
M. Shakeri
Department of Animal Sciences, College of Agriculture, Tehran University, Tehran, Iran
H. Roshanfekr
Department of Animal Sciences, University of Ramin Agricultural and Natural Resources, Ahvaz, Iran
M. Mamoei
Department of Animal Sciences, University of Ramin Agricultural and Natural Resources, Ahvaz, Iran
Kh. Mirzadeh
Department of Animal Sciences, University of Ramin Agricultural and Natural Resources, Ahvaz, Iran
Asian Journal of Animal and Veterinary Advances
Year: 2008 | Volume: 3 | Issue: 6 | Page No.: 400-408







ABSTRACT
The present research studied the effect of the interval between animal`s death and sperm recovery (0, 24 or 48h) on the quality and freezability of ram spermatozoa from cauda epididymidis. Storage temperature of epididymes (room temperature or 5°C) was also analyzed. Spermatozoa were diluted with Tes-Tris-Fructose solution supplemented with egg yolk (10%) and glycerol (4%) and frozen using a programmable bio-freezer (-20°C min-1). Pre-freeze and post-thaw sperm samples showed viable spermatozoa up to 48h after the animal`s death, although their quality declined significantly as post-mortem storage time increased. Epididymes sperm stored at 5°C showed better motility and a lower percentage of abnormal forms than epididymes stored at room temperature after 24 and 48h. The fertilizing ability of cauda epididymis ram spermatozoa obtained at 0 and 24h after the animal`s death is similar to that of ejaculated spermatozoa. Therefore, a good protocol for post-mortem semen collection in rams when epididymal spermatozoa cannot be collected immediately is to preserve the epididymes at 5°C and process the samples in the first 24h after the animal`s death.
PDF Abstract XML References Citation
How to cite this article
M. Shakeri, H. Roshanfekr, M. Mamoei and Kh. Mirzadeh, 2008. Effect of Epididymis Handling Conditions on the Quality of Ram Spermatozoa Recovered Post-Mortem. Asian Journal of Animal and Veterinary Advances, 3: 400-408.
DOI: 10.3923/ajava.2008.400.408
URL: https://scialert.net/abstract/?doi=ajava.2008.400.408
DOI: 10.3923/ajava.2008.400.408
URL: https://scialert.net/abstract/?doi=ajava.2008.400.408
INTRODUCTION
The unexpected death of animals of high genetic value or zoological interest, as well as the difficulty in collecting semen from wild species, is a handicap to the application of assisted reproduction techniques for the preservation of biodiversity. The recovery and freezing of viable sperm from the epididymes of dead animals (post-mortem recovery) is an interesting option for preserving male gametes and thus for maintaining germplasm banks. There are many studies on semen collection from the cauda epididymis of several species (Fournier-Delpech et al., 2001; Lambrechts et al., 2006), but there are no data available on the effects of different storing conditions on ram epididymal spermatozoa. The conditions (time and temperature) under which epididymis is handled could cause important changes in the viability of sperm samples. It should be kept in mind that animals die unexpectedly and far away from the lab where the sperm sample could be properly processed and stored. In breeding rams, Aguado et al. (2005) reported the preservation of ram sperm stored at room temperature for 0, 3, 6, 9, 12 and 24h. These authors found semen of better quality, both before and after freezing, when it was collected in the first 3h after death. Under similar conditions, Garde et al. (2004) found no remarkable variations in the fertilizing ability of ram epididymal samples processed in the first 24h after death, noticing a marked diminution in sperm viability for longer periods of time. In red deer (Cervus elaphus) and moufflon (Ovis musimon), Garde et al. (2005) concluded that viability and in vitro fertility (percentage of penetration in hamster oocytes) of sperm decreased when the time between the animal`s death and the moment of semen collection increased (up to 40h at ambient temperature). They pointed out that there were appreciable differences between species with regard to the hamster oocyte penetration test, where a particularly marked decrease was observed in the case of the red deer.
These studies did not analyze the effect of storage temperature on epididymal semen quality. However, Kikuchi et al. (2004) in pig and Kishikawa et al. (2001) in mice suggested that when valuable male animals die unexpectedly and sperm cryopreservation is not possible immediately, temporal storage of epididymides at 40°C may help to preserve the genome of individuals. In order to establish a model for post-mortem sperm recovery in ram, we studied the effect of the interval between death and sperm collection (0, 24 or 48h) as well as the storage temperature of epididymes (room temperature or 5°C) on the quality and fertilizing ability (evaluated by in vitro fertilization) of pre-freezing and post-thawed sperm from the cauda epididymidis.
