
Osama M. Ahmed
Department of Zoology, Faculty of Science, Beni-Suef University, Beni-Suef, Egypt
Gamal Allam
Department of Zoology, Faculty of Science, Beni-Suef University, Beni-Suef, Egypt
Asian Journal of Animal and Veterinary Advances
Year: 2009 | Volume: 4 | Issue: 4 | Page No.: 172-190







ABSTRACT
The effect of Vasoactive Intestinal Peptide (VIP) on Insulin Dependent Diabetes Mellitus (IDDM) and schistosomiasis together in combination has not been previously investigated. To assess its efficacy in such condition, VIP was administered to Schistosoma mansoni-infected streptozotocin-induced diabetic (ID) mice at a dose level of 41.6 ng kg–1 b.wt., 3 times/week, for 8 consecutive weeks starting from the 1st week of infection. The administration of VIP to ID mice induced a potential amelioration of serum glucose, insulin and C-peptide levels indicating the insulinogenic effect of this peptide. VIP also produced a significant decrease of hepatic granuloma volume and worm fecundity in the ID mice without affecting worm burden. The granuloma volume was found to be lower in the ID mice as compared to that of the infected non-diabetic ones. VIP administration produced marked decreases of the elevated liver collagen, serum carbohydrate antigen (CA.19.9) and liver alpha fetoprotein (AFP) content of ID mice as well as it succeeded, at least partially, to alleviate the altered liver enzyme activities. It also successfully increased the anti-inflammatory cytokine, IL-10 and decreased the elevated pro-inflammatory chemokines, IL-12 and TNF-α level in the serum of ID mice. These changes in cytokines explain the decrease in hepatic granuloma volume and reflect the anti-inflammatory effects of VIP. The increased oxidative stress markers and perturbed antioxidant defense system were profoundly improved in the ID mice treated with VIP. In conclusion, the VIP may have anti-hyperglycemic and insulinotropic effects, decrease liver and intestinal egg count and ameliorate liver pathologic deteriorations via its immunomodulatory effects on cytokines released from macrophages and T helper cells in addition to its improvement effect on the antioxidant defense system of the infected diabetic mice.
PDF Abstract XML References Citation
How to cite this article
Osama M. Ahmed and Gamal Allam, 2009. The Significance of Vasoactive Intestinal Peptide in the Treatment of Schistosoma mansoni-Infected Diabetic Mice. Asian Journal of Animal and Veterinary Advances, 4: 172-190.
DOI: 10.3923/ajava.2009.172.190
URL: https://scialert.net/abstract/?doi=ajava.2009.172.190
DOI: 10.3923/ajava.2009.172.190
URL: https://scialert.net/abstract/?doi=ajava.2009.172.190
INTRODUCTION
Insulin-Dependent Diabetes Mellitus (IDDM) or type 1 diabetes, characterized by absolute insulin deficiency, is associated with cellular infiltration and an inflammatory response in the islets of Langerhans (Nielsen et al., 1999). Cellular components of this infiltration include monocytes, macrophages and CD4+ and CD8+ T-cells (Mandrup-Poulsen, 1996). The balance between T-helper (Th)-1 type and T-helper (Th)-2 is crucial in the pathogenesis of IDDM; thus, a bias toward the proinflammatory Th-1 cytokines promotes insulitis and IDDM (Amirshahrokhi et al., 2008). Released cytokines, interleukin 1β, Tumor Necrosis Factor (TNF)-α and interferon (IFN)-γ, exert cytotoxic effects specifically on β-cells in the islets of Langerhans, in part via., the induction of free radicals production (Kubisch et al., 1997). Th2 anti-inflammatory cytokines, IL-4 and IL-10, have been found to suppress insulitis, β-cell damage and IDDM in Non-Obese Diabetic (NOD) mice (Cameron et al., 1997) and in Multiple-Low-Dose Streptozotocin (MLDS)-induced diabetic mice (Amirshahrokhi et al., 2008).
Schistosomiasis remains to be one of the most prevalent parasitic infections and has significant public health consequences (Engels et al., 2002). Similarly to IDDM, the regions of inflammation in schistosomiasis, granulomas, contain macrophages and CD4+ and CD8+ T-cells in addition to eosinophils (Caldas et al., 2008). Both Th1 and Th2 cytokines can regulate granuloma formation. Initially, the host develops a Th1 response, which is characterized by the production of IL-1, IFN-γ, TNF-α and nitric oxide (De Jesus et al., 2002; Caldas et al., 2008). However, as the infection progresses, this Th1 response gradually succumbs to a dominant Th2 response induced by the parasite eggs' antigens and is characterized by the secretion of IL-4, IL-5, IL-10 and IL-13 (Grzych et al., 1991; Sher et al., 1991) resulting in diminished inflammation (Araujo et al., 2004). IDDM has been described to affect schistosomiasis in laboratory animals by different authors. Several experiments performed by Mahmoud et al. (1975, 1976, 1979) revealed a smaller size of granulomas in the chemically-induced and genetic diabetic mice infected with S. mansoni. Moreover, Hulstijn et al. (2001) found lower faecal egg excretion in STZ-induced diabetic mice infected with S. mansoni.
Vasoactive Intestinal Peptide (VIP) is a neuropeptide synthesized by immune cells that can modulate several immune aspects, including the function of cells involved in the inflammatory response (Metwali et al., 2002). It was reported that VIP has several functions: (1) Promotes Th2 differentiation and inhibits Th1 responses, (2) inhibits in vitro and in vivo production of the proinflammatory cytokines TNF-α, IL-2, IL-6, IL-12, IFN-γ and of Nitric Oxide (NO) and (3) stimulates the production of the anti-inflammatory cytokine IL-10 (Xin and Sriram, 1998; Delgado et al., 1999a-d, 2002). Granulomas in murine schistosomiasis mansoni contain VIP (Weinstock and Blum, 1990) and contain VIP-responsive lymphocytes (Weinstock et al., 1991). Administration of VIP to S. mansoni-infected mice reduces liver pathology induced by schistosome eggs, inhibits IL-12 and TNF-α production and stimulates IL-10 production (Allam, 2007). In addition to the immunoregulatory role in schistosomiasis, VIP was found to evoke profound increases in insulin secretion from the pancreas of diabetic rats (Adeghate et al., 2001) and mice (Ahmed, 2009).
Pathogenesis of auto-immune diseases, including IDDM, mediated by T cells involves the type I immune response with production of IFN-γ and TNF-α. However, egg antigens of S. mansoni infection induce a strong Th2-type cytokine response with increased production of IL-10 and prevent the onset of type 1 (autoimmune) diabetes (Zaccone et al., 2003). Thus, schistosomiasis may affect the induction of the immune-mediated diabetes mellitus and vice versa (El-Wakil et al., 2002). Thus, this study was designed to evaluate the significance of VIP in the treatment of diabetic mice infected with S. mansoni.
MATERIALS AND METHODS
Animals
Male 8-week old CD1 albino mice were purchased from the Schistosome Biological Supply Centre, Theodore Bilharz Research Institute (TBRI), Imbaba, Giza, Egypt. Mice were maintained under suitable living conditions in the animal house (Zoology Department, Faculty of Science, Beni-Suef University, Egypt) at controlled temperature (20-25°C) and natural daily 12 h light-dark cycles throughout the experiment. They were supplied with food pellets, which contain at least 25.4% protein and tap water ad libitum. They were also kept under observation for 1 week before the onset of the experiment to exclude any intercurrent infection and for proper acclimatization. All animal procedures follow the animal research bioethics of SBSC, TBRI and are in accordance with the recommendations for the proper care and use of laboratory animals (CCAC, 1993).
