
Taixiang Zhang
Laboratory of National Animal TSE, College of Veterinary Medicine, China Agricultural University, Beijing, China
Hongxiang Liu
Departrnent of Histopathology, Addenbrooke’s Hospital,
University of Cambridge, Cambridge, UK
Xiangmei Zhou
Laboratory of National Animal TSE, College of Veterinary Medicine, China Agricultural University, Beijing, China
Xiangmei Zhou
Laboratory of National Animal TSE, College of Veterinary Medicine, China Agricultural University, Beijing, China
Deming Zhao
Laboratory of National Animal TSE, College of Veterinary Medicine, China Agricultural University, Beijing, China
Asian Journal of Animal and Veterinary Advances
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 23-32







ABSTRACT
Transmissible Spongiform Encephalopathy (TSE) or prion disease has been reported in many herbivores but not in equus and the species barrier might be playing a role in resistance of these species to the disease. Therefore, analysis of genotype of prion protein (PrP) in these species may help understand the transmission of the disease. Dezhou donkey is a species of equus reared widely in China for service, food and medicine but its PrP gene has not been studied. Based on the reported PrP sequence in GenBank we designed primers and amplified, cloned and sequenced the PrP gene of Dezhou donkey. The sequence analysis showed that the Dezhou donkey PrP gene was consisted of an open reading frame of 767 nucleotides encoding 256 amino acids. Sequence comparison showed that the PrP gene of Dezhou donkey and horse shared 91.53% homology in nucleic acids and 94.03% homology in amino acids. Amino acid residues unique to donkey as compared with horse, mink, bovine, goat, human, elk and hamster were identified. The results provided the PrP gene data for an additional equus species, which should be useful to the study of the species barrier of TSE.
PDF Abstract XML References Citation
How to cite this article
Taixiang Zhang, Hongxiang Liu, Xiangmei Zhou, Xiangmei Zhou and Deming Zhao, 2006. Molecular Cloning and Sequence Analysis of Prion Protein Gene of Dezhou Donkey in China. Asian Journal of Animal and Veterinary Advances, 1: 23-32.
DOI: 10.3923/ajava.2006.23.32
URL: https://scialert.net/abstract/?doi=ajava.2006.23.32
DOI: 10.3923/ajava.2006.23.32
URL: https://scialert.net/abstract/?doi=ajava.2006.23.32
INTRODUCTION
Transmissible Spongiform Encephalopathy (TSE) is a group of sub-acute infectious diseases in human and animals including scrapie in ovine, bovine spongiform encephalopathy in cattle, chronic wasted disease in deer, transmissible mink encephalopathy in mink, feline spongiform encephalopathy in cat and Creutzfeldt-Jakob disease in human (Prusiner, 1991; Prusiner , 1997). It is understood that the key of development of these diseases is the transformation of a cellular prion protein (PrPc) into PrPsc, a diseased form of PrP characterized by a β-sheet-rich topology and anti-proteolytic property. Misform PrPsc has a tendency of aggregation, shaping first into short amyloidal fiber and then aggregated amyloidal spot before becoming the spongiform-like degenerative changes in brain, which can be identified by clinic detection.
Dezhou donkey is a domestic breed of donkey widely reared in northern China. It is a species of asinus, belonging to Equus, Equide of Perissodactyla, by biological classification. Dezhou donkey is mainly used for service and meat. Since the outbreak of the BSE, beef has increasingly been substituted by meat from other animals including donkey. In addition, donkey skin is used as the main ingredient of Ajiao, an essential blood-nourishing traditional Chinese medicine that has been certified by FDA in the USA and is widely exported to European and American market. Donkey is herbivore, like bovine and ovine. While both bovine and ovine are susceptible to the prion disease, up to the present there has been no report of the TSE cases in donkey as well as horse. It was thought that the species barrier might be playing a role in resistance of these species to the prion disease.
