
T. Farahvash
Department of Animal Science, Islamic Azad University-Shabestar Branch, Iran
Sh. Golzar Adabi
Organization of Agricultural Jahad, East Azarbijan, Iran
A. Ahmadzadeh
Department of Herbalogy, Islamic Azad University-Shabestar Branch, Iran
J. Davoodi
Department of Veterinary, Islamic Azad University-Miyaneh Branch, Iran
Asian Journal of Animal and Veterinary Advances
Year: 2008 | Volume: 3 | Issue: 5 | Page No.: 357-362







ABSTRACT
The objective of this retrospective study was to investigate the potential factors affecting sex ratio on dairy herds in East Azarbaijan province of Iran. The data used in this experiment were collected from 186 registered dairy herds are routinely compiled on a monthly basis (from June 1990 to July 2007). For statistical analyses, sex ratios were compared with expected value using goodness-of-fit Chi-square ( χ2) analysis. The effect of some factors such as herd size, artifical insemination year and kind of frozen semen (internal and external ) on sex ratio was determined. Statistical analysis of data shown that herd size had significant effect on sex ratio (χ2 = 5.989, df = 4, p < 0.05). The χ2-test for year variables were significant (χ2 = 52.496, df = 17, p < 0.01), Also the effect of frozen semen (internal and external) on sex ratio was not significant (χ2 = 5.893, df = 7, p > 0.05). Results suggested a positive effect of herd size and year on sex ratio therefor it is possible using this result as a practical and inexpensive method of sex ratio modification.
PDF Abstract XML References Citation
How to cite this article
T. Farahvash, Sh. Golzar Adabi, A. Ahmadzadeh and J. Davoodi, 2008. Some Factors Affecting Sex Ratio of Dairy Herds in East Azarbijan, Iran. Asian Journal of Animal and Veterinary Advances, 3: 357-362.
DOI: 10.3923/ajava.2008.357.362
URL: https://scialert.net/abstract/?doi=ajava.2008.357.362
DOI: 10.3923/ajava.2008.357.362
URL: https://scialert.net/abstract/?doi=ajava.2008.357.362
INTRODUCTION
Sex ratio theory is one of the more developed areas of evolutionary ecology, yet adaptive explanations for sex ratio variation among vertebrates are still the subject of much debate and uncertainty, partly because it can be difficult to infer process from pattern and adaptive and non-adaptive explanations may be confounding (Ian and Hardy, 1997). In animal production systems, the possibility of modify sex ratio can result in a substantial increase of the production in intensive cattle farms. Also, sex ratio manipulation can sensibly enhance the effectiveness of selection and genetic improvement programs, through the differential increment of males or females born after AI (Seidel, 2003).
Gutiérrez-Adán et al. (1999) suggested that the differential ability of X-or Y-bearing spermatozoa to fertilise oocytes depending either on time of insemination or oocyte maturation state, may be due, at least partially, to intrinsic differences in the physiological activity of X-or Y-bearing spermatozoa before fertilisation. Trivers and Willard (1973) hypothesized that in species in which reproductive success varies more among one sex than the other, mothers in better physiological conditions would gain an advantage by investing more heavily in the more variable sex. Similarly, mothers with limited resources would gain an advantage by investing in the more reproductively stable sex, thereby ensuring a continuation of the genetic line. Although many studies have reported support for this hypothesis (Berry and Cromie, 2006; Sheldon and West, 2004; Roche et al., 2006a). Recent results by Roche et al. (2006b) confirmed positive effect of maternal Body Condition Score (BCS) on the Secondary Sex Ratio (SSR) in dairy cows, indicating that despite domestication of the species, they conform to the Trivers-Willard hypothesis. One of the critical parameters for reproduction is the energy availability of the mother (Wade and Schneider, 1992). Because of this, it is possible that variation in maternal energy storage is associated with a modification of the secondary sex ratio.
