). It is widely accepted that learning and memory are deeply associated with the functional activity of the cholinergic system in the Central Nervous System (CNS). The basal Fore Brain (BF) provides the major source of cholinergic in put to the neocortex and hippocampus (Miranda et al., 2003) and the cholinergic neurons in the Nucleus Basalis of Mynert (NBM) are markedly degenerated in AD (Etienne et al., 1986). Because the Nucleus Basalis Magnocelluralis (NBM) in the rat is analogous to the NBM in humans, the NBM-lesioned rat has been regarded as an animal model for cholinergic dysfunction in the cerebral cortex of patients with AD (Wu et al., 2005). The NBM-lesioned rat shows decreases in cholinergic markers, such as acetylcholine (ACh) release (Meyer et al., 1987) and choline acetyltransferase (ChAT) activity (Arendash et al., 1987), in the cerebral cortex and shows learning and memory impairment (Salamone et al., 1984). In AD the principal neurochemical abnormality is the alteration of the cholinergic system in the Central Nervous System (CNS) (Zatta et al., 2002; Kaizer et al., 2005).
Hematopoiesis, the dynamic process of blood cell production and development, is characterized by a continuous, robust turnover of cells. The hematopoietic system, which consists of the Bone Marrow (BM), liver, spleen, lymph nodes and thymus, provides leukocytes, erythrocytes and platelets through a complex network of tissues, organs, stem cells and regulatory factors (Huang and Liu, 2009).
However, the mechanism causing anemia associated with autonomic dysfunction is not well explained. Catecholamines and their corresponding receptors are widely distributed in both the central and peripheral nervous system. Besides their vasoactive effect (Siarakas et al., 1997), catecholamines have been known to be involved in different forms of learning and memory (Hefco et al., 2003; Spreng et al., 2001). Immune cell types associated with innate immunity such as natural killer cells, neutrophils and macrophages are the potential subjects to be regulated by catecholamines because these cells express functional β2- and/or α-adrenergic receptors (Dent et al., 2001).
The processes of hematopoiesis in the bone marrow are regulated by a variety of factors including those produced by the stromal elements, the developing hematopoietic cells themselves and by exogenous substances, such as hormones and neurotransmitters, like acetylcholine. In particular, cutting nerves which enter the hip bones influenced erythropoiesis, while injecting nicotine inside the bone induced changes in the processes controlled by the autonomic nervous system (Desforges, 1984). These data demonstrated the role of the bone marrow innervation in hematopoiesis and the presence of nicotinic acetylcholine receptors within the bone marrow. This view was further supported by the recent studies of nicotine-stimulated changes in hematopoiesis (Khaldoyanidi et al., 2001; Serobyan et al., 2005). Nicotinic acetylcholine receptors (nicotinic receptors) are ligand-gated ion channels mediating synaptic transmission in nerve and muscle cells. They are composed of several types of alpha and beta subunits forming either homomeric or heteromeric functionally distinct receptor subtypes (Paterson and Nordberg, 2000). Nicotinic receptors are also present in many non-excitable cells, such as skin keratinocytes (Arredondo et al., 2003), respiratory tract epithelial cells, vascular endothelium (Conti-Fine et al., 2000) and most of the blood cells: leukocytes (Cormier et al., 2004), lymphocytes (Kawashima and Fujii, 2003), macrophages (Wang et al., 2003) and erythrocytes (Bennekou, 1993), where their functions are quite different from those in muscles or neurons. In summary, the primary goal of this study was to evaluate whether disorders hematopoiesis regulation via Nucleus Basalis Magnocelluralis (NBM) neuron lesion may induce hematological disorders.
MATERIALS AND METHODS
This research was conducted from June 2008 to June 2009.
Animals and Surgery
Thirty male inbreed Wistar rats, weighing 250-300 g and housed with free access to food and water. They were maintained in a 12 h light/dark cycle with lights on at 6:00 am at constant temperature (25°C). The animals were divided into three equal number groups (n = 15): Control, sham operated (ShO) and ibotenic acid (IBO). The rats were not found to have other causes of anemia, the nutritional state was adequate and no evidence of any hemorrhagic and thrombotic disorder.