Prion species barrier is a phenomenon of one species being able to protect the prion infection from other species (Hill and Collinge, 2002). There are species particularity and strain particularity of prion in the process of prpsc propagation. The species particularity is that one species can protect the prion attack from the other species, representing the prolonged preclinical period or even no occurrence of the disease. The essential reason of species particularity is the difference in primary and secondary structure of PrP between the recipient and host species(Hill et al., 2000). Until now, the PrP gene has been cloned in human and over 30 other animals (Wopfner et al., 1997). The DNA of PrP is consisted of a single exon and its coded protein is highly conserved, with human and many species of animals sharing 92.9-99.6% of homology in PrP amino acids. Variation or the polymorphism of PrP gene in human being, mouse, sheep, bovine and deer is considered to be associated with the prion disease (Liberski and Brown, 2004; Aguzzi and Polymenidou, 2004).
Studies on the PrP gene in equus are very limited. The available data showed that the PrPs of the domestic horse (Equus familiaris caballus) and the falabella horse (Equus przewalskii caballus) are identical on the amino acid level and have two substitutions compared to the zebra PrP (Equus quagga boehmi) (Wopfner et al., 1997). However, there is no report about the PrP gene in donkey. In the present study, the authors cloned and sequenced the PrP gene of Dezhou donkey and analyzed the sequence of the PrP gene with that from other animals. The results provided the PrP gene data for an additional equus species, which should be useful to the study of the species barrier of TSE and also of interest to public healthy.
MATERIALS AND METHODS
Biological Materials
Genomic DNA was extracted from peripheral blood of six Dezhou donkeys using a Genomic DNA Rapid Isolation Kit for Blood (Biodev, USA) according to the manufacturer’s instructions.
Polymerase Chain Reaction (PCR)
Each DNA sample was PCR-amplified for the PrP gene in a final reaction volume of 50 μL containing 250 ng of genomic DNA, 20 pmol of each primer, 250 uM of dNTP (Pharmacia) and 1.5 unit of Taq polymerase (Perkin Elmer Cetus). The primers were deduced from white-tail deer PrP sequence in GenBank (accession number AY360089), which were 5'-ATGGTGAAAAGCCACATAGGC-3' for sense primer and 5'-GAAGATAATGAAAACAGGAA G-3' for anti-sense primer. The PCR procedure comprised of an initial denaturation at 94°C for 5 min and 30 cycles of denaturation at 94°C for 45 sec, annealing at 58 °C for 50 sec and elongation at 72°C for 50 sec. Finally, an extension step at 72°C for 10 min was performed (Yang et al., 2005; Zhang et al., 2004; Wang et al., 1998).
Cloning, sequencing and analysis of the amplified PrP products
Following confirmation of the amplified DNA fragment of 767 bp on a 1% agarose gel with DNA marker (DL2000, TAKALA), the PCR products were cloned into T-easy vectors (Promega, USA) and the propagated plasmids were prepared using Qiagen Plasmids Mini Kit (Qiagen, UK). Four positive clones were identified from each sample by restriction enzyme digestion with EcoRI and sequenced in both directions on an ABI Prism 377 automated DNA sequencer (Applied Biosystems, Foster City, USA). The nucleotide and amino acid sequences were analyzed using the DNAMAN software.
RESULTS AND DISCUSSION
The amplified PCR product of Dezhou donkey PrP gene was 767 bp in size, similar to PrP gene reported in other animals (Fig. 1).
Sequencing of the amplified PCR fragments revealed a single open reading frame of 767 nucleotides encoding 256 amino acids. The newly identified sequence has been deposited into GenBank under the accession number AY968590.
Sequence comparison showed that the Dezhou donkey PrP gene had 91.53% homology in nucleic acid sequence and 94.03% homology in amino acids with that of domestic horse (GenBank AF117317) (Fig. 2). Dezhou donkey PrP gene also shared high homology in both nucleic acid and amino acid sequences with those from other mammals, including elk, human, mink, bovine, goat and hamster (Table 1).
In keeping with the structure of PrP in other animals, Dezhou donkey PrP had an N-terminal 23 amino acid signal peptide.