Fisher`s (1930) theory dictated that maternal investment in male and female offspring is similar and that secondary sex ratio (SSR, the proportion of males to females at birth) should be 50:50 if one sex does not require greater maternal investment than the other. Nevertheless, there is compelling evidence to conclude that, under certain conditions, natural selection favors systematic deviations from this expected 50:50 sex ratio. Furthermore, factors as diverse as latitude of residence, ethnicity, dominant weather patterns, timing and frequency of coitus relative to ovulation, diet; paternal age and alcohol consumption, parental age gap, maternal blood type, BCS, vaginal pH and the systemizing and empathizing skills of the dam have all been statistically associated with altered SSR in mammals (Roche et al., 2006a).
The prediction that females in better body condition would produce more male than female progeny has been observed in red deer, roe deer, mature ewes, reindeer, Barbary sheep, domestic pigs and a number of other species, although there are exceptions (Cheryl et al., 2004). The data on roe deer (Wauters et al., 1995) were obtained with farmed animals on a diet controlled for low-and high-energy intake by varying the oil content. In that study, 75% of the calves born to the high-energy does were male, while the low-energy group produced only 46% males. Some recent studies, however, indicate that, within species, the sex ratio varies with the costs or benefits of producing male or female offspring.
The aim of this retrospective study was investigate the potential some factors affecting sex ratio on dairy herds in East Azarbaijan province of Iran.
MATERIALS AND METHODS
The data used in this experiment were collected from 186 registered dairy herds are routinely compiled on a monthly basis (from June 1990 to July 2007 ) by the Jahad Agricultural Organization (JAO) of East Azarbaijan. Data were recorded by the farm staff on a prepared sheets. The validity and reliability of records were checked regularly by JAO personnel. Although the herds have different management practices, all of them were crossbred or Holsteins and non-seasonally calving dairy herds that were artificially insemination as routine.
The animals were inseminated at different times from the detection of the onset of estrus (between 8 and 44 h). The insemination was carried out using standard methodology for cattle, using frozen/thawed semen, which was deposited in the uterus. Estrus onset was detected and noted down by an experienced person, who carried out continue observations throughout the day, watching behavior and clinical and gynecological symptoms characteristic of the estrus in females of this species (Hafez and Hafez, 2000).
Sex ratio was defined as the ratio of male to female live births multiplied by 100. the expected value for sex ratio was assumed to be equal numbers of males and females. For statistical analyses, sex ratios were compared with expected value using goodness-of-fit Chi-square (χ2) analysis. Also, χ2-test for trend, pearson χ2 and analysis of variance were used. All analysis were done using SPSS soft ware (version 11.5). A p < 0.01 was considered to be statistically significant.
RESULTS AND DISCUSSION
Over the study years, 25812 single live births were recorded, which comprised 13138 male and 12674 female pregnancies. Statistical analyses were done on all regardless of the number of calves, considering that herd size has no significant effect on sex ratio. The frequency of the number of live
| Table 1: | Percent of live births by sex and herd size in dairy cattle in East Azarbijan, Iran |
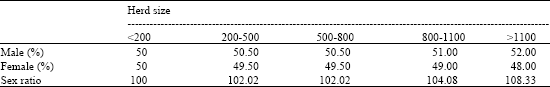 | |
| χ2 = 5.989, df = 4, p < 0.05 | |
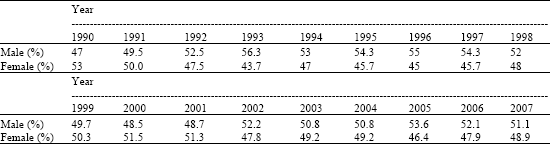
| Table 2: | Percent of male and female by birth year in dairy cattle in East Azarbijan, Iran |
 | |
| Table 3: | Percent of male and female by internal and external sperm in dairy cattle in East Azarbijan, Iran |
births by sex and herd size has been shown in Table 1. Deviation from the expected ratio was significant in herds (p < 0.05). Also, the χ2-test for trend analysis was significant (χ2 = 5.989, df = 4, p < 0.05).