Induction of Insulin-Dependent Diabetes Mellitus
Experimental diabetes mellitus was induced in overnight fasted CD1 albino mice by intraperitoneal injection of streptozotocin (STZ) (Sigma Chemical Company, USA) at dose level of 150 mg kg–1 b.wt. dissolved in citrate buffer (pH 4.5) (Miyamoto et al., 2008). Ten days after STZ injection, blood samples were obtained from the lateral tail vein of mice that had been deprived of food for 10-12 h and after 2 h post-oral glucose loading (3 g kg–1 b.wt.). Mice having 2 h serum glucose concentration that ranged from 180 to 300 mg dL–1 were considered mild diabetics and were included in the experiment.
Parasite and Infection
Schistosoma mansoni (John Bruce Egyptian strain) cercariae were obtained from infected Biomphalaria alexandrina snails (TBRI) as previously described by Schubert (1948). Normal and STZ-diabetic mice were infected with 40 live cercariae for each individual according to Stirewalt and Bronson (1955) and Bruce and Radke (1971).
Experimental Design
Synthetic porcine VIP (Sigma Chemical Company, USA) was reconstituted in sterile 0.9% saline-0.1% gelatin containing 0.5% albumin as described by Karlsson and Ahren (1990). The peptide was divided into aliquots and immediately stored at -70°C until use. Infected Diabetic Treated (IDT) mice were injected intraperitoneally with VIP at a dose of 41.6 ng kg–1 b.wt. 3 injections/week for 8 consecutive weeks, starting from the first week of infection; the total dose at the end of the experiment was 1 μg kg–1 b.wt. (Allam, 2007; Ahmed, 2009). The other groups [naive (N), infected only (I) and Infected Diabetic (ID) mice] were given the same amount of the vehicle at the same time as treated groups.
Parasitological Analysis
At the 8th week post-infection, worm burdens were estimated by portal perfusion according to Duvall and DeWitt (1967). The numbers of S. mansoni eggs in the perfused liver and in the intestine were estimated after alkali digestion as described by Cheever (1968).
Blood and Tissue Sampling
Sera were collected from the clotted blood samples after centrifugation at 3000 rpm for 15 min, then divided into aliquots and stored at -70°C until use. The liver from each mouse was excised immediately after perfusion and rinsed with ice-cold saline solution. The ventral median lobe of the liver was fixed in 10% neutral buffered formalin for preparation of hematoxylin and eosin-stained sections. Another portion was homogenized by Teflon homogenizer (Glas-Col, USA) in 0.9% saline (0.5 g tissue/5 mL saline), centrifuged at 3000 rpm for 15 min and the supernatant was decanted and stored at -70°C until use for different biochemical estimations.
Histopathological Analysis
The diameters of granulomas surrounding schistosome eggs were measured in the hematoxylin and eosin-stained liver sections by using an ocular micrometer (Carl Zeiss, Thornwood, NY, USA). Twenty granulomas per mouse were measured and the largest diameter and that perpendicular to it were averaged (Lichtenberg, 1962). The volume (mm3) of each granuloma was calculated assuming a spherical shape using the following formula: Volume of sphere = 3πR3 (the radius R was obtained by dividing the main diameter of the lesion by two) and the mean volume for each group was calculated (Mahmoud and Warren, 1974).
Immunological Analysis
Cytokines level was measured by using sandwich Enzyme-Linked Immunosorbent Assay (ELISA) in the Virology Research Laboratory (Faculty of Veterinary, Beni-Suef University, Egypt). Cytokine concentrations (pg mL–1) for mouse IL-10, IL-12 +p40 and TNF-α were determined with commercially available reagents and ELISA kits purchased from BioSource International (Camarillo, California, USA). Cytokine concentrations were determined using a standard curve obtained from the known concentration of cytokine standards included in each assay plate according to manufacture’s instructions.
Biochemical Investigations
All biochemical determinations were performed in the Zoology Department Research Laboratory (Faculty of Science, Beni-Suef University, Egypt) except for those mentioned elsewhere. Humalyzer 2000 Chemistry Analyzer (HUMAN Gesellschaft für Biochemica und Diagnostica mbH, Wiesbaden, Germany) was used for spectrophotometric and colorimetric determinations.
Serum glucose concentration (mg dL–1) was determined according to the enzymatic method described by Siest and Schielef (1981) using reagent kits obtained from BioMerieux Chemical Company (France). Serum insulin (μIU mL–1) and C-peptide (pmol L–1) concentrations were assayed in the Radioactive Isotopes Unit, National Research Centre (Dokki, Cairo, Egypt) using radioimmunoassay kits supplied by Diagnostic Products Corporation (Los Angeles, USA) according to the methods of Marschner et al. (1974) and Bonser and Garcia-Webb (1984), respectively.
Hepatic alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities (mU/100 mg tissue) in the homogenate supernatant were estimated according to the kinetic method of Bergmyeyer et al. (1978), using reagent kits purchased from Spinreact Company (Spain). Hepatic γ-glutamyl transferase (γ-GT) activity (mU g–1 tissue) was determined according to the kinetic method of Young (1990) and Tietz (1991) using reagent kits obtained from BioSystems (Spain). Liver LDH activity (mU/100 mg tissue) was determined according to the method described by Buhl and Jackson (1978), using reagent kits purchased from Stanbio Laboratories (Texas, USA).
The amount of collagen (μg/100 mg tissue) in liver samples was determined from its hydroxyproline content by the method of Neuman and Logan (1950a, b). Briefly, the defatted and dried samples were hydrolyzed by autoclaving in 2 mL of 6 N HCl at 50 pounds pressure. The protein hydrolysates were used for the determination of hydroxyproline and the percentage of collagen of a tissue sample was calculated by using the 7.46 factor of Neuman and Logan (1950a) and Baykal-Erkilic et al. (1995). Meanwhile, a standard curve was constructed in order to determine the reproducibility of the assay.
The levels of hepatic reduced glutathione (GSH) (nmol/100 mg tissue), total thiol (nmol/100 mg tissue) and lipid peroxidation products (nmol MDA/100 mg tissue) were determined by the methods of Beutler et al. (1963), Koster et al. (1986) and Preuss et al. (1998), respectively, using reagents prepared in the laboratory. Liver catalase (k.102) level was determined according to the technique of Cohen et al. (1970) using reagents prepared in the laboratory. Results of catalase activity were expressed in terms of the first-order reaction rate constant (k) as indicated by Cohen et al. (1970). Heme peroxidases (myeloperoxidase and eosinophil peroxidase) activity (U g–1 tissue) in the liver of naive and infected diabetic mice and infected diabetic mice treated with VIP was assayed according to the methods of Chance and Maehley (1955) and Kar and Mishra (1976) with some modifications, using pyrogallol as a substrate (Conyers and Kidwell, 1991; Joseph et al., 1992) in the presence of H2O2. To 25 μL of supernatant, 2 mL phosphate buffer solution (pH 6.8), 100 μL 1.4% pyrogallol and 100 μL 1.1 mmole H2O2 were added. After exactly 5 min, the density of colour of formed purpurogallin was measured against the blank by recording absorbance at 420 nm. The enzyme activity (in units) for each sample was obtained from the standard curve performed by using different dilutions of heme peroxidase (EC.1.11.1.7) (Sigma-Aldrich Company, USA).
Serum CA 19.9 concentration (U mL–1) was estimated by an immunoradiometric assay kit (Coat-A-Count GI-MA IRMA, DPC, USA) according to the method of Frebourg et al. (1988). α-Fetoprotien (AFP) concentration (U g–1 tissue) in liver homogenate supernatant was determined by a radioimmunoassay (RIA) kit (Double antibody kit, DPC, USA) according to the methods of Waldmann and Mclntire (1974) and Krikpatrick et al. (1977). Both CA19.9 and AFP concentrations were measured in the Radioactive Isotopes Unit, National Research Center (Dokki, Cairo, Egypt).