MATERIALS AND METHODS
Experimental Design
This experiment was conducted in spring, 2007 at Ziaran Slaughter House and Physiology Laboratory of Animal Science Department, College of Agriculture, Karaj, Tehran University in Iran. Testicles from 50 Mehraban breed rams were collected at an abattoir. One testicle from each pair was transported at room temperature (22°C) and semen collection was carried out in the first 2h after the slaughter of the ram (control group, CG). The other testicle was transported either at room temperature (22°C, group R) or at 5°C (group C). Testicles were stored prior to sperm recovery for 24h (groups R24 and C24) or 48h (groups R48 and C48).
Collection and Quality Evaluation of Sperm
After isolation, the epididymis-testicle complexes were dissected into three parts: testicle, epididymis and cauda epididymis. Sperm was obtained by slicing the tissue of the cauda epididymis with a scalpel; the fluid was collected and its volume was estimated. To limit contamination, epididymis samples were carefully dissected free of blood clots and extraneous tissues. Care was taken not to cut blood vessels. Sperm concentration was determined using a haemocytometer. The percentage of Total Motility (TM) and Progressive Motility (PM) was determined microscopically, observing eight random fields in a flat drop (4μL) of semen diluted in freezing extender (200x phase contrast; heating plate at 37.5°C). Acrosome integrity was determined using 5μL of a semen sample fixed in 0.5mL of glutaraldehyde fixative solution (GS glutaraldehyde at 2% in 100mL aqueous solution with 2.9g glucose monohydrate, 1g sodium citrate tribasic dihydrate and 0.2g sodium bicarbonate; Sigma, USA). A flat drop of fixed sample was placed on a microscope slide. For each group, two slides were observed by phase-contrast microscopy (600x), 200 spermatozoa were counted for each slide and percentage of spermatozoa with normal acrosome was noted.
The functional integrity of the sperm plasma membrane was evaluated using the hypo osmotic swelling test (HOS test): 5μL of semen were diluted in 0.5μL of 100mOsm kg-1 aqueous sodium citrate solution; after 18min of incubation (at room temperature) samples were fixed with a drop of GS solution. Response to the test was quantified (percentage of spermatozoa swollen = percentage of positive endosmosis or % E+) with a phase-contrast microscope (400x).
Sperm morphology (percentage of Cytoplasmic Droplets (CD) and percentage of Total Abnormal Spermatozoa (TAS)) were determined from a sample of 5μL of semen fixed in 0.5mL of GS solution, using phase-contrast microscopy (600x). Sperm abnormalities were grouped according to the location of the morphological anomaly (head, middle piece or flagellum) using criteria of sperm morphology established by WHO (2004). All the above mentioned parameters were calculated for pre-freezing and post-thawed semen. Recovery Rates (RR) for total motility, progressive motility, percentage of normal acrosomes and percentage of positive endosmosis were calculated, using the following formula:
Freezing and Thawing of Sperm
Ten minutes after collection, semen was diluted 1:1 with Tes-Tris-Fructose extender (Sigma) containing 10% egg yolk and 4% glycerol. The diluted sample was chilled to 5°C (at a rate of -0.2°C min-1) and further dilution with the same diluent`s was carried out to yield a final concentration of 200x106spermatozoa mL-1. Diluted sperm was allowed to equilibrate for 2h and was loaded into 0.25mL straws. Finally, it was frozen in a programmable bio-freezer (Planner MRIII®) at a rate of -20°C min-1 to -100°C. Frozen samples were stored in liquid nitrogen until further use. Thawing was carried out in a water bath at 65°C for 6sec.
In vitro Fertilization
Briefly, visible follicles (3-7mm in diameter) of slaughterhouse ovaries were aspirated into TCM-19 medium (Sigma) and complemented with gentamycine (0.4%; Sigma) and heparin (5 UI mL-1; Sigma). Only oocytes with homogenous cytoplasm and a complete and compact cumulus cell investment were used. Oocyte maturation took place in TCM-199 medium supplemented with ovine 5μg mL-1 FSH (Ovagen, ICP), sheep follicular fluid (10%) and gentamycine (0.4%), for 24h at 38.5°C in a controlled atmosphere (5% CO2) and humidity at saturation level. Six hundred selected oocytes were divided into six groups (four sets of 25 oocytes for each experimental group) for the IVF. These groups were used to compare the fertility of the five types of frozen epididymal semen corresponding to the experimental design and a control of frozen ejaculated ovine semen. Thawed sperm was rinsed and selected on a Percoll gradient (Percoll 45/90; Sigma) and the highly motile fraction was selected for IVF at a concentration of 106cells mL-1. Fertilization was carried out in a modified Brackett`s defined medium (DM-H) buffered with 10mM HEPES (Huneau and Crozet, 2001) and containing 20% (v/v) estrus sheep serum for 18h at 38.5°C in a controlled atmosphere (5% CO2) and humidity at saturation level. Embryos were then cultured in modified synthetic oviduct fluid (mSOF) according to Takahashi and First (2000) for 24h at 38.5°C (5% CO2, 5% O2 and 90% N2) and then the rate of embryo cleavage was determined.
