
Kandarat Lamchumchang
Department of Pathobiology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
Wannee Jiraungkoorskul
Department of Pathobiology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
Somphong Sahaphong
Department of Pathobiology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
Tawewan Tansatit
Faculty of Veterinary Medicine, Mahidol University, Thailand
Piya Kosai
Department of Pathobiology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
Raviporn Madarasmi
Mahidol University International College, Mahidol University, Salaya Campus, Nakhonpathom 73170, Thailand
Asian Journal of Animal and Veterinary Advances
Year: 2007 | Volume: 2 | Issue: 3 | Page No.: 104-114







ABSTRACT
This study investigated the influence of dietary calcium supplementation to reduce the toxicity of sublethal lead concentration in Nile tilapia with emphasis on biochemical and histopathological analysis. The values of 24, 48, 72 and 96 h LC50 of lead to tilapia were 247.51, 197.47, 193.36 and 182.38 mg L-1, respectively. Therefore, lead concentration tested in the sublethal experiment was 45 mg L-1, which correspond to 25% of the 96 h LC50. Fish were fed with 0, 20 and 60 mg Ca2+ g-1 food for 30 days. In the gills of Pb treatment, edema, lamellar cell hyperplasia, epithelial lifting, lamellar fusion and aneurysm were observed. In the liver, there were blood congestion in sinusoids, vacuolation of hepatocytes and necrosis. In the kidney, glomerulus’s atrophy, tubular swelling and also necrosis were seen. The structural damages could be correlated to the significant increase (p≤0.05) in aminotransferase activities. Fish fed with Ca2+ supplemented diets showed slightly alteration when compare the only Pb treatment groups. These results indicated that dietary Ca2+ will be protective in reducing Pb burdens in fish exposed to environments contaminated with waterborne Pb.
PDF Abstract XML References Citation
How to cite this article
Kandarat Lamchumchang, Wannee Jiraungkoorskul, Somphong Sahaphong, Tawewan Tansatit, Piya Kosai and Raviporn Madarasmi, 2007. Dietary Calcium Reducing Effects of Waterborne Lead Uptake in Nile Tilapia (Oreochromis niloticus). Asian Journal of Animal and Veterinary Advances, 2: 104-114.
DOI: 10.3923/ajava.2007.104.114
URL: https://scialert.net/abstract/?doi=ajava.2007.104.114
DOI: 10.3923/ajava.2007.104.114
URL: https://scialert.net/abstract/?doi=ajava.2007.104.114
INTRODUCTION
Heavy metals enter aquatic environments by a number of pathways. The earth’s crust, geologic weathering phenomena and volcanic activity account for natural sources, but most waterborne lead (Pb) derives from human activities such as mining and smelting, coal burning, cement manufacturing and use in gasoline, batteries and paint (WHO, 1995). Lead toxicity arising from chronic waterborne exposure has been reported for a number of freshwater species and occurs at concentrations as low as 4 μg L-1 (Grosell et al., 2006a). The problem of Pb contaminated surface water and sediment in Klity Village, Kanjanaburi province, Thailand was reported in April 1998 (Tonmanee, 2001). Waterborne Pb causes the disruption of Na+, Cl- and Ca2+ regulation during acute exposure, the induction of spinal deformities and black tails during chronic exposure and disruption in hemoglobin synthesis during both types of exposure (Rogers and Wood, 2004). Generally, divalent metals such as Pb, Cd and Zn are considered Ca2+ antagonists. In an early study, coho salmon (Oncorhynchus kisutch) force-fed gelatin capsules containing 8.4 mg of CaCl2 and then exposed to 1300 μg L-1 of waterborne Pb for 168 h, had reduced Pb tissue burdens (Varanasi and Gmur, 1978). Similarly, several more recent studies (Zohouri et al., 2001; Baldisserotto et al., 2005; Franklin et al., 2005; Grosell et al., 2006b) have shown that dietary Ca2+ is protective against the uptake of both waterborne and dietary Cd, as well as against the uptake of waterborne Zn (Niyogi and Wood, 2006).