Neurosurgery and Drug Administration
On the day of surgery, the animals were anesthetized with ketamine/xylazine (50 mg kg-1, i.p.) and placed in a stereotaxic apparatus (Kopf). The incisor bar was set -0.16 mm posterior and ±0.40 mm lateral to the bregma and -0.55 mm below the top of the skull (Ahmed et al., 2004; Paxinos and Watson, 2007; Yamamoto et al., 2003) to reach the nucleus basalis magnocellularis. Rats received unilateral infusions of 0.5 μL volume of vehicle (NaCl 0.9%) or ibotenic acid (8 μg μL-1) using a 5 μL Hamilton syringe. The injection was given over a period of 5 min and the needle was left in the injection site for a further 5 min. After behavioral tasks, rats were sacrificed for neurochemical analysis. The protocol concerning this research is in accordance with the guidelines of the committee on the care and use of Experimental Animal Resources, School of Medicine, Ahwaz Jondishapour University of Medical Sciences, Ahwaz, Iran. Hematological parameters were assayed 1 week after the neurosurgery.
Morris Water Maze Task
Training in the maze taken place during the light phase of the cycle between 8:00 am and 5:00 pm. A circular pool was used as described by Morris with some modification (Widy-Tyszekoeiwecz et al., 2002). It was a black circular pool (120 cm in diameter and 80 cm in height) filled with tap water (27±2°C) with a depth 60 cm. The maze was divided geographically into four equal size quadrants and release points were designed in each quadrant as North (N), East (E), South (S) and West (W). A hidden circular escape platform (12 cm in diameter), was emerged 2 cm below the water level and was located in the center of the northeast quadrant. Some fixed visual cues including computer, desk, shelves, posters and illumination lights were placed on the walls around the pool. A camera was positioned above the center of the pool that was connected to a computer to record the animal motions. An automated tracking system (Radiab ver. 2, Tehran, Iran) was used to measure the escape latency, swimming distance and speed.
Training Procedure
Twenty four hours after the last infusion of GH (8th day) animals were subjected to the training procedure of one session of four trials (block) daily for four consecutive days in the water maze. In each trial, the animals were allowed 60 sec to find the platform, after then were allowed to remain there for 30 sec, if did not find the platform within 60 sec animals were gently guided to the platform. After the completion of a trial, animals were returned to a holding cage for an intertrial interval of 60 sec. After 24 h of the last trial, platform was removed and rats were released from Southwest as a probe trial (consisted of a 60 sec free swim period) and the time spent in the target quadrant was recorded (Alaeia et al., 2008). The behavioral test include calculation of path length, escape latency, swimming speed and probe trial.
Blood Sampling Protocol
One week after neurosurgery, blood samples were withdrawn from lateral tail vein via the Biotrol sampling catheter from 15 sham operated and 15 ibotenic acid-treated rats. Blood samples (0.5 mL approximately/sample) were collected in vials containing EDTA for hematological investigations. Hematological parameters were assayed by a COULTER® Ac•T 5diff™ CP-precision instrument for hematology research. In order to rule out the iron deficiency anemia, the standard discrimination indices were calculated by using red blood cell indices as in our previous study (Keikhaei et al., 2007).
Histological ShO
At the end of the experiment, all rats were sacrificed with an overdose of sodium pentobarbital (100 mg kg-1 i.p.) followed by a transcardial infusion of 0.9% saline and a 10% formalin solution. The brains were removed and placed in a 30% sucrose/formalin solution. The brains were frozen and cut into coronal sections (50 μm) using a freezing microtome and stained with crystal violet for verification of the point of the syringe needle. Only experimental data from lesions correctly located in the substantia nigra were used for statistical analysis.