The effect of year variables during the study is summarized in Table 2. The χ2-test for trend analysis were significant (χ2 = 52.496, df = 17, p < 0.01). One of the most important changes during year is climatic factors (p < 0.05, data not show). The odds ratios indicated positive associations between minimum, maximum air temperature and evaporation rate and the likelihood of male calf subsequently being born. Also the effect of kind of frozen semen on sex ratio is not significant (χ2 = 5.893, df = 7, p > 0.05). The effect of internal and external frozen semen on sex ratio are shown in Table 3.
The control of sex ratio in farms conveys many advantages, since it allows to sensibly improve the yield of the operations based on the type of production (milk or meat). At this moment, there are many techniques of proven effectiveness that allow to alter the normal sex ratio, such as flow cytometry stored sperm. However, they are of little practical use, because the cost of preparation of purified samples of X or Y sperm is still excessively high (Martinez et al., 2004).
Furthermore, mares in better condition produced more male foals than expected from an assumption of parity. Where sex ratios differ significantly from parity in both directions, it is unlikely that the difference is due solely to differential loss of less viable or more costly fetuses-usually males in mammalian species (Ansari-Lari, 2006). Alternatively, mechanisms that favor the differential conception of males or females have been postulated (Krackow, 1995).
Hilder et al. (1944) said that there seems to be no indication that the age of the parents has any particular effect on the sex ratio of the offspring.
In Suadi Arabia, Ryan and Boland (1991) reported that the sex of single calves from primiparous cows revealed no significant deviation from the expected ratio. However, the sex ratio was biased towards males offspring in multiparous cows (Foote, 1977). Owing to the small sample size, assessment of sex ratio was not done separately in higher-parity groups.
In present study, the linear trend revealed that the odds of male birthes increases in parallel with herd size (Table 1). This is in agreement with Ansari-Lari (2006) findings. It is apparent that herd size per se cannot for this difference and some other factors closely associated with herd size may be responsible. With respect to factors such as condition of dam and nutritional status, it can be presumed that animals would be kept in better circumstances in large dairies than in smaller ones (Ansari-Lari, 2006). It is reported that dairy cows, but not heifers, on a high plane of nutrition give birth to proportionately more bull than female calves than cows on a poorer diet (Skjervold and James, 1979).
Repeat breeder cows, i.e., ones that have problems becoming pregnant by artificial insemination, also tend to produce more males (King et al., 1985).
In particular, a low pre-pregnancy weight is associated with a deficit of males. The results are consistent with a recent reports showing that in rural Ethiopia, well-fed women were more likely to conceive males (Cagnacci et al., 2004), that the relationship between caloric availability and sex ratio is observed across several countries (Williams and Gloster, 1992) and that there is a higher energy demand for pregnacies with male embryos (Tamimi et al., 2003). These report support the theory of Trivers and Willard (1973), suggesting that a higher proportion of females are produced in non-optimal maternal conditions. The effect of pre-pregnacy weight on sex ratio can be exerted through a selection of gametes as well as through an increase in the rate of male embryo abortions.
There is no obvious reason for the significant effect of year on sex. Of course Roche et al. (2006a) revealed that one contributing factor to the annual effect on sex ratio may be climate and its associated effects on sex ratio.
In agreement with the present study Roche et al. (2006b), using 34 years data on New Zealand dairy cows, reported a significant effect of time of the year at conception on secondary sex ratio. Nonetheless, Foote (1977) failed to report any significant effect of time of the year on sex ratio in dairy cattle while Skjervold and James (1979) reported a significant effect of calving season on sex ratio in parity 1 animals only.
The physiology underpinning an effect of climate on the SSR adjustment is unknown. A number of physiological and endocrine traits can be influenced by environmental factors. Similarly, some psychological disorders have a pronounced annual rhythm (Lerchl, 1998), indicating climatic effects.
Xu et al. (2000) reported a heritability of sex ratio of 2%, suggesting that the permanent environmental effect of the dam is the main contributor to the repeatability of sex ratio. Vandenbergh and Huggett (1995) in an experiment of rodents reported a greater probability of male offspring in dams born between two male siblings. They attributed this effect to a possibly higher level of testosterone in the dam acquired during development in the womb.