In fish, gills are the first target organs of several xenobiotics because of their very large interface area between the external and internal fish environments. Performing vital functions such as gas exchange, ion osmoregulation and nitrogen excretion, the gills are particularly sensitive to adverse environmental conditions and changes in gill epithelia have been considered good indicators of the effects of xenobiotics on fish.
The liver is generally regarded as the central organ of xenobiotic metabolism in fish. Structural damages from the effects of pollutants on liver metabolism have been supported by the results of biochemical studies. Monitoring changes in biochemical parameters may provide early warning indicators of general as well as specific toxicological responses. Plasma aminotransferase also may be used as a biological indicator of tissue lesions caused by chemical pollutants (Gingerich, 1982).
Morphologically and physiologically, the fish kidney is composed of three distinct systems: endocrine, hematopoietic and excretory. Lesions that develop in the kidney may involve one or all of the three tissue systems. Thus, it is essential to study the changes that may occur in different cell types. Knowledge of the sublethal effects of xenobiotic compounds on biochemical and histopathological analysis, are very important for delineating fish health status and for understanding future ecological impact. Nile tilapia (Oreochromis niloticus), is a commercialized freshwater fish that is not only widely known around the world, but it is also one of the most popular fish in Thailand. Whereas comprehensive information is available on the effects of heavy metals exposure on fish, there are few reports on the efficiency of fish dietary supplementation on reducing their toxicity. In view of this background, the present study was designed to examine the efficiency of the dietary Ca2+ intake reduces the toxicity of lead in Nile tilapia via biochemical and histopathological analysis.
MATERIALS AND METHODS
Experimental Fish
This study was performed at the Department of Pathobiology, Faculty of Science, Mahidol University, Bangkok, Thailand, in 2006. Nile tilapia, O. niloticus, 14.50±4.03 g in body weight and 9.20±0.71 cm in total length, were purchased from a commercial hatchery in Thailand. Tap water was filtered to eliminate chemical contamination. The physicochemical characteristics of water were measured daily, according to the experimental procedures described in Standard methods for the Examination of Water and Wastewater (APHA, 2005). Conductivity was measured with Hanna instruments Model 3 DiST WP (Hanna Instruments, Rhode Island, USA). The pH was measured with a Cyberscan 510 (Eutech Instruments, Illinois, USA) and the temperature was measured with a glass mercury thermometer. A 16 h light and 8 h dark photo-period was maintained.
Acclimatization to laboratory conditions for 7 days was done using dechlorinated tap water that had the following physicochemical characteristics: temperature = 27.0±1.0°C, pH = 6.5-6.8, total hardness = 65-75 mg L-1 (as CaCO3), alkalinity = 75-80 mg L-1 and conductivity = 185-210 μmhos cm-1. Chlorine residual and ammonia were below detection limits. Fish were fed twice a day with commercial fish food. The quantity of food was 2% of the initial body weight per day. Lead (II) nitrate (Pb (NO3)2 Merck, Germany) was directly diluted in water to obtain the desired exposure concentrations.
Acute Toxicity Test
The acute toxicity tests were performed according to the US EPA procedure for the static non-renewal technique (US EPA, 2002) as described in the previous study (Jiraungkoorskul et al., 2002). Fish were not fed 48 h before starting and 96 h during the experiment. Preliminary screening was carried out to determine the appropriate concentration range for testing the chemical. The test consisted of a control and at least five concentrations of Pb, three replicates per group, with ten fish in each replicate. At the beginning of the tests and after every 24 h, the symptoms and the number of dead fish were recorded. The results of the median lethality concentration (LC50) at 24, 48, 72 and 96 h were computed using the SPSS probit analysis computer program (Finney, 1971).