Statistical Analysis
Sperm quality data were evaluated using General Linear Model (GLM) procedures of the SAS™ program to analyze variance components. Factors of variation were the type of post-mortem storage of epididymes and spermatozoa status (pre-freeze or post-thaw). Comparison of means was performed with Duncan`s test (p<0.05). The rates of embryo cleavage were compared using the Chi-square test.
RESULTS
Table 1 shows the mean values of general parameters for the 100 testicles and epididymes processed in this study.
| Table 1: | General parameters of epididymal sperm recovery in this study |
 | |
| Table 2: | Effect of post-mortem storage on epididymal rams spermatozoa motility (Mean ± SEM) |
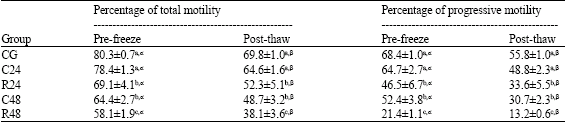 | |
| Different superscripts (a, b, c) indicate a difference (p<0.05) among groups. Different superscripts (α, β) indicate a difference (p<0.05) among pre-freeze and post-thaw spermatozoa | |
| Table 3: | Effect of post-mortem storage on acrosomal status and HOS-test reactivity of epididymal ram spermatozoa (Mean ± SEM) |
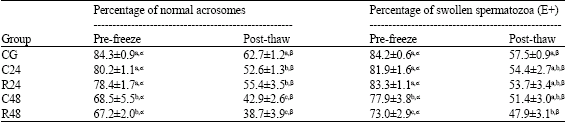 | |
| Different superscripts (a, b, c) indicate a difference (p<0.05) among groups. Different superscripts (α, β) indicate a difference (p<0.05) among pre-freeze and post-thaw spermatozoa | |
Motility
No significant decrease was observed after 24h at 5°C in comparison to the control values. Pre-freeze motility shows a significant decrease after 48h (at both 5°C and room temperature) and 24h at room temperature with regard to control and C24 groups (Table 2). The pre-freezing values of total and progressive motility of all experimental groups decrease significantly (p<0.05) after thawing.
Acrosomal Status and Cell Membrane Integrity
Epididymal spermatozoa stored for 24h (C24 and R24) show no significant differences in pre-frozen percentages of normal acrosomes (NA) in comparison with the control group (Table 3). In contrast, this percentage decreases significantly (p<0.05) after 48h of storage (C48 and R48). However, NA percentages of post-thaw samples show significant differences (p<0.05) amongst the three groups defined by storage time (0, 24 and 48h). In these groups, acrosome status of post-thaw samples is not affected by the temperature at which the epididymes are stored.
Before freezing, the percentages of swollen spermatozoa in 48h groups (R48 and C48) are significantly different (p<0.05) to those in the control and 24h groups (R24 and C24). After thawing, these differences (p<0.05) are only detected between the control group and R48.
All groups show a significant decrease in the percentage of spermatozoa with normal acrosome after thawing. This effect of the freezing procedure is also observed in the percentage of swollen spermatozoa.
Abnormal Spermatozoa and Cytoplasmic Droplets
In pre-freezing sample, epididymes stored at room temperature show the percentage of total abnormal spermatozoa than the control group (Table 4). This increase is not observed when epididymides are stored at 5°C. After thawing, the TAS percentage shows significant differences only between group R48 and all other groups. The pre-frozen percentage of spermatozoa with CD is not affected by post-mortem handling of epididymes (Table 5). In post-thaw samples, this percentage decreases significantly in group R48 in comparison with groups C24 and R24.
Post-Thaw Recovery Rate
Analysis of the recovery rates for total motility and progressive motility after storage (Table 6) shows that the freezing procedure produces a significant reduction in the 48h storage groups (C48 and R48). Rates of normal acrosomal spermatozoa are affected by freezing at both 24 and 48h but sample R24 does not differ from the control group. Recovery rates of E+ do not change significantly amongst groups (Table 6).