Statistical Analysis
The data were analyzed using one way Analysis of Variance (ANOVA) followed by LSD analysis to compare various groups with each other using PC-STAT program, University of Georgia, USA (Rao et al., 1985). The results were expressed as Mean±SE and values of p>0.05 were considered statistically insignificant, while those of p<0.05 and p<0.01 were considered statistically significant and highly significant respectively. F-probability for each variable expressed the general effect between groups.
RESULTS
Worm Burden
As shown in Fig. 1, there was no significant (p>0.05) difference in worm burden of the different infected groups.
Granuloma Volume
Granuloma volume was measured in hematoxylin and eosin-stained liver sections of 8 weeks I, ID and IDT mice. The data of the present study showed that, VIP treatment of ID mice highly significantly reduced (p<0.01) hepatic granuloma volume by 84.7 and 60.2% as compared with I and ID mice, respectively. Also, ID mice showed 61.6% significant (p<0.01) reduction in hepatic granuloma volume compared to I mice (Fig. 1). One-way ANOVA indicated that the effect between groups was very highly significant (p<0.001) throughout the experiment.
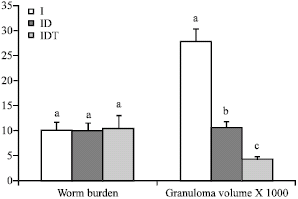 | |
| Fig. 1: | Effect of VIP treatment on worm burden (number of worms) and hepatic granuloma volume (mm3) of S. mansoni-infected (I), infected diabetic (ID) and infected diabetic VIP treated (IDT) mice. Values represented as the Mean±SE of ten mice. Columns not sharing common superscript(s) denote significant differences. F-probability: p>0.05 for worm burden and p<0.01 for granuloma volume |
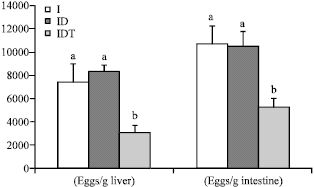 | |
| Fig. 2: | Effect of VIP treatment on tissue egg load of infected (I), infected diabetic (ID) and infected diabetic VIP treated (IDT) mice. Values represented as the Mean±SE of ten mice. Columns not sharing common superscripts denote significant differences. F-probabilitiy: p<0.05 |
Liver and Intestine Egg Load
After the perfusion had been completed, about 0.5 g of both liver and intestine for each animal was digested in 5 mL 5% KOH overnight at 37°C. Eggs were counted in triplicates in liver and intestine homogenates for each mouse by using MacMaster counting slide and mean eggs per gram liver and intestine was calculated. VIP treatment of infected diabetic mice led to significant decrease (p<0.05) in the liver egg load which was 58.6 and 63.5 % as compared with infected non-diabetic and infected diabetic mice, respectively. Similarly, the intestine egg load of IDT mice showed 51.2 and 50% significant (p<0.05) reduction as compared to I and ID mice, respectively (Fig. 2). With regards to one-way ANOVA, it was found that the general effect between groups on both liver and intestine egg counts was only significant (p<0.05) throughout the experiment.
Level of Cytokines
Serum levels of IL-10, IL-12 and TNF-α were quantified by using sandwich ELISA. As depicted in Fig. 3, IL-10 level of IDT mice showed about 9- and 2-folds significant (p<0.01) increase compared to those of N and ID mice, respectively, at 8 weeks post-infection and treatment. Also, ID mice showed about 5-fold significant (p<0.01) increase in IL-10 level compared to N mice. However, IL-12 and TNF-α levels of ID mice were significantly increased as compared to N group. The administration of VIP to ID mice induced a profound decrease (p<0.01) of the elevated IL-12 and TNF-α levels (Fig. 3). With regards one-way ANOVA, it was found that the effect between groups on serum IL-12 and IL-10 was highly significant (p<0.01), while the effect on TNF-α was only significant (p<0.05) throughout the experiment.
Biochemical Results
The serum glucose concentration, at fasting state and after 2 h of oral glucose loading, was tremendously increased (p<0.01) in schistosoma-infected diabetic mice. The administration of VIP 3 times/week for 8 consecutive weeks to the ID mice induced a potential amelioration of the hyperglycemia. Fasting serum insulin and C-peptide levels, on the other hand, were remarkably decreased (p<0.01) in the infected diabetic mice. The administration of VIP profoundly ameliorated (p<0.01) the lowered levels of these parameters. One-way ANOVA depicted that the general effect between groups on serum glucose, insulin and C-peptide levels was very highly significant (p<0.001) throughout the experiment (Table 1).
With the exception of liver LDH activity, which showed a significant increase, the other tested liver enzyme activities were significantly diminished in the ID mice. The altered levels of LDH and γ-GT activities were normalized by VIP treatment.
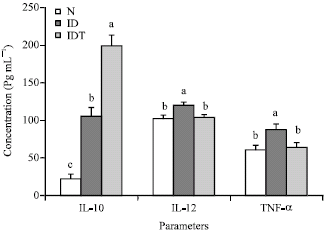 | |
| Fig. 3: | Effect of VIP treatment on IL-10, IL-12 and TNF-α serum levels of infected diabetic (ID) and infected diabetic VIP treated (IDT) mice as compared to normal (N). Values represented as the Mean±SE of 8 mice. Columns not sharing common superscripts denote significant differences: F-probability: p<0.05 for TNF-α and p<0.01 for IL-10 and IL-12 levels |
| Table 1: | Effect of VIP administration on serum glucose, insulin and C-peptide levels of S. mansoni-infected diabetic mice |
 | |
| Data are expressed as Mean±SE. Number of animals in each group is ten. Mean values, which share the same superscript symbol(s) are not significantly different | |
| Table 2: | Effect of VIP administration on liver enzyme activities of S. mansoni-infected diabetic mice |
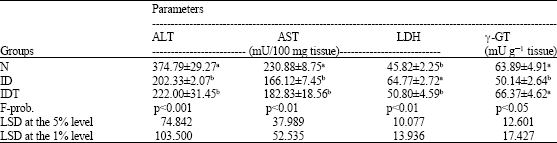 | |
| Data are expressed as Mean±SE. Number of animals in each group is ten. Mean values, which share the same superscript symbol(s) are not significantly different | |
One-way ANOVA depicted that the general effect between groups on the liver enzymes was at least significant throughout the experiment (Table 2).
The liver collagen, serum CA 19.9 and liver α-fetoprotein levels were highly significantly elevated (p<0.01) in the ID mice. The VIP administration normalized the elevated level of liver collagen, while it significantly decreased serum CA 19.9 and liver α-fetoprotein levels below the normal one. One-way ANOVA depicted that while general effect between groups on liver collagen was highly significant (p<0.01), the effect on serum CA 19.9 and α-FP was very highly significant (p<0.001) throughout the experiment (Table 3).
| Table 3: | Effect of VIP administration on liver collagen and AFP and serum CA19.9 levels of S. mansoni-infected diabetic mice |
 | |
| Data are expressed as Mean±SE. Number of animals in each group is ten. Mean values, which share the same superscript symbol(s) are not significantly different | |
| Table 4: | Effect of VIP administration on various liver oxidative stress markers of S. mansoni-infected diabetic mice |
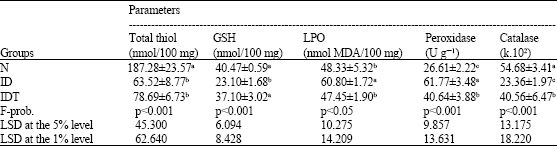 | |
| Data are expressed as Mean±SE. Number of animals in each group is ten. Mean values, which share the same superscript symbol(s) are not significantly different | |
Regarding the oxidative stress and antioxidant defense system (Table 4), liver total thiol, glutathione and catalase levels were tremendously decreased in the ID mice as compared to normal ones while the liver peroxidation and peroxidase activity were vigorously increased. The administration of VIP induced a detectable amelioration to various extents. The effect of VIP on liver total thiol content was non-significant, while its effect on other tested oxidative stress and antioxidant markers was at least significant. One-way ANOVA revealed that the effect between groups on liver lipid peroxidation was only significant (p<0.05), while the effect on the liver total thiol, glutathione, peroxidase and catalase levels was very highly significant (p<0.001) between the groups throughout the experiment.