Research in humans has revealed a higher sex ratio in mothers with greater testosterone levels (Grant, 1996). Nonetheless, not all studies have documented an effect of previous neonate sex on the sex of the subsequent birth (Berry and Cromie, 2006).
Skjervold and James (1979) reported a significant effect of breed of sire on sex ratio in Norway. Although not statistically tested in the study, Chi-square analysis of the data provided by Foote (1977) identified a significantiy (p < 0.05) lower proportion (50.5%) of males born from Holstein sires than from Jersey sires (52.8%).
Some researchers have studied the variation of the sex ratio depending on the time of the mating relative to ovulation (Seidel, 2003; Rorie, 1999). Numerous efforts have been made to alter the sex of calves by varying time of insemination (Martinez et al., 2004). It has been suggested that early inseminations (i.e., far before ovulation) would result in more female calves whereas late inseminations (i.e., close to ovulation) would result in more male calves, due to different timing of capacitation and survival time of the X-and Y-chromosome bearing spermatozoa in the female reproductive tract (Martinez et al., 2004). However, several other studies offer contradicting explanations for potential effects of varying insemination time on sex ratio (Rorie et al., 1999).
Although the mechanisms underlying sex selection are still unclear, the data seem to indicate that males are more fragile (Naeye et al., 1971) and that a greater attrition is exerted on them when reproductive/metabolic conditions are non-optimal (Crawford et al., 1998). Cagnacci et al. (2004) indicated that maternal weight and probably maternal metabolism, as important determinants of secondary sex ratio in human offspring. In another study, 75% of the calves born to a high energy group were male, while a low energy group produced only 46% males (Cheryl et al., 2004).
A study on New Zealand Holstein-Friesien dariy cattle (Roche et al., 2006b) revealed a higher probability of male calves in dams that lost less body condition score and body weight from calving to conception. Although primiparous animals do not loose as much body condition post-calving as multiparous cows (Berry et al., 2006).
The present study has a limitation. The cows BCS was not recorded in farm charts and it was not possible to calculate the effect of this factor on sex ratio. Morever there is some practical and inexpensive method of sex ratio modification such as: time of artificial insemination, dietary manipulation of dairy cattle and Body Condition Score (BSC) between calving and conception.
REFERENCES
- Ansari-Lari, M., 2006. Sex ratio at birth in dairy herds in Fars province, Southern Iran. Trop. Anim. Health Prod., 38: 593-595.
CrossRef - Berry, D.P., R.F. Veerkam and P. Dillon, 2006. Phenotypic profiles for body weight, body condition score, energy intake and energy balance across different parities and concentrate feeding levels. Liv. Prod. Sci., 140: 1-12.
CrossRef - Cagnacci, A., A. Renzi, S. Arangino, C. Alessandrini and A. Volpe, 2004. Influences of maternal weight on secondary sex ratio of human offspring. Hum. Rep., 19: 442-444.
CrossRef - Rosenfeld, C.S. and R.M. Roberts, 2004. Maternal diet and other factors affecting offspring sex ratio: A Review. Bio. Rep., 71: 1063-1070.
CrossRefDirect Link - Crawford, M.A., C. Lowy, E. Koukkou, L. Poston and K. Ghebremeskel, 1998. Sex ratio of offspring of diabetics. Lancet, 351: 1515-1516.
CrossRef - Foote, R.H., 1977. Sex ratios in dairy cattle under various conditions. Theriogen, 8: 349-356.
CrossRef - Grant, V., 1996. Sex determination and the maternal dominance hypothesis. Hum. Reprod., 11: 2371-2375.
Direct Link - Gutierrez-Adan, A., Perez-Garnelo, J. Granados, J.J. Garde, M. Perez-Guzman, B. Pintado and J.D.L. Fuente, 1999. Relationship between sex ratio and time of insemination according to both time of ovulation and maturational state of oocyte. Zygote, 7: 37-43.