Data Presentations and Statistical Analysis
Statistical analysis was performed with SPSS software (v. 12.0; SPSS Inc., Chicago, IL, USA). Results were expressed as Mean±SEM. Because the data were not normally distributed, the non-parametric statistic Mann-Whitney U-test was employed. Results were considered significant if p<0.05. The number of observation was 30.
RESULTS AND DISCUSSION
Experimental data were registered 1 week after the ibotenic acid administration (Table 1). Comparing the swimming path length between the various groups did not indicate any differences during 4 days spatial training in water maze (p>0.05). Path length in each session in ibotenic acid-lesioned group was longer than control group significantly (p<0.01). Comparing the escape latency between the various non-lesioned groups did not indicate any differences during four days spatial training in water maze (p>0.05). Time required to find and locate on escape platform in each session in ibotenic acid-lesioned group was more than control group significantly (p<0.01). Total escape latency was increased in ibotenic acid-lesioned group after 4 days training significantly comparing to control (p<0.001). Comparing the swimming speed between the various groups did not indicate any differences during four days spatial training in water maze (p>0.05). The percent of time spent in goal quarter while escape platform removed during probe trial 24 h after 4 days training, decreased significantly in ibotenic acid-lesioned group (p<0.001) when comparing to control group.
| Table 1: | Demographic variables of study groups with their U and p-values |
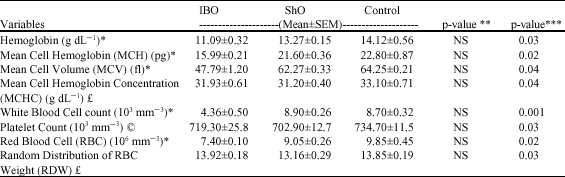 | |
| ©: Significant increase in values of variable among the ibotenic acid exposed group (IBO). *Significant decrease in values of variable among the sham operated group (ShO). £: Non significant difference. **p-values between control and ShO groups, ***p-values between control and IBO groups, NS: Not significant | |
| Table 2: | Differential value for each discrimination index in differentiation between non iron deficiency and iron deficiency anemia |
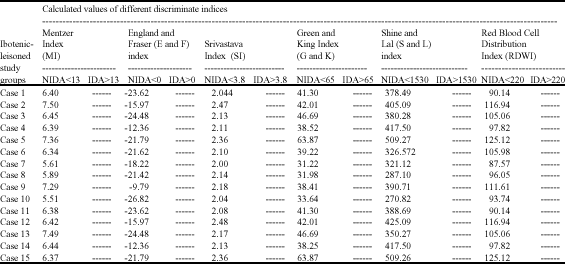 | |
| IDA: Iron deficiency anemia, NIDA: Non Iron deficiency anemia | |
We have calculated the different indices for ibotenic acid-lesioned rats individually. The differential value for each discrimination index in differentiation between non iron deficiency and iron deficiency anemia are shown in Table 2. In the ibotenic acid-lesioned rats, we observed a significant decrease in the total number of white blood cells (p<0.001), Mean Cell Volume (MCV) (p<0.05), red blood cells (p<0.05) and hemoglobin level (p<0.05) (Fig. 1A-E), compared with the sham-operated groups. Ibotenic acid significantly increased Red Cell Distribution Width (RDW) (p<0.05), platelet (p<0.05) and Mean Cell Hemoglobin Concentration (MCHC) (p<0.05) compared with sham-operated groups (Fig. 2A-C). In case of this three increased values, the high level was observed comparing to sham-operated group, but the values was also decreased comparing to control group (Fig. 2).