DISCUSSION
Several recent reviews have recapitulated previous numerous studies demonstrating the involvement of VIP immune functions in the control of homeostasis and health. The widespread knowledge of the anti-inflammatory function of VIP has led investigators to suggest the use of VIP for the treatment of several inflammatory and autoimmune diseases including type I diabetes mellitus (Delgado et al., 2002, 2004; Abad et al., 2006; Gomariz et al., 2001, 2006, 2007; Gonzalez-Rey et al., 2007). IDDM is associated with cellular infiltration consisting of monocytes/macrophages and T-cells subsets (CD4+ and CD8+) and an inflammatory response in the islets of Langerhans (Nielsen et al., 1999). Similarly, schistosomiasis exhibited cellular filtration (eosinophils, macophages, CD4+ and CD8+ T cells) and an inflammation around the schistosome eggs entrapped in the liver. There are numerous studies showing that proinflammatory Th1 cytokines IL-1β, IFN-γ, TNF-α and NO are critically involved in the pathogenesis of IDDM and schistosomiasis (Rabinovitch, 1998; Caldas et al., 2008). The preponderance of the anti-inflammatory Th2 cytokines IL-4 and IL-10 in both diseases counteracts Th1 cytokine effects and may prevent liver damage or β-cells destruction (Allam, 2007; Amirshahrokhi et al., 2008). Thus, the two diseases at different stages may affect each other. Based on these findings, this study highlights this interaction by following the progression of each disease in the presence of the other. In addition, the effect of VIP on the two diseases together in combination as an animal model is assessed.
The data of the present study showed that the administration of VIP to ID mice significantly reduced serum level of pro-inflammatory cytokines, TNF-α and IL-12. These results are in agreement with the previous reports which stated that VIP inhibits the transcription (Delgado and Ganea, 1999) and production of pro-inflammatory cytokine IL-12 from macrophages (Xin and Sriram, 1998; Gomariz et al., 2000) and from Th1-cells (Wang et al., 1999) as well as it reduces the serum level of IL-12 in I mice (Allam, 2007). Delgado et al. (1998, 1999d, 1999e) also reported that VIP inhibits TNF-α production in vitro and in vivo from LPS-stimulated macrophages through the binding to VIP/pituitary adenylate cyclase-activating peptide (VPAC) receptor 1 that leads to an activation of the adenylate cyclase system and a reduction of the nuclear factor-κB (NF-κB) binding. Increase in cAMP as a result of the adenylate cyclase activation has been demonstrated to inhibit the expression of a variety of T-cell functions in schistosomiasis (Weinstock et al., 1991). In a earlier study, Allam (2007) showed that serum TNF-α level of I mice was potentially reduced with VIP treatment. On the other hand, the IDT mice exhibited a significant increase in the level of serum IL-10 as compared with N and ID mice. This finding confirmed the previous reports which have shown that VIP stimulates in vitro and in vivo production of the anti-inflammatory cytokine IL-10 (Delgado et al., 1999b; Gomariz et al., 2000; Allam, 2007; Larocca et al., 2007). It has been shown that in addition to increasing IL-10 production from Th2 cells, VIP treatment can increase production of regulatory T cells, which produce high levels of IL-10 (Chorny et al., 2005; Delgado et al., 2005; Reinke and Fabry, 2006). The stimulation of IL-10 production by VIP administration attenuates the production of pro-inflammatory cytokines including TNF-α and NO (Nicoletti et al., 1997). Consequently, VIP strongly reduces inflammatory response and exerts its anti-inflammatory function in several ways: (1) Direct inhibition of pro-inflammatory cytokine production (TNF-α, IL-6 and IL-12); (2) up-regulation of IL-10 production (a potent anti-inflammatory cytokine); (3) inhibition of expression and release of pro-inflammatory chemokines from activated macrophages and microglia; (4) inhibition of B7.1/B7.2 expression in activated macrophages and dendritic cells and subsequent inhibition of their stimulatory activity for antigen-specific T cells and (5) inhibition of Th1 responses (reduction in both the amounts of Th1 cytokines and the number of cytokine-producing Th1 cells) (Delgado et al., 2004).
In the current study, the serum glucose concentration at fasting state and after 2 h of oral glucose loading was profoundly elevated in the S. mansoni-infected diabetic mice as compared to the normal group. This change was associated with a significant deficiency of circulating insulin and C-peptide level, which was due to the deleterious damage of β-cells in the islets of Langerhans in the pancreas after streptozotocin (STZ) injection in these animals. Several mechanisms were reported by various authors to explain the action of streptozotocin to induce β-cells damage and in turn to decrease blood insulin and C-peptide levels. These mechanisms include: an increase in the oxidative stress and production of reactive oxygen and nitrogen species, an induction of DNA strand breaks, a depletion of NAD, an inhibition of ATP synthesis and/or a perturbed immune regulation (Okamoto, 1985; Pusztai et al., 1996; Amirshahrokhi et al., 2008). Moreover, Ahmed (2009) suggested that TNF-α, which was remarkably increased in STZ diabetic mice, may have a role in the β-cell destruction. Pehuet-Figoni et al. (1992) suggested that TNF-α could play a fundamental role in the progressive autoimmune destruction of β-cells in IDDM. Kaneto et al. (1995) stated that in the presence of TNF-α, large amounts of NO were produced and DNA cleavage occurred more noticeably in the islets of Langerhans. The NO-induced DNA cleavage is considered as an important step in the destruction and dysfunction of pancreatic β-cells induced by inflammatory stimulation or treatment with STZ (Kaneto et al., 1995). Also, Lukic et al. (1998) and Amirshahrokhi et al. (2008) reported that multiple-low-dose of STZ diabetes, a model of IDDM, is induced by Th1 lymphocytes that secrete soluble effector molecules that activate the macrophages and promote the destruction of β-cells possibly by both nitric oxide and non-nitric oxide - mediated mechanisms. In addition to the diabetogenic action of STZ, it was postulated that chronic and persistent elevated levels of glucose induce the rate of β-cell damage and apoptosis (Butler et al., 2003; Mellado-Gil and Aguilar-Diosdado, 2004; Cnop et al., 2005). The administration of VIP to diabetic mice just after infection for 8 consecutive weeks, in the present study, induced potential amelioration of the elevated serum glucose and the lowered serum insulin and C-peptide levels. These results are in accordance with those of Adeghate et al. (2001) and Ahmed (2009), who found that VIP improved the glycemic state and evoked an increase of insulin secretion associated with the increase in the number of β-cells in STZ-induced diabetic rats and mice. The parallel increase of serum C-peptide, a β-cell function marker, with serum insulin level, in the present study, support that VIP is an insulinotropic agent in the infected diabetic mice. It is relevant here to mention that the effect of VIP in the pancreatic islets to increase insulin secretion seems to occur via adenylate cyclase signaling pathway (Winzell and Ahren, 2007). The activation of adenylate cyclase leads to an elevation of cyclic adenosine monophosphate (cAMP) which in turn activates protein kinase A (PKA) to promote insulin secretion via several different pathways (Winzell and Ahren, 2007).