Diet Preparation
All diets were prepared with commercially fish food that was used to feed the fish during acclimation. The manufacturer’s specifications for the fish food were of 37% crude protein; 14% crude fat; 3% crude fiber; 12% ash and 1% sodium (Charoen Pokphand Group, Bangkok, Thailand). This fish food was ground in a blender, followed by hydration with approximately 40% v/w deionized water. To prepare the calcium-supplemented diet, the control diet was supplemented with CaCO3 (Merck, Germany) to yield an experimental diet with 20 and 60 mg Ca2+ g-1 dry wt. of food. Calcium carbonate was dissolved in the deionized water and added to the food paste. The resulting paste was mixed and extruded through a pasta maker, air-dried and broken into small pellets by hand. The control diet was prepared by the same method but with the addition of deionized water only.
Sublethal Toxicity Test
Test Concentration
The 96 h LC50 value of nile tilapia exposed to Pb was determined in our laboratory as 182.38 mg L-1 (Fig. 1). In this study, fish were exposed to 45 mg L-1, which correspond to 25% of the 96 h LC50.
Test Procedure
Fish (n = 60) were randomly divided into six groups. Each fish was transferred to each aquarium as follows: group1: normal diet and water (control); group 2: normal diet and waterborne lead; group 3: low Ca2+ supplemented diet (20 mg Ca2+ g-1) and normal water; group 4: high Ca2+ supplemented diet (60 mg Ca2+ g-1) and normal water; group 5: low Ca2+ supplemented diet and waterborne lead; and group 6: high Ca2+ supplemented diet and waterborne lead. All glass flow-through aquaria (50x50x120 cm3) with continuous aeration were filled with 200 L of dechlorinated tap water whose physicochemical characteristics were the same as those described previously.
After 30 days treatment, fish from each group were anesthetized with 0.2 g L-1 MS-222 (tricaine methan sulphonate, Sigma, Germany), weighed and measured. Peripheral blood samples were drawn from the caudal vessel and placed in tube containing lithium heparin and then blood was centrifuged at 12,000 g for 5 min. The plasma was collected and frozen at -20°C until the biochemical study. Following blood withdrawal, the organs (gills, liver and kidney) were removed and prepared for histopathological studies. The relative weight of the organ was expressed as a percentage of the body weight.
Biochemical Studies
Plasma aminotransferase activities of aspartate (AST; EC.2.6.1.1; Bergmeyer and Horder, 1986a) and alanine (ALT; EC. 2.6.1.2; Bergmeyer and Horder, 1986b) were estimated according to the recommendations of the International Federation of Clinical Chemistry. The reaction mixture contained α-ketoglutarate and AST- and ALT-specific substrates in buffer phosphate. The reaction was started by adding aliquots of the serum in the mixture and was measured photometrically at 505 nm.
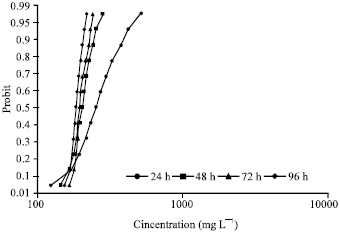 | |
| Fig. 1: | The probit transformed responses of Nile tilapia exposed to LC50 value of lead nitrate |
Specimen Preparation for Light Microscopic Studies
Small pieces of tissues were fixed in the 10% buffered formaldehyde for 24 h, dehydrate through a graded series of ethanol and clear with xylene solutions. They were embedded in a block using melted paraffin at the embedding station (Axel Johnson Lab System, USA). The paraffin blocks were sectioned at 4-5 μm thickness using a rotary microtome (HistoSTAT, Reichert, USA) and stained with hematoxylin and eosin. The tissue glass slides were examined for abnormalities by a Nikon E600 light microscope and photographed by a Nikon DXM 1200 digital camera (Tokyo, Japan) (Humason, 1972).
Semiquantitative Scoring
Histopathological alterations were evaluated semiquantitatively by ranking tissue lesion severity. Ranking from − to + + + depending on the degree and extent of the alteration as follows: (-) no pathological alterations; (+) mild; (+ +) moderate and (+ + +) severe pathological alterations. This ranking was used by Schwaiger et al. (1997) to establish at an overall assessment value of the histopathological lesion for each individual fish gill. Ten slides were observed from each organ and treatment.