Beard et al. (1997) suggested that anemia increases the risk of Alzheimer's disease, although the mechanism remains to be elucidated. In addition, anemia can lead to other neurologic symptoms, such as headaches, loss of concentration and depression (Katz et al., 1993). It is well recognized that the immune response is under the influence of a variety of neural or neuroendocrine mechanisms. Much less studied is the possible influence of these mechanisms on hematopoiesis. Earlier studies reported that the central dopaminergic system has a crucial role in regulation of the immune processes as well as hematopoiesis (Nanda et al., 2005; Pacheco-Lopez et al., 2003). In the present study, we used a procedure of chemical sympathectomy by lesioning the basalis magnocellularis with ibotenic acid. By means of this particularly electrolytic lesion, we observed a significant decrease in hematological parameters registered 1 week after ibotenic acid administration, tested by the total number of leukocytes, erythrocytes, hemoglobin level and the erythrocyte indexes (MCV and MCH). Interestingly, we observed increase in platelet numbers, Red Dell Width (RDW) distribution and other erythrocyte index (MCHC) in ibotenic acid-lesioned rats comparing to sham operated group. We demonstrated that rats treated with ibotenic acid showed anemia. In addition, since the WBC significantly decreased during ibotenic acid-induced anemia, the effect of ibotenic acid in this experiment may be specific for erythropoiesis, as well as for bone morrow suppression.
In this study, we also demonstrated the low hemoglobin associated with AD which was in agreement with study done by Pandav et al. (2004). McCaddon et al. (2004) showed thathemoglobin and platelet counts fell only slightly with increasing dementia duration, but there were no other changes in hematological indices. In particular, macrocytosis and red cell distribution width were unrelated to disease duration and no patients were anemic. Present result not only showed the significant decrease in hemoglobin and other hematological indices also increase in red cell distribution width.
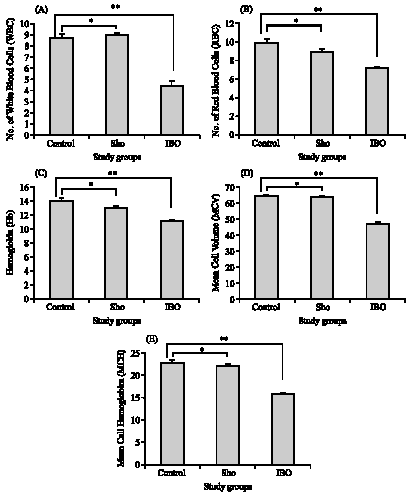 | |
| Fig. 1: | (A) The effect of the chemical lesion with ibotenic acid on total number of leukocytes (**p<0.03 vs. control group), (B) total number of erythrocytes (**p<0.01 vs. control group), (C) hemoglobin level (**p<0.02 vs. control group), (D) mean cell volume (**p<0.04 vs. control group) and (E) mean cell volume (**p<0.04 vs. control group) all tested one week after lesion; *all p-values of control vs. sham-operated group (p>0.05). Values are Mean±SEM (n = 15 per group) |
The regulation of the hematopoietic system is achieved at three levels: (1) at the cellular level of bone marrow stroma, (2) at the humoral level by cytokines and (3) by catecholamines and other neuroendocrine factors. Sympathetic nerve endings and bone marrow cells are the main source of bone marrow catecholamines (Felten, 1993; Maestroni, 1998; Hoogduijn et al., 2006).
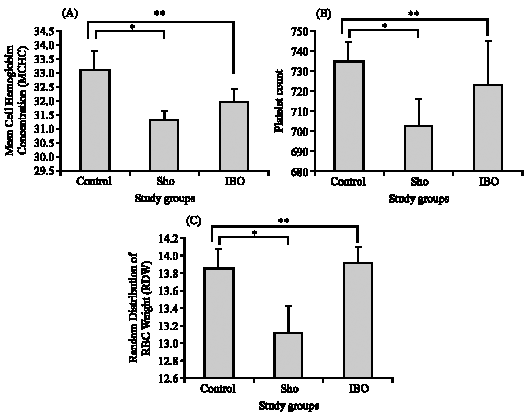 | |
| Fig. 2: | (A) The effect of the chemical lesion with ibotenic acid on increased mean cell hemoglobin concentration (**p<0. 04 vs. sham-operated group), (B) platelet (**p<0.03 vs. sham-operated group) and (C) red cell distribution width (p<0.03 vs. sham-operated group) all tested one week after lesion; *all p-values of control vs. sham-operated group (p>0.05). Values are Mean±SEM (n = 15 per group) |
Among the catecholamines, a substantial amount of dopamine was detected in bone marrow (Marino et al., 1997). Bone marrow catecholamines originate from sympathetic nerve fibers and from hematopoietic cells directly. Catecholamines of neural origin show a circadian rhythmicity. Adrenoceptors present on bone marrow cells include the 1-subtype, which seems to mediate the catecholaminergic ShO of hematopoiesis.