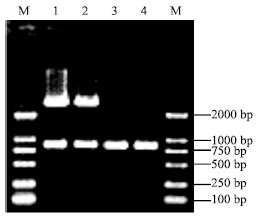 | |
| Fig. 1: | Agarose gel electrophoresis of recombinant T-easy-vectors following digestion with EcoRI (M: marker DL2000; 1 and 2: EcoRI digested recombinant T-easy-vectors; 3 and 4: PCR products using recombinant T-easy-vectors as template) |
| Table 1: | Homologies of PrP between Dezhou donkey and other species |
 | |
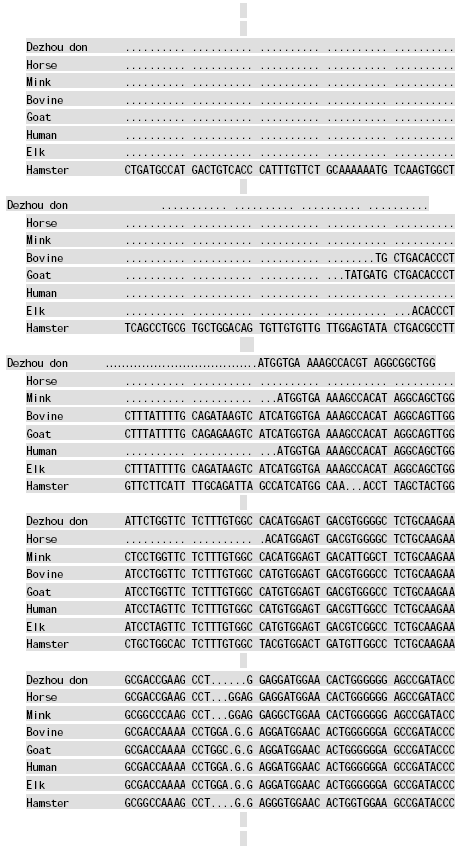 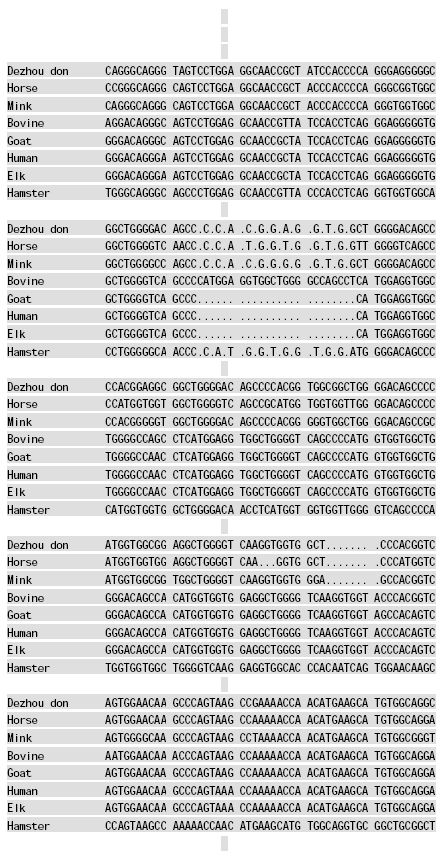 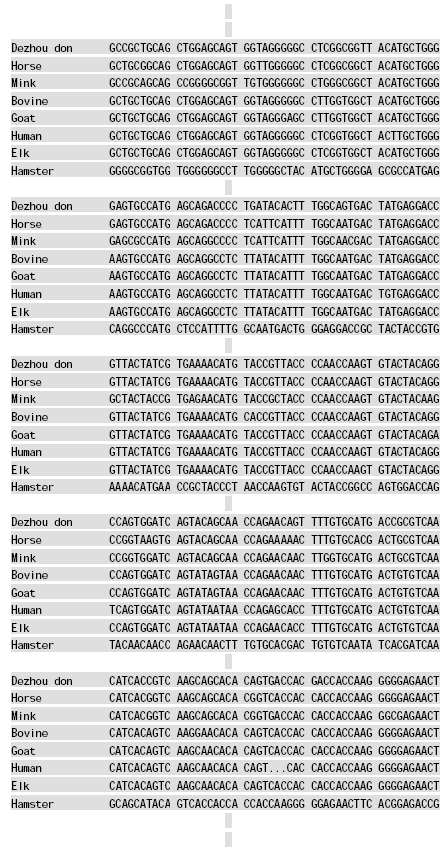 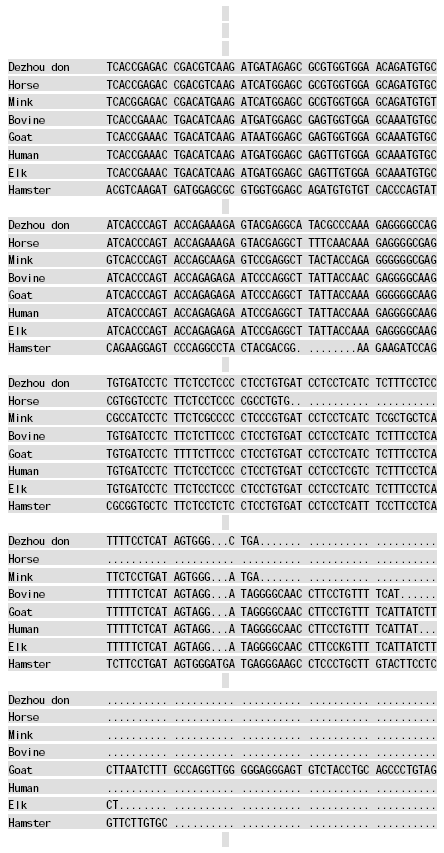 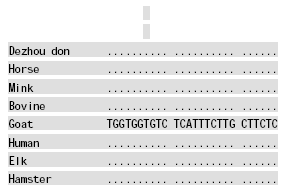 | |
| Fig. 2: | The comparison of nucleic acid sequence of PrP gene between Dezhou donkey and seven other animals (accession number of GenBank are AF117317, U08952, AY367640, 326330, AY 458651, AY748455, 33958) |
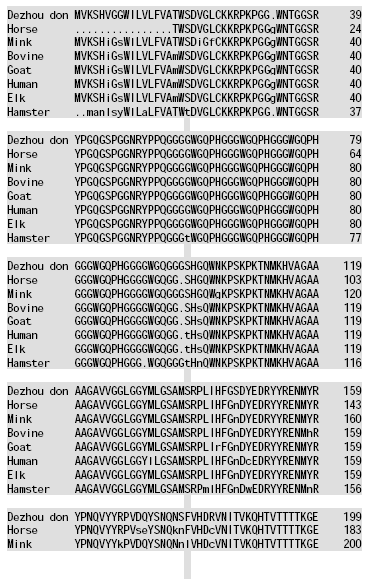 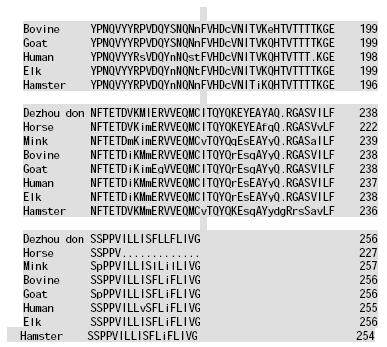 | |
| Fig. 3: | The comparison of amino acid sequence of PrP between Dezhou donkey and seven other animals. (Accession numbers of GenBank are AF117317, U08952, AY367640, AY326330, AY 458651, AY748455 and M33958) |
This was followed by a glycine and proline-rich region (24-96) encompassing a N-terminal nonapeptides (PQGGGGWGQ), four repeats of the octapeptides (PHGGGWGQ) and a C-terminal nonapeptides (PHGGGGWGQ). Amino acids 97~112 is a functional domain controlling the topology structure of the PrPc; 113~135 is an α-helix; 157~177 is a hydrophilic helix; 234~256AA is a hydrophobic signal region (Fig. 3).
Comparing with horse, bovine, goat, deer, mouse, human being and mink, Dezhou donkey was unique in its PrP amino acids at the following positions: there was no Gly at position 32, nor in zebra (Wopfner et al., 1997) and there was a Gly at position 97; the positions 146, 182, 209, 225 and 229 were Ser, Arg, Iso, Tyr, Ala respectively in Dezhou donkey but Asp, Cys, Met, Ser and Tyr in other animals analyzed. Overall, the PrP of all species analyzed mainly varied in three regions: 90~103, 167~169 and 206~230. It has been reported that amino acid sites 90~130 are corresponding to the species barrier and 108~189 are related to incubation. Within these regions, Gly at position 100 in Dezhou donkey and horse was substituted with Ser in bovine, goat, human and elk. Outside these regions, positions 225 (Tyr) and 229 (Ala) were unique to Dezhou donkey and horse as compared to other species (Ser and Tyr). While the latter species but not the equus have been reported to be infected by prion disease, it is not known if these residues in particular are implicated in the species barrier in equus.