Statistical Analysis
All data were expressed as means±SD. A two-way analysis of variance was performed separately tested in each group. The Least-Significant Difference (LSD) was used for determination of significant differences at p≤0.05.
RESULTS
Acute Toxicity Tests
From the probit transformed responses curve of Nile tilapia exposed to 24, 48, 72 and 96 h of Pb, the values of LC50 were log of 2.39, 2.30, 2.29 and 2.26 or 247.51, 197.47, 193.36 and 182.38 mg L-1, respectively (Fig. 1).
General Growth Parameters
Mortality did not occur during the experiments and the control fish did not show any gross or behavioral changes. Otherwise, the treated fish were less active than the controls and a few of the treated fish rested on the bottom of the aquaria. The swimming became slower and there was reduction in their rate of feeding. Overall, the exposure to lead caused only a minor reduction in mean growth rate. These differences were not statistically significant.
Biochemical Studies
Changes in enzyme activity measurements (mean±SD, n =10) in Nile tilapia exposed to lead are presented in Table 1. The values of AST in each group were 85, 140, 84, 61, 120 and 125 U L-1, respectively. As similarly, the values of ALT in each group were 77, 90, 80, 49, 37 and 74 U L-1, respectively. Although lead has a significant increase (p≤0.05) on AST activity but as seen in the figure calcium reduce the effectiveness of lead on these enzymes but it is not statistically significant.
| Table 1: | Change in enzyme activity measurements (mean±SD, n = 10) in Nile tilapia (O. niloticus) exposed to sublethal concentration lead nitrate for 30 days |
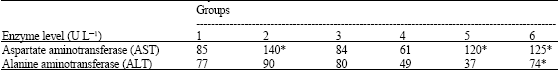 | |
| *Mean difference was significant when compared with the control group at 0.05 level | |
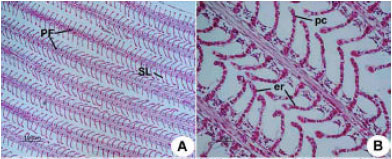 | |
Fig. 2: | Light micrographs of gills of Oreochromis niloticus in each treatment. (A) Control group showing normal appearance of primary filament (PF) and secondary lamellae (SL). (B) High magnification showing erythrocytes (er) within capillary lumen delimited by pillar cells (pc). (C) Group 2, hypertrophy and hyperplasia of secondary lamellar epithelium were seen. (D-E) High magnification in group 2 showing sever edema of epithelial cells (arrows) and aneurysm (*). (F) Group 6, gills showed mild hyperplasia of epithelial cells, Note: Group (1) normal diet + water; (2) normal diet + lead; (3) low Ca diet + normal water; (4) high Ca diet + normal water; (5) low Ca diet+ lead and (6) high Ca diet + lead |
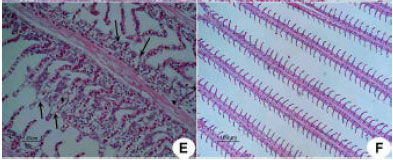
Histopathological Studies
Gills
Control
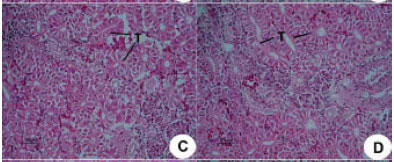
No recognizable changes were observed in the gills of the control and Ca2+ supplemented diet in groups 3 and 4 throughout this experiment. Briefly, there were four gill arches on each side of the buccal cavity. Each gill consisted of a primary filament and secondary lamellae. The primary filament had two rows of secondary lamellae that run perpendicular to each filament. The primary lamellar epithelium was one or two cell layers thick. Chloride cells were identified as large epithelial cells with light cytoplasm, usually present at the base of secondary lamellae. Each secondary lamella was made up of two sheets of epithelium delimited by many pillar cells, which were contractile and separated the capillary channels. One to two erythrocytes were usually recognized within each capillary lumen (Fig. 2A-B).