Kaizer et al. (2008) have suggested that AChE activity in erythrocytes may be considered a marker of easy access of the central cholinergic status. Many researches showed that the bone marrow acetylcholine can be released by the stromal elements affecting generation and differentiation of blood cells. In the cited work nicotine affected hematopoiesis supportive stromal microenvironment, interfering with the stem cell homing. Later it was shown that nicotinic receptors are expressed in the very hematopoietic cell precursors (Serobyan et al., 2007; Koval et al., 2008).
It has been well documented that there are α- and β-adrenergic receptors on the surface of the erythrocytes. However, β-receptors are more important in erythropoiesis. Administration of an α-adrenergic stimulant did not elicit the erythropoietic effect, whereas a β-adrenergic stimulant induced erythropoiesis in an in vitro culture of erythroid progenitor cells (Mladenovic and Adamson, 1984). Since β-adrenergic receptors are abundant on the surface of erythrocytes, the effect of α-adrenergic blocking agents has often been investigated using erythrocytes (Miklavc et al., 1989; Hritcu, 2006). In accordance with these findings, in our present study we observed some abnormalities of hematopoiesis after electrolytic lesion of the central dopaminergic neurons from the substantia nigra by means of ibotenic acid. Ibotenic acid is a useful chemical agent for inducing neurogenic anemia.
Serum levels of methylmalonic acid rise in B12 deficiency, whereas homocysteine levels rise in both folate and B12 deficiency (Andres et al., 2008). Hematological features of B12 and folate deficiencies include anemia, macrocytosis, thrombocytopenia, neutropenia and neutrophil hypersegmentation. Recently elevated serum homocysteine in patients with Alzheimer's Disease (AD) has been described (McCaddon et al., 1998). Other studies have confirmed this observation and suggested that elevated homocysteine is a risk factor for cognitive decline and dementia (McCaddon et al., 2001, 2002; Seshadri et al., 2002; Seshadri, 2006; Schulz, 2007).
In conclusion, this study confirms earlier observations of increased plasma homocysteine and reduced serum folate levels in patients with clinically diagnosed AD, Hb and platelet counts fell only slightly with increasing dementia duration, also there were other changes in hematological indices; macrocytosis and RDW in particular were related to disease duration and there were anemic subjects. Thus, in present study sample, low Hb and RBC indices may have increased susceptibility to AD. The potential role of this changes in hematological parameters in increasing risk of AD should be explored prospectively, in both developed and developing countries, considering its high prevalence in older adults and the intriguing possibility that low Hb and RBC indices could be a modifiable risk factor.
ACKNOWLEDGMENT
This study (PRC44) was supported by Ahwaz Jondishapour University of Medical Sciences, Ahwaz, Iran.
REFERENCES
- Ahmed, M.M., H. Hoshino, T. Chikuma, M. Yamada and T. Kato, 2004. Effect of memantine on the levels of glia cells, neuropeptides and peptide-degrading enzymes in rat brain regions of ibotenic acid-treated Alzheimer's disease model. Neuroscience, 126: 639-649.
PubMed - Alaei, H., R. Moloudi and A. Sarkaki, 2008. Effects of treadmill running on mid-term memory and swim speed in the rat with Morris water maze test. J. Bodyw. Mov. Ther., 12: 72-75.
CrossRefPubMedDirect Link - Andres, E., T. Vogel, L. Federici, J. Zimmer, E. Ciobanu and G. Kaltenbach, 2008. Cobalamin deficiency in elderly patients: A personal view. Curr. Gerontol. Geriatr. Res., 2008: 848267-848267.