PubMed - Hilder, E.A., M. H. Fohrman and E.E. Graves, 1944. Relation of various factors to the breeding efficiency of dairy animals and to the sex ratio of the offspring. J. Dairy Sci., 27: 981-992.
CrossRef - King, K.K., G.E.J. Seidel and R.P. Elsden, 1985. Bovine embryo transfer pregnancies. I. Abortion rates and characteristics of calves. J. Anim. Sci., 61: 747-757.
PubMed - Krackow, S., 1995. Potential mechanisms for sex ratio adjustment in mammals and birds. Biol. Rev., 70: 225-241.
CrossRef - Martinez, V., M. Kaabi, F. Martinez-Pastor, M. Alvarez and E. Anel et al., 2004. Effect of the interval between estrus onset and artificial insemination on sex ratio and fertility in cattle: A field study. Theriogen, 62: 1264-1270.
CrossRef - Naeye, R.L., L.S. Burt, D.L. Wright, W.A. Blanc and D. Tatter, 1971. Neonatal mortality, the male disadvantage. Pediatrics, 48: 902-906.
Direct Link - Roche, J.R., J.M. Lee and D.P. Berry, 2006. Pre-conception energy balance and secondary sex ratio-Support for the Trivers-Willard hypothesis in dairy cows. J. Dairy Sci., 89: 2119-2125.
CrossRef - Roche, J.R., J.M. Lee and D.P. Berry, 2006. Climatic factors and secondary sex ratio in dairy cows. J. Dairy Sci., 89: 3221-3227.
Direct Link - Rorie, R.W., 1999. Effect of timing of artificial insemination on sex ratio. Theriogenology, 52: 1273-1280.
CrossRefDirect Link - Rorie, R.W., T.D. Lester, B.R. Lindsey and R.W. McNew, 1999. Effect of timing of artificial insemination on gender ratio in beef cattle. Theriogenology, 52: 1035-1041.
CrossRefDirect Link - Ryan, D.P. and M.P. Boland, 1991. Frequency of twin births among Holstein-Friesian cows in a warm dry climate. Theriogenology, 36: 1-10.
Direct Link - Seidel, G.E., 2003. Economics of selecting for sex: The most important genetic trait. Theriogen, 59: 585-598.
CrossRef - Sheldon, B.C. and S.A. West, 2004. Maternal dominance, maternal condition and offspring sex-ratio in ungulate mammals. Am. Nat., 163: 40-54.
CrossRef - Skjervold, H. and J.W. James, 1978. Causes of variation in the sex ratio in dairy cattle. Z. Tierz. Zuchtungsbio., 95: 293-305.
CrossRef - Tamimi, R.M., P. Lagiou, A.L. Mucci, C.C. Hsieh, H.O. Adami and D. Trichopoulos, 2003. Average energy intake among pregnant women carrying a boy compared with a girl. Br. Med. J., 326: 1245-1246.
CrossRef - Trivers, R.L. and D.E. Willard, 1973. Natural selection of parental ability to vary the sex ratio of offspring. Science, 179: 90-92.
CrossRefDirect Link - Vandenbergh, J. and D. Huggett, 1994. Mother's prior intrauterine position affects the sex ratio of her offspring in house mice. Proc. Nat. Acad. Sci., 91: 11055-11059.
PubMed - Wade, G.N. and J.E. Schneider, 1992. Metabolic fuels and reproduction in female mammals. Neurosci. Biobehav. Rev., 16: 235-272.
Direct Link - Wauters, L.A., S.A. Crombrugghe, N. Nour and E. Matthysen, 1995. Do female roe deer in good condition produce more sons than daughters? Behav. Ecol. Sociobiol., 37: 189-193.
Direct Link - Williams, R.G. and S.P. Gloster, 1992. Human sex ratio as it relates to caloric availability. Soc. Biol., 39: 285-291.
Direct Link - Xu, Z.Z., D.L. Johnson and N.D. Burton, 2000. Factors affecting the sex ratio in dairy cattle in New Zealand. Proc. N.Z. Soc. Anim. Prod., 60: 301-302.
Direct Link