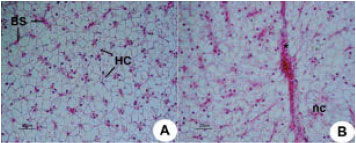 | |
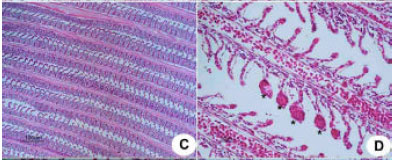
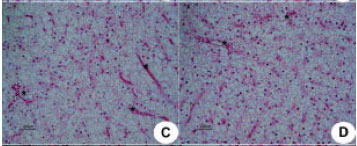
| Fig. 3: | Light micrographs of liver of Oreochromis niloticus in each treatment. (A) Control group showing normal hepatocytes (HC) and blood sinusoids (BS). (B) Group 2, hepatocytes swelling, sinusoid dilation with blood congestion (*), vacuolation and cell necrosis (nc) were seen. (C-D) Groups 3 and 4, mild congestion (*) and vacuolization were also observed. (E-F) Groups 5 and 6, hepatocytes were still swelling and exhibiting necrotic nuclei (nc) |
Treated Groups
Light microscopic study of the gills in each group showed several pathological changes. In group 2, the gills showed severe edema of epithelial cells with some lifting, hypertrophy and hyperplasia of secondary lamellar epithelium (Fig. 2C-E). In groups 5 and 6, filament cell proliferation was quantified by the height of the filament epithelium. The thickening of the primary lamellar epithelium appeared regular. They showed similar alterations as those observed after Pb alone treatment but they were less severe (Fig. 2F). The semiquantitative scoring of gill lesion is shown in Table 2.
Liver
Control
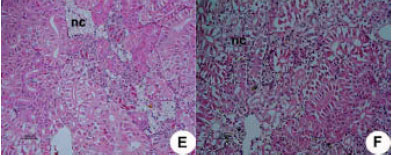
At the light microscopic level, liver had parenchyma, which contains numerous hepatocytes with interspersed connective tissue containing blood vessels and bile ducts. Vacuolization of hepatocytes resulting from glycogen or fat storage could produce histological variability. Other cell types typically found in liver parenchyma include hematopoietic tissue and macrophage aggregates. The venous blood entered and left liver usually through hepatic portal veins, which collect blood from capillaries known as sinusoids. Bile ducts also occurred within the parenchyma of the liver and form between nearby hepatocytes and bile canaliculi anastomose. Eventually the ducts merged to form the common bile duct (Fig. 3A).
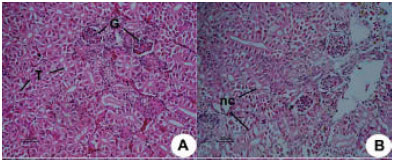 | |
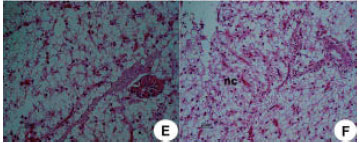
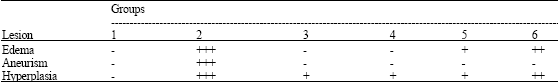
| Fig. 4: | Light micrographs of kidney of Oreochromis niloticus in each treatment. (A) Control group showing normal glomerulus (G) and proximal tubules (T). (B) Group 2, glomeruli atrophy (*) and tubular necrosis (nc) were observed in some areas (C-D) Groups 3 and 4, showing normal appearance of renal tubules (T) similar to those of the control (E-F), tubular necrosis (nc) were also observed |
| Table 2: | Semiquntitative scoring of gill lesion in Nile tilapia (O. niloticus) in each groupa |
 | |
| a Score value: (-) none, (+) mild, (+ +) moderate, (+ + +) severe occurrence | |
| Table 3: | Semiqunatitative scoring of liver lesion in Nile tilapia (O. niloticus) in each groupa |
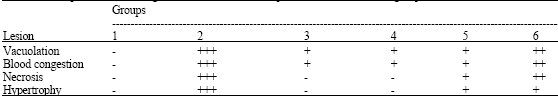 | |
| aScore value: (-) none, (+) mild, (+ +) moderate, (+ + +) severe occurrence | |
Treated Groups
In Pb treated group, the hepatocytes began to swell and there was edema and presence of macrophages. The widening of sinusoids could also be observed which were congested with blood. Vacuolization of hepatocytes was visible along with pyknotic nuclei in many areas (Fig. 3B).