PubMed - Arendash, G.W., W.J. Millard, A. Dunn and E.M. Meyer, 1987. Long-term neuropathological and neurochemical effects of nucleus basalis lesions in the rat. Science, 238: 952-956.
PubMed - Arredondo, J., V.T. Nguyen, A.I. Chernyavsky, D. Bercovich, A. Orr-Urtreger, D.E. Vetter and S.A. Grando, 2003. Functional role of alpha 7 nicotinic receptors in physiological control of cutaneous homeostasis. Life Sci., 72: 2063-2067.
Direct Link - Beard, C.M., E. Kokmen, P.C. O'Brien, B.J. Ania and L.J. Melton, 1997. Risk of Alzheimer's disease among elderly patients with anemia: Population-based investigations in Olmsted County, Minnesota. Ann. Epidemiol., 7: 219-224.
PubMed - Bennekou, P., 1993. The voltage-gated non-selective cation channel from human red cells is sensitive to acetylcholine. Biochim. Biophys. Acta, 1147: 165-167.
PubMed - Conti-Fine, B.M., D. Navaneetham, S. Lei and A.D.J. Maus, 2000. Neuronal nicotinic receptors in non-neuronal cells: New mediators of tobacco toxicity. Eur. J. Pharmacol., 393: 279-294.
PubMed - Cormier, A., Y. Paas, R. Zini, J.P. Tillement, G. Lagrue, J.P. Changeux and R. Grailhe, 2004. Long-term exposure to nicotine modulates the level and activity of acetylcholine receptors in white blood cells of smokers and model mice. Mol. Pharmacol., 66: 1-7.
PubMed - Desforges, J.F., 1984. Regulation of hematopoiesis: 14th annual paul aggeler memorial lecture. West J. Med., 141: 88-92.
Direct Link - Dent, G.W., M.A. Smith and S. Levine, 2001. Stress-induced alterations in locus coeruleus gene expression during ontogeny. Dev. Brain Res., 127: 23-30.
PubMed - Etienne, P., Y. Robitaille, P. Wood, S. Gauthier, N.P.V. Nair and R. Quirion, 1986. Nucleus basalis neuronal loss, neuritic plaques and choline acetyltransferase activity in advanced Alzheimer's disease. Neuroscience, 19: 1279-1291.
PubMed - Felten, D.L., 1993. Direct innervation of lymphoid organs: Substrate for neurotransmitter signaling of cells of the immune system. Neuropsychobiology, 28: 110-112.
PubMed - Hefco, V., K. Yamada, A. Hefco, L. Hritcu, A. Tiron and T. Nabeshima, 2003. Role of the mesotelencephalic dopamine system in learning and memory processes in the rat. Eur. J. Pharmacol., 475: 55-60.
PubMed - Hoogduijn, M.J., Z. Rakonczay and P.G. Genever, 2006. The effects of anticholinergic insecticides on human mesenchymal stem cells. Toxicol. Sci., 94: 342-350.
PubMed - Hritcu, L., 2006. Effects of beta-adrenergic receptor blockade on stressinduced changes in haematological parameters of rats. Turk. J. Hematol., 23: 90-93.
Direct Link - Huang, M.X. and W.J. Liu, 2009. Effect of cluster a in hox gene on proliferation and differentiation of hematopoietic stem/progenitor cells and its relation to leukemia-review. Zhongguo Shi Yan Xue Ye Xue Za Zhi., 17: 835-839.
PubMed - Kaizer, R.R., M.C. Correa, R.M. Spanevello, V.M. Morsch, C.M. Mazzanti, J.F. Goncalves and M.R.C. Schetinger, 2005. Acetylcholinesterase activation and enhanced lipid peroxidation after longterm exposure to low levels of aluminum on different mouse brain regions. J. Inorganic Biochem., 99: 1865-1870.
PubMed - Kaizer, R.R., M.C. Correa, L.R.S. Gris, C.S. da Rosa, D. Bohrer, V.M. Morsch and M.R.C. Schetinger, 2008. Effect of long-term exposure to aluminum on the cetylcholinesterase activity in the central nervous system and erythrocytes. Neurochem. Res., 33: 2294-2301.
