
Sogolo L. Lebelo
Department of Life and Consumer Sciences, University of South Africa, Cnr Christiaan de Wet and Pioneer Ave, Private Bag X6,
1710 Florida, South Africa
Gerhard Van Der Horst
Department of Medical Biosciences, University of the Western Cape, Private Bag X17, 7530 Bellville, South Africa
Asian Journal of Animal Sciences
Year: 2016 | Volume: 10 | Issue: 4-5 | Page No.: 247-254







ABSTRACT
Background: Chlorocebus aethiops commonly known as the vervet monkey has been constantly used as an important animal model for human-related studies. Materials and Methods: The process of spermiogenesis of the vervet monkey was investigated with electron microscopic techniques. Twenty-nine male vervet monkeys were used in this study. Results: Based on the ultrastructural changes, spermatids were divided into 12 steps. The acrosome develops from merging transport vesicles that arise from the Golgi and subsequently partition into acrosomal cap. Initial spermatids have dense cytoplasm with scattered mitochondria, a well-developed Golgi body and nuclei with patches of heterochromatin. Most of the cytoplasmic structures and mitochondria have undergone profound transformations. Nuclei and cell elongate and chromatin begins to condense near the nuclear envelope. An acrosomal complex appears at the tip of the nucleus. Manchette is clearly observed in the developing spermatids. The acrosomal filament runs the entire length of the nucleus. As a final step of spermiogenesis, cytoplasm has been significantly reduced. Mitochondria surround the midpiece as the anlage of the annulus forms. Phagocytic vesicles are observed in association with the developing spermatids. Conclusion: The results indicate that the processes of spermiogenesis of the vervet monkey are found to be fundamentally similar to that of other mammalian species, non-human primates and in particular to that of human. The findings of this study could also assist in early detection of defective sperm development.
PDF Abstract XML References Citation
Received: June 20, 2016;
Accepted: July 16, 2016;
Published: August 15, 2016
How to cite this article
Sogolo L. Lebelo and Gerhard Van Der Horst, 2016. Ultrastructural Changes Occurring During Spermiogenesis of the Vervet Monkey, Chlorocebus aethiops. Asian Journal of Animal Sciences, 10: 247-254.
DOI: 10.3923/ajas.2016.247.254
URL: https://scialert.net/abstract/?doi=ajas.2016.247.254
DOI: 10.3923/ajas.2016.247.254
URL: https://scialert.net/abstract/?doi=ajas.2016.247.254
INTRODUCTION
Spermiogenesis is defined as the process of differentiation of the newly formed haploid round spermatids to mature spermatozoa in the seminiferous epithelium just before their release into the lumen of the seminiferous tubule1,2. Spermiogenesis begins after spermatocytes have completed meiotic reductive division2. This post-meiotic process involves complex morphological, biochemical, physiological alterations and orientation of germ cells resulting in the formation of spermatozoa2-4. The major events in this extraordinarily complex process involve elaboration of the acrosome from the Golgi apparatus, condensation and elongation of the nucleus, formation of motile flagellum, centrosome disintegration and extensive shedding of the cytoplasm. The complex structural and morphological evolution of the spermatozoon from a round haploid cell with partial loss of certain organelles and the formation of new ones is an interesting biological phenomenon which has drawn attention to many researchers because of its biochemical changes and species specific nature5. A comprehensive understanding of the morphology and ultrastructure of male gametes is important for various reasons such as conservation, early detection of defective spermatozoa and phylogenetic evidence6.