It is astonishing in the present study that the granuloma volume was greatly decreased in the S. mansoni-infected diabetic mice as compared with the infected non-diabetic ones although the worm burden and liver and intestine egg count were not significantly altered between these two groups. These changes agree with those of Mahmoud et al. (1975, 1976) and Mahmoud (1979), who reported that IDDM affects cell mediated response around schistosome eggs. These authors found that the diabetic infected animals revealed markedly suppressed lesions in granulomas similar to those in animals treated with powerful immunosuppressive agents. Also, MacCuish et al. (1974) found that lymphocytes from poorly controlled diabetics, in contrast to those from well-controlled patients have been found to respond poorly to phytohemoagglutinin stimulation. Since lymphocytes play a key role in cell-mediated immunologic responses, the alteration in the function of these cells in diabetes may in part explain the impairment in granuloma formation. Kazura et al. (1979) found that lymphoid cells of STZ and diabetic db/db mice have a reduced capacity to produce/secrete the lymphokine Eosinophil Stimulator Promoter (ESP) in response to soluble egg antigens of S. mansoni. In accordance with the present study, Hulstijn et al. (2001) depicted that the total amount of eggs in the intestine and the number of worms recovered was not affected in S. mansoni-infected diabetic mice in comparison with the infected non-diabetic ones. However, the data of the present study showed that the treatment of ID mice with VIP led to a significant decrease in liver and intestine egg loads. This result is in accordance with pervious studies that showed S. mansoni-infected mice treated with VIP had low tissue egg loads (Osman et al., 1997; Allam, 2007).
The present study indicated that the ID mice administered VIP showed a marked decrease in the granuloma size in comparison to the infected diabetic control. According to Weinstock et al. (1991), the granulomas around schistosome ova have eosinophils that produce VIP and T-lymphocytes that have VIP receptors. Therefore, it is possible that VIP may participate in the immunomodulation in the granulomas. It was postulated that VIP suppressed T lymphocytes proliferation in the granulomas by decreasing IL-2 and IL-4 production (Metwali et al., 1993; Sun and Ganea, 1993; Tang et al., 1995). Indeed, both IL-2 and IL-4 play a role in granuloma formation (Wynn et al., 1993; Cheever et al., 1994; Brunet et al., 1998). Also, the present study demonstrated that ID mice treated with VIP showed a high level of IL-10 and low level of TNF-α. Both cytokines play an important role in granuloma formation. Mice treated with IL-10 have smaller granulomas (Flores-Villanueva et al., 1996), whereas IL-10-deficient mice make larger granulomas during the acute period of disease (Wynn et al., 1998). In contrast, TNF-α deficient mice are unable to form granulomas and the treatment of such mice with TNF-α, restore the capacity of such mice to form discrete granulomas around parasite eggs (Amiri et al., 1992). Moreover, treatment of immunocompetent mice with anti-TNF-α serum resulted in reduced granuloma size (Joseph and Boros, 1993). So, VIP could reduce the granuloma size through up-regulation of IL-10 and down-modulation of TNF-α production (Allam, 2007).
With regards to the enzymatic changes in the liver, the current data indicated that liver ALT, AST and γ-GT activities were remarkably depleted in the S. mansoni-infected diabetic mice, while LDH activity was increased. Concomitant with these results, Awadalla et al. (1975), Allam and Ahmed (2005) and Allam (2007) noticed that liver ALT, AST and γ-GT activities were profoundly reduced in the S. mansoni-infected mice. In addition, Mansour et al. (1982) and Mahmoud et al. (2002) found that the level of these enzymes was elevated in serum of the parasite-infected mice. However, Abdel Reheim et al. (2007) and Ahmed (2009) revealed that liver ALT, AST and LDH activities were increased in the diabetic rats and mice due to greater need to the gluconeogenic substrates in such condition. Thus, based on these findings, it can be suggested that the depletion of liver activities of ALT, AST and γ-GT in the present study may be attributed to the increased leakage of these enzymes from necrotic tissue and the replacement of normal liver tissue by granulomatous lesions which develop around the entrapped schistosome eggs as well as due to the increased permeability as a result of anoxia and irritation by toxic metabolic products of worms in infected diabetic mice (Ahmed and Mostafa, 2003; Allam, 2007). Overall, in our opinion, the changes of these liver enzyme activities in the ID mice may be the net result of the effects of STZ diabetes and schistosomiasis together. The administration of VIP to the ID mice, in the current study, induced a detectable amelioration of these altered liver enzyme activities. This alleviation could be attributed to a reduction in the granuloma size and number as well as the decrease in the damage of hepatocytes in the infected diabetic treated group as compared to infected diabetic control. VIP may protect hepatocytes from the destructive inflammatory response induced by schistosome eggs via inhibition of pro-inflammatory, as well as, stimulation of anti-inflammatory cytokines. The data of the present study support this suggestion since it indicates that the elevated pro-inflammatory cytokine TNF-α level in serum was significantly decreased while the anti-inflammatory Th2 cytokine IL-10 was potentially increased in the infected diabetic mice administered VIP as compared to the infected diabetic control. The decrease in the size and number of granulomas in the liver as a result of VIP treatment of infected diabetic mice was associated with nearly normalization of the elevated liver collagen content. This indicates that VIP may protect the liver of infected diabetic mice from fibrosis and cirrhosis. VIP may reduce collagen deposition by direct effect on hepatic stellate cells similar to somatostatin, which has been shown to modulate collagen I and III synthesis in activated hepatic stellate cells during schistosomiasis (Chatterjee et al., 2004; Allam, 2007). It was also reported by Kmiec (2001) that VIP may attenuate the conversion of quiescent stellate cells into moyfibroblast-like cells that play a key role in the development of inflammatory fibrotic response.
Tumor markers CA 19.9 and AFP levels were detectably increased in the ID mice and were decreased as a result of VIP administration to values below normal levels. These results are in accordance with those of Allam (2007). Elevated serum CA 19.9 level is associated with gastrointestinal cancer, colorectal cancer and gastric ulceration (Fouad and Khalaf, 1994; Chan and Sell, 1996), while elevated AFP level is associated with hepatocellular carcinoma, massive hepatic necrosis, hepatitis and liver cirrhosis (Noeman et al., 1994; Abelev, 2001). The amelioration of these parameters after VIP administration may be secondary to the improvement effect on liver and intestinal eggs count and the decrease in liver inflammation, necrosis and granuloma size as indicated in this current study.
Schistosoma mansoni infection not only triggers the production of reactive oxygen species in the mouse liver but also leads to an alteration in antioxidant defenses (Abdallahi et al., 1999; Pascal et al., 2000; La Flamme et al., 2001). This finding raises the possibility that oxidative stress may be a contributor of S. mansoni-associated pathology (Pascal et al., 2000). On the other hand, considerable clinical and experimental evidence now exists suggesting the involvement of free radicals-mediated oxidative processes in the pathogenesis of diabetes mellitus as well as its complications (Ahmed, 2003; Jain et al., 2007). These findings agree well with the results of the present study that revealed a profound depletion of the liver total thiol, glutathione, catalase levels while lipid peroxidation and peroxidase activity were elevated in the infected diabetic mice as compared to normal. The free-radical formation was evoked by the pro-inflammatory and inflammatory cytokines (La Flamme et al., 2001; Tabatabaie et al., 2003) such as IL-1β and TNF-α which was significantly increased in the infected diabetic mice of our study. Also, Reactive Oxygen Species (ROS) are considered to be a strong stimulus for the release of cytokines (Vassilakopoulos et al., 2003). The heme peroxidase detected in the liver of naive mice of the present study may be attributed to its activity in Kupffer cells and endothelial cells lining the hepatic sinusoids (Stohr et al., 1978; Pino and Bankston, 1979; Brown et al., 2001; Tafazoli and O’Brien, 2005). Nonetheless, this enzyme is not found in hepatocytes as indicated by Abdallahi et al. (1999). It is also relevant to mention that the peroxidase, assayed in this study using pyrogallol as a substrate, in normal and infected mice, may be myeloperoxidase and eosinophil peroxidase (heme peroxidases) rather than glutathione peroxidase which has selenium in its active site, is highly specific for glutathione and can not oxidize any other substrate (Shigeoka et al., 1991). The marked increase of peroxidase activity in the liver of S. mansoni-infected mice may be the result of the release of eosinophil peroxidase from eosinophil granulocytes and myeloperoxidase from mononuclear phagocytes in the region of granulomas around parasite eggs in addition to the stimulation of Kupffer cells (Weiss et al., 1983; Anderson et al., 1997; Abdallahi et al., 1999; Gharib et al., 1999). The myeloperoxidase and eosinophil peroxidase oxidize halides (I–, Br–, Cl–) using H2O2 generating hypohalous acids. This reaction leads to the production of long-lived oxidant species (Weiss et al., 1983; Kazura et al., 1985; Anderson et al., 1997; Abdallahi et al., 1999) in vicinity of the parasite eggs. The administration of VIP to the infected diabetic mice, in the present study, induced a remarkable amelioration of the tested liver oxidative stress markers and antioxidant defense markers. This alleviation in oxidative stress markers in VIP-treated infected diabetic mice may be secondary to the improvement in the hyperglycemic and metabolic states, at one hand and a decrease in the inflammatory cytokine TNF-α as well as an increase of anti-inflammatory cytokines as IL-10, on the other hand. In accordance with this suggestion, Larocca et al. (2007) stated that VIP inhibits nitric oxide through an increase of IL-10 level. Also, Ahmed (2009) found that VIP improved oxidative stress concomitant with both improvement in the glycemic state and great increase of serum IL-10.