PubMed - Katz, I.R., P. Beaston-Wimmer, P. Parmelee, E. Friedman and M.P. Lawton, 1993. Failure to thrive in the elderly: Exploration of the concept and delineation of psychiatric components. J. Geriatr. Psychiatry Neurol., 6: 161-169.
PubMed - Kawashima, K. and T. Fujii, 2003. The lymphocytic cholinergic system and its biological function. Life Sci., 72: 2101-2109.
PubMed - Keikhaei, B., F. Rahim, K.H. Zandian and M. Pedram, 2007. Comparison of different indices for better differential diagnosis of iron deficiency anemia from β-thalassemia trait. Scient. J. Iran. Blood Transfusion Organiz. (Khoon), 4: 95-104.
Direct Link - Khaldoyanidi, S., I. Sikora, I. Orlovskaya, V. Matrosova, V. Kozlov and P. Sriramarao, 2001. Correlation between nicotine-induced inhibition of hematopoiesis and decreased CD44 expression on bone marrow stromal cells. Blood, 98: 303-312.
CrossRefPubMedDirect Link - Koval, L.M., A.S. Zverkova, R. Grailhe, Y.N. Utkin, V.I. Tsetlin, S.V. Komisarenko and M.V. Skok, 2008. Nicotinic acetylcholine receptors alpha4beta2 and alpha7 regulate myelo- and erythropoiesis within the bone marrow. Int. J. Biochem. Cell Biol., 40: 980-990.
PubMed - Maestroni, G.J., 1998. Is hematopoiesis under the influence of neural and neuroendocrine mechanism. Histol. Histopathol., 13: 271-274.
PubMed - Marino, F., M. Cosentino, R. Bombelli, M. Ferrari and G.J. Maestroni et al., 1997. Measurement of catecholamines in mouse bone marrow by means of HPLC with electrochemical detection. Haematologica, 82: 392-394.
PubMed - McCaddon, A., G. Davies, P. Hudson, S. Tandy and H. Cattell, 1998. Total serum homocysteine in senile dementia of Alzheimer type. Int. J. Geriatr. Psychiatry, 13: 235-239.
PubMed - McCaddon, A., P. Hudson, G. Davies, A. Hughes, J.H. Williams and C. Wilkinson, 2001. Homocysteine and cognitive decline in healthy elderly. Dement. Geriatr. Cogn. Disord., 12: 309-313.
PubMed - McCaddon, A., B. Regland, P. Hudson and G. Davies, 2002. Functional vitamin B12 deficiency and Alzheimer disease. Neurology, 58: 1395-1399.
PubMed - McCaddon, A., S. Tandy, P. Hudson, R. Gray, G. Davies, D. Hill and J. Duguid, 2004. Absence of macrocytic anaemia in Alzheimer's disease. Clin. Lab. Haematol., 26: 259-263.
PubMed - Meyer, E.M., G.W. Arendash, J.H. Judkins, L. Ying, C. Wade and W.R. Kern, 1987. Effects of nucleus basalis lesion on the muscarinic and nicotinic modulation of [3H]-acetyl choline release from the rat cerebral cortex. J. Neurochem., 49: 1758-1762.
CrossRef - Miranda, M.I., G. Ferreira, L. Ramirez-Lugo and F. Bermudez-Rattoni, 2003. Role of cholinergic system on the construction of memories: Taste memory encoding. Neurobiol. Learn. Mem., 80: 211-222.
PubMed - Mladenovic, J. and J.W. Adamson, 1984. Adrenergic modulation of erythropoiesis: In vitro studies of colony-forming cells in normal and polycythaemic man. Br. J. Haematol., 56: 323-332.
PubMed - Miklavc, A., D. Kocjan, D. Hadzi, J. Mavri and J. Koller, 1989. Binding of agonists and antagonists to beta-adrenergic receptor. Prog. Clin. Biol., 291: 275-280.