Different superscripts (α, β) indicate a difference (p<0.05) among pre-freeze and morphological defects in the flagellum are the most frequent abnormality in pre-frozen samples (64.5%, Fig. 1) and these undergo a significant increase in post-thaw samples (74.8%).
In vitro Fertility
In IVF experiments, cleaved oocytes using epididymal sperm from groups CG, R24 and C24 do not differ significantly from values observed with post-thaw ejaculated ram semen (Fig. 2). When epididymal semen is recovered 48h after the animal`s death, the embryo cleavage rate reduces significantly (p<0.05) for both storage temperatures (C48 and R48) with regard to the other groups.
| Table 4: | Effect of post-mortem storage on percentage of abnormalities in epididymal rams spermatozoa (Mean ± SEM) |
 | |
| Different superscripts (a, b, c) indicate a difference (p<0.05) among groups. Different superscripts (α, β) indicate a difference (p<0.05) among pre-freeze and post-thaw spermatozoa | |
| Table 5: | Effect of post-mortem storage on percentage of cytoplasmic droplets in epididymal rams spermatozoa |
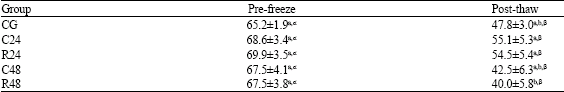 | |
| Different superscripts (a, b) indicate a difference (p<0.05) among groups. Different superscripts (α, β) indicate a difference (p<0.05) among pre-freeze and post-thaw spermatozoa | |
| Table 6: | Post-thaw recovery rate (%) of qualitative parameters in epididymal rams spermatozoa according to storage method (Mean ± SEM) |
 | |
| Different superscripts (a, b, c) indicate a difference (p<0.05) among groups in each parameter | |
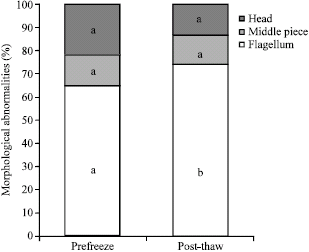 | |
| Fig. 1: | Comparison of pre-freeze and post-thaw proportion of different kind of abnormalities on ram epididymal spermatozoa. Different letters (a, b) indicate differences among columns (p<0.05) |
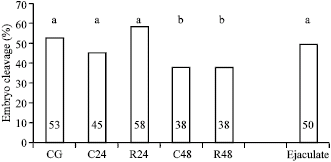 | |
| Fig. 2: | Percentage of cleavage (24h) by different storage types of frozen-thawed epididymal rams spermatozoa. Different letters (a, b) indicate differences among groups (p<0.05) |
DISCUSSION
The qualitative parameters of ram epididymal spermatozoa observed in the control group (80.3% TM, 84.3% NA, 84.2% E+ and 65.2% CD) are similar to those published by Aguado et al. (2005) and Cognié et al. (1999). Total motility of epididymal sperm from breeding rams has been estimated at between 70 and 80% by other authors. According to Amann et al. (2000), in ram cauda epididymidis, about 88% of the sperm had an intact plasma membrane. This result is similar to the one obtained for ovine ejaculate, with 85% of swollen spermatozoa (Nehring, 2003). The percentages of CD described in the present study are similar to those reported by Amann et al. (2000). In contrast, Blash et al. (2001) observed that pre-freeze motility and viability of goat epididymal sperm were higher than those of ejaculated semen.
The quality parameters, total motility, progressive motility and normal spermatozoa, which do not show significant variations during the first 24h post-mortem at 5°C, drop significantly when the epididymis is stored for 48h. Aguado et al. (2005) in ram and Garde et al. (2005) in red deer and moufflon, find that viability of sperm collected from the cauda epididymis decreases progressively as the time between the animal`s death and sperm collection increases.
Present data indicate that refrigeration enables us to obtain better quality semen samples than storage at room temperature. The beneficial effect of refrigeration on various parameters of sperm quality, especially motility, may be explained by the reduced metabolic rate of sperm cells when they are at 5°C (Salamon and Maxwell, 1983). In this sense, Sankai et al. (1968) found that motility of mouse epididymal spermatozoa decreases when the storage temperature is increased, suggesting that this effect is related to changes in spermatozoa metabolic activity. Nevertheless, Kikuchi et al. (2004) concluded that motility of boar spermatozoa collected at 4°C and stored for 1 or 2 days, decreased significantly in comparison with that of control spermatozoa from non-refrigerated epididymis. Also, motility of dog spermatozoa recovered from epididymides stored at 4°C decreased significantly within the first 48h of refrigeration (Yu and Leibo, 1998). These results may evidence that there are differences between species relative to maintenance of spermatozoa viability from post-mortem stored epididymis.