| Table 4: | Semiqunatitative scoring of kidney lesion in Nile tilapia (O. niloticus) in each groupa |
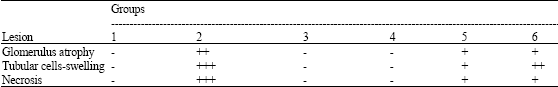 | |
| a Score value: (-) none, (+) mild, (+ +) moderate and (+ + +) severe occurrence | |
In groups 3 and 4, mild congestion and vacuolization were observed in these types of treatment (Figs. 3C-D). In groups 5 and 6, the hepatocytes were still swelling and exhibiting necrotic nuclei, they showed similar alterations as those observed after Pb treatment but they were less severe (Figs. 3E-F). The semiquantitative scoring of liver lesion is shown in Table 3.
Kidney
Control
No recognizable changes were observed in the kidney of the control group. At the light microscopic level, the kidney was composed of numerous renal corpuscles with developed glomeruli and a system of tubules. The proximal tubule composed low columnar epithelial cells with round basal nuclei and brush border (Fig. 4A).
Treated Groups
In Pb treated group, some glomeruli were collapsed or atrophy and tubular necrosis; there were also observed apoptotic cells and necrosis in some areas (Fig. 4B). In groups 3 and 4, the epithelial cells of many tubules had shown fewer damages, their morphology similar to those of the control (Fig. 4C-D). In groups 5 and 6, their changes occurred similarly the Pb treated group but they were less severe (Figs. 4E-F). The semiquantitative scoring of kidney lesion is shown in Table 4.
DISCUSSION
In the fish gills, serve as a major organ for osmotic and ion regulation and respiration. Because of the highly vascular structure of the gill epithelium, it is a primary target for waterborne toxicants. Histopathological alteration on gill epithelial in several previous studies have been described similarly in this study i.e., gill cells degeneration, uplifting of epithelial, necrosis, lesions and inflammatory infiltration. The first sign of lesions in the present study included edema of epithelial cells. The secondary lamellae showed capillary congestion or aneurism, similar to those reported in Puntius altus exposed to cadmium (Jiraungkoorskul et al., 2006), Poronotus triacanthus exposed to copper (Jiraungkoorskul et al., 2007). The lamellar aneurism resulted from the collapse of the pillar cell system and the breakdown of vascular integrity with a release of large quantities of blood that push the lamellar epithelium outward. Otherwise, thickening of the primary lamellar epithelium, hypertrophy and hyperplasia of epithelial, chloride cells, lifting and fusion of secondary lamellae were also observed. The distal extremities of secondary lamellae were bent, with a reduction of the interlamellar space. Several studies pointed out that chloride cell hyperplasia occurred in response to the need to eject the toxicant absorbed by the gills (Gill et al., 1988; Marshall and Grosell, 2005).
In the fish liver not only acts as a storage organ, but is also the primary site for detoxification mechanisms (Olsson et al., 1996). The histopathological alterations in the liver observed in the present study were sinusoid dilation with blood congestion, vacuolation, hemosiderin accumulation and cell necrosis. The vacuolation of hepatocytes might indicate an imbalance between the rate of synthesis of substances in the parenchymal cells and the rate of their release into the systemic circulation (Gingerich, 1982). Toxic of Pb can result in the injured cells dying by necrosis. The structural damages could be correlated to the significant increase (p≤0.05) in AST and ALT activities. Low levels of these enzyme activities are usually found in the blood. There will be an additional release of these enzymes in the bloodstream if there is any damage in the body.