In conclusion, the VIP may have anti-hyperglycemic and anti-schistosomal effects in Schistosoma mansoni-infected diabetic albino mice. These ameliorative effects may be mediated via insulinotropic, immunomodulatory (by increasing anti-inflammatory and decreasing pro-inflammatory cytokines) and antioxidant effects of VIP. However, further studies are required to assess the efficacy and safety of VIP in S. mansoni-infected diabetic human beings.
REFERENCES
- Abad, C., R.P. Gomariz and J. Wascheck, 2006. Neuropeptide mimetics and antagonist in the treatment of inflammatory diseases. Curr. Top. Med. Chem., 6: 151-163.
PubMed - Abdallahi, O.M.S., S. Hanna, M. de Reggi and B. Gharib, 1999. Visualization of oxygen radical production in mouse liver in response to infection with Schistosoma mansoni. Liver, 19: 495-500.
CrossRefDirect Link - Adeghate, E., A.S. Ponery, A.K. Sharma, T. Sharkawy and T. Donath, 2001. Diabetes mellitus is associated with a decrease in vasoactive intestinal polypeptide content of gastrointestinal tract of rat. Arch. Physiol. Biochem., 109: 246-251.
PubMed - Ahmed, O.M. and O.M.S. Mostafa, 2003. Experimental use of black seed oil against Schistosoma mansoni in albino mice. IV. Oxidative stress markers and some biochemical parameters in liver and kidney. J. Egypt. Ger. Soc. Zool., 41A: 227-254.
Direct Link - Allam, G., 2007. Vasoactive intestinal peptide inhibits liver pathology in acute murine Schistosomiasis mansoni and modulates IL-10, IL-12 and TNF-α production. Immunobiology, 212: 603-612.
PubMed - Amirshahrokhi, K., A.R. Dehpour, J. Hadjati, M. Sotoudeh and M. Ghazi-Khansari, 2008. Methadone ameliorates multiple-low-dose streptozotocin-induced diabetes in mice. Toxicol. Applied Pharmacol., 232: 119-124.
CrossRef - Anderson, M.M., S.L. Hazen, F.F. Hsu and J.W. Heinecke, 1997. Human neutrophils employ the myeloperoxidase-hydrogen peroxide-chloride system to convert hydroxy-amino acids into glycolaldehyde, 2-hydroxypropanal and acrolein. J. Clin. Invest., 99: 424-432.
PubMed - Araujo, M.I., P.S. Hoppe, M. Medeiros Jr. and E.M. Carvahlo, 2004. Schistosoma mansoni infection modulates the immune response against allergic and auto-immune diseases. Mem. Inst. Oswaldo Cruz, Rio de Janeiro, 99: 27-32.
CrossRef - Baykal-Erkilic, A., E. Erkilic, M. Erkilic and T. Ozden, 1995. The effect of streptozotocin-induced diabetes on renal plasma flow. Exp. Clin. Endocrinol., 103: 367-372.
Direct Link - Bergmeyer, H.U., P. Scheibe and A.W. Wahlefeld, 1978. Optimization of methods for aspartate aminotransferase and alanine aminotransferase. Clin. Chem., 24: 58-73.
PubMedDirect Link - Beutler, E., O. Duron and B.M. Kelly, 1963. Improved method for the determination of blood glutathione. J. Lab. Clin. Med., 61: 882-888.
PubMedDirect Link - Bonser, A. and P. Garcia-Webb, 1984. C-peptide measurement: Methods and clinical utility. Crit. Rev. Clin. Lab. Sci., 19: 297-352.
PubMed - Brown, K.E., E.M. Brunt and J.W. Heinecke, 2001. Immunohistochemical detection of myeloperoxidase and its oxidation products in Kupffer cells of human liver. Am. J. Pathol., 159: 2081-2088.
PubMed - Buhl, S.N. and K.Y. Jackson, 1978. Optimal conditions and comparison of lactate dehydrogenase catalysis of the lactate to pyruvate and pyruvate to lactate in human serum at 25, 30 and 37 oC. Clin. Chem., 24: 828-831.
Direct Link - Butler, A.E., J. Janson, S. Bonner-Weir, R. Ritzel, R.A. Rizza and P.C. Butler, 2003. Beta-cell deficit and increased beta-cell in humans with type 2 diabetes. Diabetes, 52: 102-110.
PubMed - Caldas, I.R., A.C. Campi-Azevedo, L.F.A. Olivera, A.M.S. Silveira, R.C. Olivera and G. Gazzinelli, 2008. Human schistosomiasis mansoni: Immune responses during acute and chronic phases of infection. Acta Trop., 108: 109-117.
CrossRef - Chance, B. and A. Maehley, 1955. Assay of Catalases and Peroxidases: Peroxidase Assay by Spectrophotometeric Measurement of Disappearance of Hydrogen Donor or the Appearance of their Coloured Oxidation Products. In: Methods in Enzymology, Colowick, S. and N. Kaplan (Eds.). Vol. 2, Academic Press, New York, USA., ISBN-13: 9780121818029, pp: 764-775.
- Chatterjee, S., A. Mbaye, A.T. Alfidja, J. Weyler and J.T. Scott et al., 2004. Circulating levels of the neuropeptide hormone somatostatin may determine hepatic fibrosis in Schistosoma mansoni infections. Acta Tropica, 90: 191-203.
CrossRefDirect Link - Cheever, A.W., 1968. Condition affecting the accuracy of potassium hydroxide digestion techniques for counting Schistosoma mansoni eggs in tissues. Bull. World Health Org., 39: 328-331.
PubMed - Cheever, A.W., M.E. Williams, T.A. Wynn, F.D. Finkelman and R.A. Seder et al., 1994. Anti-IL-4 treatment of Schistosoma mansoni-infected mice inhibits development of T cells and non-B, non-T cells expressing Th2 cytokines while decreasing egg-induced hepatic fibrosis. J. Immunol., 153: 753-759.
PubMed - Chorny, A., E. Gonzalez-Rey, A. Fernandez-Martin, D. Pozo, D. Ganea and M. Delgado, 2005. Vasoactive intestinal peptide induces regulatory dendritic cells with therapeutic effects on autoimmune disorders. Proc. Natl. Acad. Sci. USA., 102: 13562-13567.