In the present study, post-thaw total motility in both the control group (69.8%) and C24 group (64.6%) is similar to that of ovine ejaculate (70%) mentioned by other author (Garde, 1993). The good freezability of sperm from the cauda epididymidis was also observed by Rath and Niemann (1990) who found 72.2% post-thaw motility in boar spermatozoa. Also, Kikuchi et al. (2004) showed that post-thaw motility of boar spermatozoa from epididymides stored for 1, 2 or 3 days at 4°C does not differ from that of the controls. These authors proposed that although the reason for the survival of spermatozoa in epididymides at 4°C is unclear, epididymal fluid may contain an unknown cold shock protection factor. Nevertheless, the post-thawing/pre-freezing cell motility rate of goat epididymal sperm shows a higher decrease than that of ejaculated semen (Blash et al., 2001). This result may be explained taking into consideration that post-thawing motility appears to depend more on the initial quality of the semen than on the freezing method itself (Fernandes et al., 1990).
The results of this study show that acrosomes of ram epididymal spermatozoa might be sensitive to long storage periods (48h). Our observations also indicate that the percentage of epididymal spermatozoa with intact acrosomes is affected by the freezing procedure. Kikuchi et al. (2004) suggested that pig acrosome integrity may be damaged during cryopreservation and this causes a decrease in sperm fertilizing ability. According to these authors, sperm motility and oocyte penetration ability (reflected by acrosome integrity) are affected by different mechanisms during cold storage of the epididymides. This interpretation may explain the differences between motility parameters (better when preserved at 5°C) and the percentage of normal acrosomes (better when preserved at room temperature) in post-thawing ram spermatozoa analyzed in this study. Yu and Leibo (1998) determined that there is no significant decrease in membrane integrity and acrosome integrity of dog spermatozoa recovered from epididymes and stored at 4°C within the first 48h of refrigeration. The percentage of epididymal spermatozoa with intact acrosomes is very high and shows very little variation within the first day of refrigeration. In the present study, the plasma membrane integrity of post-thawing epididymal spermatozoa does not seem to be much affected by epididymis handling conditions.
The fertilizing ability of epididymal sperm is well known. In ewes inseminated with cauda epididymis spermatozoa, Fournier-Delpech et al. (2001) reported gestation rates of 78-80%; this rate is slightly higher than the one achieved with ovine ejaculates (72%). In this study, no significant difference in IVF was found between epididymal (control and storage for 24h) and ejaculated spermatozoa. In contrast, the fertilizing ability of epididymal spermatozoa stored during 48h decreased significantly. In addition, Garde et al. (2004) found that fertilizing ability evaluated by hamster oocyte in vitro penetration assay using red deer epididymal spermatozoa, gradually decreased as the storage period was prolonged. However, there were no significant differences when using mouflon spermatozoa. These results might reflect a difference amongst several ruminant species.
A reduction in the fertilizing ability of epididymal sperm is described by Kishikawa et al. (2001) when the time between the animal`s death and the recovery of gametes increases (Kikuchi et al., 2004) in boar and in mouse. Such important differences in the fertilizing ability of frozen and thawed sperm from ovine and porcine epididymes might be explained by the different freezing ability of spermatozoa from the two species (Holt, 2000). Ikeda et al. (2002) suggested that the maintenance of acrosomal integrity in unreacted status, rather than the maintenance of sperm motility, is important for in vitro fertilization ability. This hypothesis coincides with the observation that both the cleavage rate and the acrosome status are similar in the R24 and control groups.
In conclusion, frozen-thawed spermatozoa collected from epididymides stored at 5°C for 24h show a fertilizing ability similar to frozen-thawed ejaculated spermatozoa. These data suggest that ram epididymides must be stored at 5°C for 24h when epididymal spermatozoa cannot be immediately collected and cry-preserved. We think that these storage conditions could also be used for epididymal sperm recovery in wild ruminants.
ACKNOWLEDGMENTS
The authors wish to express their thanks to Ziaran Slaughter House and Physiology Laboratory of Animal Science Department, College of Agriculture, Tehran University for their cooperation in the different phases of this research.
REFERENCES
- Fernandes, P.A., J.A. McCoshen, M. Cheang, J.V. Kredentser and A.M. Wodzicki, 1990. Quantitative analysis of the effect of freezing on donor sperm motion kinetics. Fert. Ster., 54: 322-327.
Direct Link - Holt, W.V., 2000. Fundamental aspects of sperm cryobiology: The importance of species and individual differences. Theriogenology, 53: 47-58.
CrossRefPubMedDirect Link