PubMed - Nanda, N., G.K. Pal and B.C. Koner, 2005. Effect of dopamine injection into caudate nucleus on immune responsiveness in rats: A pilot study. Immunol. Lett., 96: 151-153.
PubMed - Pacheco-Lopez, G., M.B. Niemi, W. Kou, A. Bildhauser and C.M. Gross et al., 2003. Central catecholamine depletion inhibits peripheral lymphocyte responsiveness in spleen and blood. J. Neurochem., 86: 1024-1031.
PubMed - Pandav, R.S., V. Chandra, H.H. Dodge, S.T. DeKosky and M. Ganguli, 2004. Hemoglobin levels and Alzheimer disease: An epidemiologic study in India. Am. J. Geriatr. Psychiatry, 12: 523-526.
PubMed - Paterson, D. and A. Nordberg, 2000. Neuronal nicotinic receptors in the human brain. Prog. Neurobiol., 61: 75-111.
CrossRef - Salamone, J.D., P.M. Beart, J.E. Alpert and S.D. Iverson, 1984. Impairment in T-maze reinforced alternation performance following nucleus basalis magnocellularis lesions in rat. Behav. Brain Res., 13: 63-70.
PubMed - Schulz, R.J., 2007. Homocysteine as a biomarker for cognitive dysfunction in the elderly. Curr. Opin. Clin. Nutr. Metab. Care, 10: 718-723.
PubMed - Serobyan, N., I. Orlovskaya, V. Kozlov and S.K. Khaldoyanidi, 2005. Exposure to nicotine during gestation interferes with the colonization of fetal bone marrow by haematopoietic stem/progenitor cells. Stem Cells Dev., 14: 81-91.
PubMed - Serobyan, N., S. Jagannathan, I. Orlovskaya, I. Schraufstatter, M. Skok, J. Loring and S. Khaldoyanidi, 2007. The cholinergic system is involved in regulation of the development of the hematopoietic system. Life Sci., 80: 2352-2360.
PubMed - Seshadri, S., 2006. Elevated plasma homocysteine levels: Risk factor or risk marker for the development of dementia and Alzheimer's disease? J. Alzheimers Dis., 9: 393-398.
PubMed - Siarakas, S., E. Damas and W.G. Murrell, 1997. The effect of enteric bacterial toxins on the catecholamine levels of the rabbit. Pathology, 29: 278-285.
PubMed - Spreng, M., S. Cotecchia and F. Schenk, 2001. A behavioral study of alpha-1b adrenergic receptor knockout mice: Increased reaction to novelty and selectively reduced learning capacities. Neurobiol. Learn. Mem., 75: 214-229.
PubMed - Wang, H., M. Yu, C.A. Amella, M. Ochani and M. Tanovic et al., 2003. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature, 421: 384-388.
CrossRefPubMedDirect Link - Widy-Tyszekoeiwecz, E., A. Peichal, B. Gajkowska and M. Smailek, 2002. Tellurium-induced cognitive deficits in rats are related to neuropathological changes in the central nervous system. Toxocol. Lett., 131: 203-214.
PubMed - Wu, C.K., L. Thal, D. Pizzo, L. Hansen, E. Masliah and C. Geula, 2005. Apoptotic signals within the basal forebrain cholinergic neurons in Alzheimer's disease. Exp. Neurol., 195: 484-496.
PubMed - Yamamoto, M., T. Chikuma and T. Kato, 2003. Changes in the levels of neuropeptides and their metabolizing enzymes in the brain regions of nucleus basalis magnocellularis-lesioned rats. J. Pharmacol. Sci., 92: 400-410.
PubMed - Zatta, P., M. Ibn-Lkhayat-Idrissi, P. Zambenedetti, M. Kilyen and T. Kiss, 2002. In vivo and in vitro effects of aluminum on the activity of mouse brain acetylcholinesterase. Brain Res. Bull., 59: 41-45.
CrossRefPubMedDirect Link