The histopathological alterations occurred in the kidney in this study were dilation of Bowman’s space, glomeruli atrophy and tubular swelling. The degeneration and necrosis of tubular epithelial cells of the first proximal tubule were also observed. Because the excretion of divalent ions is a major function of the renal tubular epithelium, pollution with heavy metals would be highly likely to affect these cells.
Previous studies have found significant reductions in whole body Cd uptake from the water and diet when rainbow trout were fed a diet supplemented with 60 mg CaCO3 g-1 (Baldisserotto et al., 2004b, 2005; Franklin et al., 2005), or 60 mg CaCl2 (Zohouri et al., 2001; Baldisserotto et al., 2004a). Freshwater fish have two primary uptake pathways for essential ions i.e., Ca2+, Na+, the gills as waterborne ions and the gastrointestinal tract as dietary ions. They can regulate the total uptake by changing the proportion of each kind of uptake depending on the environmental situations. For example, tilapia (O. mossambicus) up-regulate intestinal Ca2+ uptake when living in water with low Ca2+ concentration (Flik et al., 1995). Therefore, if the fish can acquire more ions via the gastrointestinal route, they may decrease branchial ion uptake rates and thereby subsequently reduce the uptake of metals sharing the common branchial pathway. For example, Cd2+, Zn2+ shares the same transport pathway with Ca2+ (Verbost et al., 1989; Niyogi and Wood, 2004) and previous studies have shown that dietary Ca2+ supplementation decreased waterborne Ca2+uptake and subsequently waterborne Cd2+uptake (Zohouri et al., 2001; Baldisserotto et al., 2004ab, 2005). There is no available data of Ca2+ concentrations in the natural diet of feral fish, but Ca2+ is available in abundance in crustacean exoskeleton and mollusk shells. Interestingly though, Sherwood et al. (2000) reported that wild yellow perch (Perca flavescens) in heavy metal impacted lakes tend to eat relatively more invertebrates than fish.
In conclusion, the results presented in this study show that the efficacy of dietary calcium supplementation in reducing biochemical and histopathological alterations associated with sublethal exposed to waterborne lead uptake in fish.
ACKNOWLEDGMENTS
This study was funded by the Thailand Research Fund (TRF) and the Commission on Higher Education: the New Researchers Grant 2006 and in part by Mahidol University International College and Faculty of Science, Mahidol University.
REFERENCES
- Baldisserotto, B., C. Kamunde, A. Matsuo and C.M. Wood, 2004. A protective effect of dietary calcium against acute waterborne cadmium uptake in rainbow trout. Aquat. Toxicol., 67: 57-73.
Direct Link - Baldisserotto, B., C. Kamunde, A. Matsuo and C.M. Wood, 2004. Acute waterborne cadmium uptake in rainbow trout is reduced by dietary calcium carbonate. Comp. Biochem. Physiol., 137C: 363-372.
Direct Link - Baldisserotto, B., M.J. Chowdhury and C.M. Wood, 2005. Effects of dietary calcium and cadmium on cadmium accumulation, calcium and cadmium uptake from the water and their interactions in juvenile rainbow trout. Aquat. Toxicol., 72: 99-117.
CrossRefDirect Link - Bergmeyer, H.U., M. Horder, R. Rej, 1986. International federation of clinical chemistry (IFCC) methods for the measurement of catalytic concentration of enzymes. Part 3. IFCC method for aspartate aminotransferase (L-aspartate: 2-oxoglutarate aminotranferase, EC 2.6.1.1). J. Clin. Chem. Clin. Biochem., 24: 481-495.
PubMedDirect Link - Bergmeyer, H.U. and M. Horder, 1986. International federation of clinical chemistry (IFCC) methods for the measurement of catalytic concentration of enzymes. Part 1 IFCC method for alanine aminotransferase (L-alanine: 2-oxoglutarate aminotranferase, EC 2.6.1.2). J. Clin. Chem. Clin. Biochem., 24: 481-495.