PubMed - Cnop, M., N. Welsh, J. Jonas, A. Jorns and S. Lenzen, 2005. Mechanisms of pancreatic β-cell death in type 1 and type 2 diabetes. Diabetes, 54: S97-S107.
PubMed - Cohen, G., D. Dembiec and J. Marcus, 1970. Measurement of catalase activity in tissue extracts. Ann. Biochem., 34: 30-38.
CrossRefPubMedDirect Link - Conyers, S.M. and D.A. Kidwell, 1991. Chromogenic substrates for horseradish peroxidase. Anal. Biochem., 192: 207-211.
Direct Link - De Jesus, A.R., A. Silva, L.B. Santana, A. Magalhaes and A.A. de Jesus et al., 2002. Clinical and immunologic evaluation of 31 patients with acute Schistosomiasis mansoni. J. Infect. Dis., 185: 98-105.
PubMedDirect Link - Delgado, M. and D. Ganea, 1999. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide inhibit interleukin-12 transcription by regulating nuclear factor B and Ets activation. J. Biol. Chem., 274: 31930-31940.
Direct Link - Delgado, M., E.J. Munoz-Elias, Y. Kan, I. Gozes, M. Fridkini and D.E. Brenneman et al., 1998. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide inhibit tumor necrosis factor-α transcriptional activation by regulating nuclear factor-kB and cAMP response element-binding protein/c-Jun. J. Biol. Chem., 273: 31427-31436.
PubMed - Delgado, M., E.J. Munoz-Elias, R.P. Gomariz and D. Ganea, 1999. VIP and PACAP inhibit IL-12 production in LPS-stimulated macrophages: Subsequent effect on IFN-γ synthesis by T cells. J. Neuroimmunol., 96: 167-181.
Direct Link - Delgado, M., E.J. Munoz-Elias, R.P. Gomariz and D. Ganea, 1999. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide enhance IL-10 production by murine macrophages: In vitro and In vivo studies. J. Immunol., 162: 1707-1716.
Direct Link - Delgado, M., E.J. Munoz-Elias, R.P. Gomariz and D. Ganea, 1999. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide prevent inducible nitric oxide synthase transcription in macrophages by inhibiting NF-kappa B and IFN regulatory factor 1 activation. J. Immunol., 162: 4685-4696.
PubMedDirect Link - Delgado, M., D. Pozo, C. Martinez, J. Leceta, J. R. Calvo, D. Ganea and R.P. Gomariz, 1999. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide inhibit endotoxin-induced TNF-alpha production by macrophages: In vitro and in vivo studies. J. Immunol., 162: 2358-2367.
Direct Link - Delgado, M., C. Abad, C. Martinez, M.G. Juarranz and A. Arranz et al., 2002. Vasoactive intestinal peptide in the immune system: Potential therapeutic role in inflammatory and autoimmune diseases. J. Mol. Med., 80: 16-24.
PubMed - Delgado, M., D. Pozo and D. Ganea, 2004. The significance of vasoactive intestinal peptide in immunomodulation. Pharmacol. Rev., 56: 249-290.
PubMed - Delgado, M., A. Chorny, E. Gonzalez-Rey and D. Ganea, 2005. Vasoactive intestinal peptide generates CD4+CD25+ regulatory T cells in vivo. J. Leukoc. Biol., 78: 1327-1338.
PubMed - Duvall, R.H. and W.B. DeWitt, 1967. An improved perfusion technique for recovering adult schistosomes from laboratory animals. Am. J. Trop. Med. Hyg., 16: 483-486.
CrossRefDirect Link - El-Wakil, H.S., T.S. Aboushousha, O. El Haddad, N.B. Gamil, T. Mansou and H. El-Said, 2002. Effect of Schistosoma mansoni egg deposition on multiple low doses streptozotocin-induced insulin dependent diabetes mellitus. J. Egypt. Soc. Parasitol., 32: 987-1002.
PubMed - Engels, D., L. Chitsulo, A. Montresor and L. Savioli, 2002. The global epidemiological situation of schistosomiasis and new approaches to control and research. Acta Tropica, 82: 139-146.
CrossRef - Frebourg, T., E. Bercoff, N. Manchon, J. Senant and J.P. Basuyau et al., 1988. The evaluation of CA 19-9 antigen level in the early detection of pancreatic cancer. A prospective study of 866 patients. Cancer, 62: 2287-2290.
Direct Link - Gomariz, R., J. Leceta, C. Martinez, C. Abad, D. Ganea and M. Delgado, 2000. Anti-inflammatory actions of VIP/PACAP. Role in endotoxemia. Ann. N. Y. Acad. Sci., 921: 284-288.
PubMed - Gomariz, R.P., C. Martinez, C. Abad, J. Leceta and M. Delgado, 2001. Immunology of VIP: A review and therapeutical perspectives. Curr. Pharm. Des., 7: 89-111.
PubMed - Gomariz, R.P., Y. Juarranz, C. Abad, A. Arranz, J. Leceta and C. Martinez, 2006. VIP-PACAP system: New insights for multitarget therapy. Ann. N. Y. Acad. Sci., 1070: 51-74.
PubMed - Gomariz, R.P., A. Arranz, Y. Juarranz, I. Gutierrez-Can, M. Garcia-Gomez, J. Leceta and C. Martinez, 2007. Regulation of TLR expression, a new perspective for the role of VIP in immunity. Peptides, 28: 1825-1832.
PubMed - Gonzalez-Rey, E., A. Chorny and M. Delgado, 2007. Regulation of immune tolerance by anti-inflammatory neuropeptides. Nat. Rev., 7: 52-63.
PubMed - Hulstijn, M., R.M. de Oliveira, E.G. de Moura and G.R. Machado-Silva, 2001. Lower faecal egg excretion in chemically-induced diabetic mice infected with Schistosoma mansoni due to impaired egg maturation. Mem. Inst. Oswaldo Cruz, 96: 393-396.
CrossRef - Jain, S.K., J.L. Rains and J.L. Croad, 2007. Effect of chromium niacinate and chromium picolinate supplementation on lipid peroxidation, TNF-α, IL-6, CRP, glycated hemoglobin, triglycerides and cholesterol levels in blood of streptozotocin-treated diabetic rats. Free Radical Biol. Med., 43: 1124-1131.
CrossRef - Joseph, A.L. and D.L. Boros, 1993. Tumor necrosis factor plays a role in Schistosoma mansoni egg-induced granulomatous inflammation. J. Immunol., 151: 5461-5471.
Direct Link - Joseph, P., K.R. Murthy, J.L. Nelson and A.P. Kulkrani, 1992. Peroxidase: A novel pathway for chemical oxidation in human term placenta. Placenta, 13: 545-554.
Direct Link - Kaneto, H., J. Fujii, H.G. Seo, K. Suzuki and T. Matsoukaet al., 1995. Apoptotic death triggered by nitric oxide in pancreatic beta cells. Diabetes, 44: 733-738.
Direct Link - Kar, M. and D. Mishra, 1976. Catalase, peroxidase and polyphenoloxidase activities during rice leaf senescence. Plant Physiol., 57: 315-319.
CrossRefDirect Link - Karlsson, S. and B. Ahren, 1990. Peptide receptor antagonists in the study of insulin and glucagons secretion in mice. Eur. J. Pharmacol., 191: 457-464.
Direct Link - Kazura, J.W., P. de Brito, J. Rabbege and M. Aikawa, 1985. Role of granulocyte oxygen products in damage of Schistosoma mansoni eggs in vitro. J. Clin. Invest., 75: 1297-1307.
CrossRefDirect Link - Kmiec, Z., 2001. Cooperation of liver cells in health and disease. Adv. Anat. Embryol. Cell. Biol., 161: 1-151.
PubMed - Koster, J.F., P. Biermond and A.J.G. Swaak, 1986. Intracellular and extracellular sulph-hydryl levels in rheumatoid arthritis. Ann. Rheum. Dis., 45: 44-46.