In mammals, acrosome biogenesis begins with the fusion of proacrosomal granules synthesized by the Golgi apparatus in pachytene spermatocytes, similar to the formation of secretory granules in many other cell types7,8. Acrosome is an important component of the spermatozoon head and develops from four phases namely; Golgi, cap, acrosomal and maturation phases. Each of these phases is supported by precise and orderly regulation of various genes9. The different phases of spermiogenesis are distinguished by the morphological appearance of the developing acrosome, changing shape of the nucleus and the formation of the axoneme8. These changes also involve nuclear and chromatin condensation as well as shaping of the sperm head by microtubule-based structure called the manchette2. The Golgi phase involves the acrosomal vesicle attaching to the nuclear envelope and grows as a result of the constant arrival and fusion of Golgi-derived vesicles8. The cap phase is characterized by the acrosomal vesicles spreading to cover the anterior half of the nucleus and become further condensed. The acrosomal cap is derived from the reshaping of the membrane of the acrosomal vesicle. This cap consists of an outer and inner acrosomal membrane enclosing the acrosomal contents5. The next phase is the acrosome phase which is characterized by the spermatid’s reorientation itself so that the head becomes deeply embedded in the sertoli cell and points towards the basal lamina. The developing flagellum extends into the lumen of the seminiferous tubule and this reorientation is crucial because the head of the spermatozoa is still attached to the sertoli cell. The Golgi complex detaches itself from the anterior pole of the nucleus and migrates freely in the cytoplasm when the acrosome ceases growing. The maturation phase is characterized by the pinching off and further reduction of excess cytoplasm and possibly extrusion of any organelles not needed for delivering the sperm’s genetic information to the ovum1,10,11. The description of these phases forms the basis for determining the kinetics of spermatogenesis and is a prerequisite to define stages of spermatogenesis in a given species12.
Cholorocebus aethiops formerly known as Cercopithecus aethiops is commonly referred to as the African green monkey, vervet, tantulus, savannah and the grivet monkey. Over the past three decades, the vervet monkey Chlorocebus aethiops has been constantly used in biomedical study as an animal model for human-related problems such as in virology and reproductive studies13. In addition, due to overwhelming interest in the growing incidence of human infertility, this has prompted the need to fully understand sperm development and pathologies in the application of assisted reproductive technologies. Therefore, the main objective of the study was to investigate the ultrastructural changes that occur during the spermiogenesis of the vervet monkey using electron microscopic techniques and evaluate this non-human primate as a potential experimental animal model for human reproductive studies.
MATERIALS AND METHODS
Study animals: The study was performed using 29 male vervet monkeys which were obtained from the National Research Institute for Nutritional Diseases at the South African Medical Research Council, Tygerberg, Cape Town, South Africa. The animals were housed permanently indoors and fed with a basic diet consisting of precooked maize meal with a protein-vitamin and mineral concentrate which was mixed to a stiff, porridge with water and fed in the mornings. Whole wheat brown bread was provided at noon, apples and carrots, washed in chlorinated water are fed in the afternoon. Drinking water was supplied ad libitum via an automatic device14. Ethical clearance was obtained from the Medical Research Council’s Ethical Committee for this study.
The ages of the animals ranged from 5-11 years. Animals were weighed to the nearest 100 g and subsequently anaesthetized with intramuscular injection of Ketamine (Ketalar at 10 mg kg–1 body mass). Immediately, after sacrificing testes were excised. Tissues were processed for both scanning electron microscopy and transmission electron microscopy.
Transmission Electron Microscopy (TEM): Samples were fixed in 2.5% buffered gluteraldehyde and then cut in cubes of 1 mm3 before placed in 1% OsO4 for contrasting for atleast 1 h. The tissues were then rinsed in buffer for 15 min. Specimens were dehydrated in increasing concentrations of ethanol. The specimens were then placed in 1:1 mixture of propylene oxide: Resin (EMS Sciences, UK) overnight for infiltration. The next morning the specimens were placed in 1:3 mixture of propylene oxide and resin for 2 h. The specimens were then placed in fresh 100% resin under vacuum for 2 h. The procedure was followed by embedding specimens in fresh resin. The specimens were then placed in an embedding oven at 70°C for curing. Gold sections (70 nm) made using ultramicrotome (Reichert Jung Ultracut E, Germany) were placed on copper grids and left to dry. Grids were contrasted or stained using heavy metal stains, both lead citrate and uranyl acetate (EMS Sciences, UK). Grids were viewed by means of a Jeol JEM1010 (Jeol Inc., Tokyo) Transmission Electron Microscope (TEM). Selected areas were photographed and micrographs printed for examination.
RESULTS
Electron microscopy
Spermiogenesis: The differentiation of spermatids can be divided into 12 steps according to the morphological changes that occur during the formation of the acrosome and the development of motile flagellum as well as condensation of the nucleus. Steps 1-3 corresponded to the Golgi phase of the spermatid development, steps 4-5 corresponded to the cap phase, steps 6-9 to the acrosomal phase and steps 10-12 to the maturation phase. These phases tend to overlap and there is no clear distinction between them hence, other phases will be described as early, mid and late phases. It should also be noted that different steps can be identified close to each other in the same seminiferous tubule. Figure 1-11 show the different steps of spermatid development during the spermiogenesis of the vervet monkey.