- Finney, D.J., 1971. Probit Analysis. 3rd Edn., Cambridge University Press, London, UK., pp: 76-80.
CrossRefDirect Link - Franklin, N.M., C.N. Glover, J.A. Nicol and C.M. Wood, 2005. Calcium/cadmium interactions at uptake surfaces in rainbow trout: Waterborne vs dietary routes of exposure. Environ. Toxicol. Chem., 24: 2954-2964.
Direct Link - Gill, T.S., J.S. Pant and H. Tewari, 1988. Branchial pathogenesis in a freshwater fish, Puntius conchonius Ham., chronically exposed to sublethal concentrations of cadmium. Ecotoxicol. Environ. Safe, 15: 153-161.
CrossRefDirect Link - Grosell, M., R.M. Geredes and K.V. Brix, 2006. Chronic toxicity of lead to three freshwater invertebrates. Environ. Toxicol. Chem., 25: 97-104.
Direct Link - Grosell, M., R. Gerdes and K.V. Brix, 2006. Influence of Ca, humic acid and pH on lead accumulation and toxicity in the fathead minnow during prolonged water-borne lead exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 143: 473-483.
PubMedDirect Link - Jiraungkoorskul, W., E.S. Upatham, M. Kruatrachue, S. Sahaphong, S. Vichasri-Grams and P. Pokethitiyook, 2002. Histopathological effects of roundup, a glyphosate herbicide, to Nile tilapia (Oreochromis niloticus). ScienceAsia, 28: 121-127.
Direct Link - Jiraungkoorskul, W., S. Sahaphong, K. Niwat and K. Myung-Huk, 2006. Histopatholo gical study: The effect of ascorbic acid on cadmium exposure in fish (Puntius altus). J. Fisher. Aquat. Sci., 1: 191-199.
Direct Link - Jiraungkoorskul, W., S. Sahaphong and K. Niwat, 2007. Toxicity of copper in butterfish (Poronotus triacanthus): Tissues accumulation and Ultrastructural changes. Environ. Toxicol., 22: 92-100.
Direct Link - Niyogi, S. and C.M. Wood, 2004. Kinetic analyses of waterborne Ca and Cd transport and their interactions in the gills of rainbow trout (Oncorhynchus mykiss) and yellow perch (Perca flavescens), two species differing greatly in acute waterborne Cd sensitivity. J. Comp. Physiol., 174B: 243-253.
Direct Link - Niyogi, S. and C.M. Wood, 2006. Interaction between dietary calcium supplementation and chronic waterborne zinc exposure in juvenile rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol., 143C: 94-102.
Direct Link - Olsson, P.E., A. Larsson and C. Haux, 1996. Influence of seasonal changes in water temperature on cadmium inducibility of hepatic and renal metallothionein in rainbow trout. Mar. Environ. Res., 42: 41-44.
Direct Link - Rogers, J.T. and C.M. Wood, 2004. Characterization of branchial lead-calcium interactions in the freshwater rainbow trout (Oncorhynchus mykiss). J. Exp. Biol., 207: 813-825.
CrossRefDirect Link - Schwaiger, J., R. Wanke, S. Adam, M. Pawert, W. Honnen and R. Triebskorn, 1997. The use of histopathological indicators to evaluate contaminant-related stress in fish. J. Aquat. Ecosyst. Stress Recovery, 6: 75-86.
CrossRefDirect Link - Varanasi, U. and D.J. Gmur, 1978. Influence of water-borne and dietary calcium on the uptake and retention of lead by coho salmon (Oncorhynchus kisutch). Toxicol. Applied Pharmacol., 46: 65-75.
CrossRef - Verbost, P.M., J. Van-Rooij, G. Flik, R.A.C. Lock and S.E. Wendelaar Bonga, 1989. The movement of cadmium through freshwater trout branchial epithelium and its interference with calcium transport. J. Exp. Biol., 145: 185-197.
Direct Link