Recent studies with bovine transgenic mice suggested that amino acid residues 184, 186, 203 and 205 form an epitope that is involved in the control of the species barrier and that amino acids 167, 171, 214 and 218 form an epitope to interact protein X that has been found to participate in the PrPsc formation(Kaneko et al., 1997; Brown et al., 1997). However, analysis of these sites showed no variation of amino acids between Dezhou donkey and the rest of the animals analyzed. As for polymorphism at 136, 154 and 171 amino acid sites that has been reported to be pertinent to impressionable of scrapie (Eghiaian et al., 2004; Mallucci et al., 2003), in which the genotype A136R154R171 induces resistance to scrapie but A136R154Q171 is intermediately sensitive and V136R154Q171 is sensitive to scrapie, the genotype of donkey is ARQ and may therefore be intermediately sensitive to scrapie. However, how these differences influence the conversion of PrPc to PrPsc are yet to be illuminated.
This article was finished in Laboratory of National Animal TSE, College of Veterinary Medicine, China Agricultural University in 2005 and supported by fund of Science and Technology department of Beijing and Education department of China, which code were Z0004100040691-7 and 20050019031.
REFERENCES
- Aguzzi, A. and M. Polymenidou, 2004. Mammalian prion biology one century of evolving concepts. Cell, 116: 313-327.
Direct Link - Brown, D.R., K. Qin, J.W. Herms, A. Madlung and J. Manson et al., 1997. The cellular prion protein binds copper in vivo. Nature, 390: 684-697.
PubMedDirect Link - Eghiaian, F., J. Grosclaude and S. Lesceu, 2004. Insight into the prpc-prpsc conversion from the structures of antibody-bound ovine prion scrapie-susceptibility variants. Proc. Natl. Acad. Sci. USA., 101: 10254-10259.
Direct Link - Hill, A.F., S. Joiner, J. Linehan, M. Desbruslais, P.L. Lantos and J. Collinge, 2000. Species-barrier-independent prion replication in apparently resistant species. Proc. Natl. Acad. Sci. USA., 97: 10248-10253.
Direct Link - Hill, A.F. and J. Collinge, 2002. Species-barrier-independent prion replication in apparently resistant species. APMIS: Acta Pathol., 110: 44-53.
Direct Link - Kaneko, K., L. Zulianell, C.M. Cooper, A.C. Wallace and T.L. James et al ., 1997. Evidence for protein X binding to a discontinous epitope on the cellular prion protein during scrapie prion propagation. Proc. Natl. Acad. Sci. USA., 94: 10069-10074.
Direct Link - Liberski, P.P., D.C. Guiroy, E.S. Williams, A. Walis and H. Budka, 2001. Deposition patterns of disease-associated prion protein in captive mule deer brains with chronic wasting disease. Acta Neuropathol. Berlin, 102: 496-500.
CrossRef - Mallucci, G., A. Dickinson, J. Linehan, P.C. Klohn, S. Brandner and J. Collinge, 2003. Depleting neuronal prp in prion infection prevents disease and reverses spongiosis. Science, 302: 871-874.
CrossRef - Wopfner, F., G. Weidenhofer and R. Schneider, 1999. Analysis of 27 mammalian and 9 avian prps reveals high conservation of flexible regions of the prion protein. J. Mol. Biol., 289: 1163-1178.
CrossRef - Wang, D., X. Wang, S. Han, B. Tian and Z. Rao, 1998. Cloning and sequencing of the chinese bovine prion protein prpc gene. Wei Sheng Wu Xue Bao, 38: 417-421.
Direct Link - Yang, J.M., D.M. Zhao, H.X. Liu, N. Li, Y.X. Hao, Z.Y. Ning and X.H. Qin, 2005. Comparative analysis of the prion protein open reading frame nucleotide sequences in peacock and parakeet. Virus Genes, 30: 193-196.
CrossRef