Direct Link - Kubisch, H.M., J. Wang, T.M. Bray and J.P. Philips, 1997. Targeted overexpression of Cu/Zn superoxide dismutase protects pancreatic beta cells against oxidative stress. Diabetes, 46: 1563-1566.
Direct Link - La Flamme, A.C., E.A. Patton, B. Bauman and E.J. Pearce, 2001. IL-4 plays a crucial role in regulating oxidative damage in liver during Schistosomiasis. J. Immunol., 166: 1903-1911.
PubMed - Larocca, L., M. Calafat, V. Roca, A.M. Franchi and C.P. Leiros, 2007. VIP limits LPS-induced nitric oxide production through IL-10 in NOD mice macrophages. Int. Immunopharmacol., 7: 1343-1349.
CrossRef - Lichtenberg, F.V., 1962. The host response to eggs of S. mansoni. I. Granuloma formation in the unsensitized laboratory mouse. Am. J. Pathol., 41: 711-731.
Direct Link - Lukic, M.L., S. Stosic-Grujcic and A. Shahin, 1998. Effector mechanisms in low-dose streptozotocin-induced diabetes. Dev. Immunol., 6: 119-128.
Direct Link - Mahmoud, M.R., H.S. El-Abhar and S. Saleh, 2002. The effect of Nigella sativa oil against the liver damage induced by Schistosoma mansoni infection in mice. J. Ethnopharmacol., 79: 1-11.
CrossRefDirect Link - Mandrup-Poulsen, T., 1996. The role of interleukin-1 in the pathogenesis of IDDM. Diabetologia, 39: 1005-1029.
Direct Link - Marschner, I., P. Bottermann, F. Erhardt, R. Linke and V. Maier et al., 1974. Group experiments on the radioimmunological insulin determination. Horm. Metab. Res., 6: 293-296.
CrossRefDirect Link - Mellado-Gil, J.M. and M. Aguilar-Diosdado, 2004. High glucose potentiates cytokine- and streptozotocin-induced apoptosis of rat islet cells: Effect on apoptosis related genes. J. Endocrinol., 183: 155-162.
PubMed - Metwali, A., A. Blum, D.E. Elliott and J.V. Weinstock, 2002. IL-4 inhibits vasoactive intestinal peptide production by macrophages. Am. J. Physiol. Gastrointest. Liver Physiol., 283: G115-G121.
PubMed - Miyamoto, A., M. Takeshita, H. Pan-Hou and H. Fujimori, 2008. Hepatic changes in adenine nucleotide levels and adenosine 3'-monophosphate forming enzyme in streptozotocin-induced diabetic mice. J. Toxicol. Sci., 33: 209-217.
PubMed - Neuman, R.E. and M.A. Logan, 1950. The determination of collagen and elastin in tissues. J. Biol. Chem., 186: 549-556.
Direct Link - Neuman, R.E. and M.A. Logan, 1950. The determination of hydroxyproline. J. Biol. Chem., 184: 299-306.
PubMedDirect Link - Nicoletti, F., G. Mancuso, F.A. Ciliberti, C. Beninatti, M. Carbone and S. Franco, 1997. Endotoxin-induced lethality in neonatal mice is counteracted by interleukin-10 (IL-10) and exacerbated by anti-IL-10. Clin. Diagn. Lab. Immunol., 4: 607-610.
Direct Link - Nielsen, K., A.E. Karlsen, M. Deckert, O.D. Madsen, P. Serup and T. Mandrup-Poulsen, 1999. Β-Cell maturation leads to in vitro sensitivity to cytotoxins. Diabetes, 48: 2324-2332.
Direct Link - Pascal, M., O.M. Abdallahi, N.E. Elwali, A. Mergani, M.A. Qurashi, M. Magzoub, M. De Reggi and B. Gharib, 2000. Hyaluronate levels and markers of oxidative stress in serum of Sudanese subjects at risk of infection with Schistosoma mansoni. Trans. R. Soc. Trop. Med. Hyg., 94: 66-70.
PubMed - Pehuet-Figoni, M., E. Ballot, J.E. Bach and L. Chatennoud, 1992. The effects of interleukin-1 on pancreatic beta cell function in vitro depend on the glucose concentration. Eur. Cytokine Networks, 3: 443-450.
Direct Link - Preuss, H.G., S.T. Jarrel, R. Scheckenbach, S. Lieberman and R.A. Anderson, 1998. Comparative effects of chromium, vanadium and Gymnema sylvestre on sugar-induced blood pressure elevations in SHR. J. Am. Coll. Nutr., 17: 116-123.
CrossRefPubMedDirect Link - Pusztai, P., J. Prechl, A. Somogyi, E. Szaleczky and J. Feher, 1996. Experimental models in research of the pathomechanism of diabetes mellitus. Orv. Hetil., 137: 1865-1869.
Direct Link - Rabinovitch, A., 1998. An update on cytokines in the pathogenesis of insulin-dependent diabetes mellitus. Diabetes Metab. Rev., 14: 129-151.
Direct Link - Reinke, E. and Z. Fabry, 2006. Breaking or making immunological privilege in the central nervous system: The regulation of immunity by neuropeptides. Immunol. Lett., 104: 102-109.
CrossRef - Tabatabaie, T., A. Vasquez-Weldon, D.R. Moore and Y. Kotake, 2003. Free radicals and the pathogenesis of type 1 diabetes. β-cell cytokine-mediated free radical generation via cyclooxygenase-2. Diabetes, 52: 1994-1999.
PubMed - Tafazoli, S. and P.J. O'Brien, 2005. Peroxidases: Role in the metabolism and side effects of drugs. Drug Discov. Today, 10: 617-625.
Direct Link - Vassilakopoulos, T., M.H. Karatza, P. Katsaounou, A. Kollintza, S. Zakynthinos and C. Roussos, 2003. Antioxidants attenuate the plasma cytokine response to exercise in humans. J. Applied Physiol., 94: 1025-1032.
PubMed - Waldmann, T.A. and K.R. Mclntire, 1974. The use of radioimmunoassay for alpha-fetoprotein in the diagnosis of malignancy. Cancer, 34: 1510-1515.
Direct Link - Wang, H.Y., X. Jiang, I. Gozes, M. Fridkin, D.E. Brenneman and D. Ganea, 1999. Vasoactive intestinal peptide inhibits cytokine production in T lymphocytes through cAMP-dependent and cAMP-independent mechanisms. Regul. Pept., 84: 55-67.
Direct Link - Weinstock, J.V., A.M. Blum and S. Khetarpal, 1991. Granulomas in murine schistosomiasis mansoni contain vasoactive intestinal peptide-responsive lymphocytes. Cell Immunol., 134: 458-472.
Direct Link - Weiss, S.J., M.B. Lampert and S.T. Test, 1983. Long-lived oxidants generated by human neutrophils characterization and bioactivity. Science, 222: 625-628.
CrossRefDirect Link - Winzell, M.S. and B. Ahren, 2007. Role of VIP and PACAP in islet function. Peptides, 28: 1805-1813.
CrossRef - Xin, Z. and S. Sriram, 1998. Vasoactive intestinal peptide inhibits IL-12 and nitric oxide production in murine macrophages. J. Neuroimmunol., 89: 206-212.
Direct Link - Young, D.S., 1990. Effects of Drugs on Clinical Laboratory Tests. 3rd Edn., AACC Press, Washington, DC.
Direct Link - Zaccone, P., Z. Fehervarti, F.M. Jones, S. Sidobre, M. Kronenberg, D.W. Dunne and A. Cooke, 2003. Schistosoma mansoni antigens modulate the activity of the innate immune respone and prevent onset of type 1 diabetes. Eur. J. Immunol., 33: 1439-1449.
PubMed