Step 1-3 (Spermatids): These groups of spermatids are characterized by round nuclei containing chromatin points attached to a nuclear envelope with a single nucleolus. Rough endoplasmic reticulum and mitochondria can be seen throughout the cytoplasm.
 | |
| Fig. 1: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 1: Spherical nucleus and the single round nucleolus, Golgi apparatus is found closer to the nucleus, Step 5: Acrosome developing, AC: Acrosome, CH: Chromatin, CP: Cap phase, F: Filament, G: Golgi, Gp: Golgi phase, N: Nucleus, NO: nucleolus, PV: Phagocytic vesicle and SER: Smooth endoplamic reticulum |
 | |
| Fig. 2: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 2: Round nucleus and nucleolus are observed, Golgi is seen close to the nucleus, Step 4: Spherical nucleus and a single round nucleolus, Acrosome is starting to form AC: Acrosome, G: Golgi, Gp: Golgi phase, N: Nucleus, NO: Nucleolus and PV: Phagocytic vesicle |
Centrioles can be seen near the plasma membrane. The Golgi complex starts to develop and associated with small vesicles.
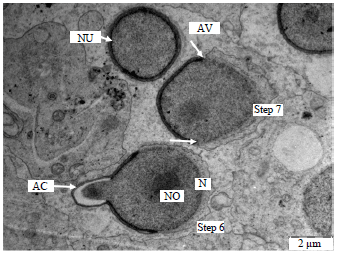 | |
| Fig. 3: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 6: Spherical nucleus and a single round nucleolus, Step 7: Nucleolus is disappearing, AC: Acrosome, AV: Acrosomal vacuole, N: Nucleus, NO: Nucleolus, PV: Phagocytic vesicle, SER: Smooth endoplamic reticulum and arrows showing migration of the acrosome |
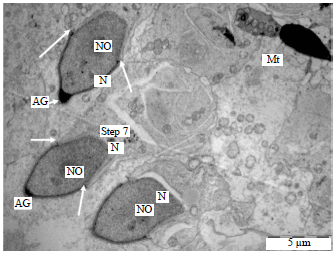 | |
| Fig. 4: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 7: Acrosomal granule, arrows showing migration of the acrosome, AG: Acrosomal granule, Mt: Mitochondria, N: Nucleus and NO: Nucleolus |
The spermatids are further characterized by the presence of a proacrosomal vesicle situated near the highly developed Golgi apparatus towards the end of this phase. Golgi apparatus consisted of several lamellae arranged in a crescent with its concavity facing the acrosome. The endoplasmic reticulum is also observed near the Golgi apparatus and numerous small vesicles were situated between these two organelles (Fig. 1, 2).
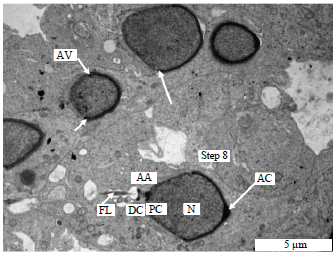 | |
| Fig. 5: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 8. Anlage of annulus and the development of centrioles, arrows showing migration of the acrosome, AA: Anlage of annulus, AC: Acrosome, AV: Acrosome vacoule, DC: Distal centriole, FL: Flagellum, N: Nucleus and PC: Proximal centriole |
Steps 4-5 (spermatids): The spermatids are characterized by development of proacrosomal vacuoles. These vacuoles greatly increase during step 5 and attach to the nuclear envelope. The nuclear shape is still spherical, chromatin condensation starts forming. The acrosome associates more intensely with the nuclear envelope. The production and subsequent fusion of proacrosomal vesicles to the acrosome by the Golgi complex continue in this phase. There are some features in these spermatids which are similar to those in the step 1-3 spermatids (Fig. 1, 2).
Steps 6-9 (spermatids): The major characteristic of the spermatids of these steps is the condensation of the acrosome. The Golgi complex is poorly developed. Associated with the developing flagellum and connected to the annulus was the presence of a proximal and distal centrioles. The nucleus migrates close to the plasma membrane. The acrosome covers a significant portion of the nuclear surface. Nuclear condensation continued during this phase when it was virtually complete and the spermatid head had assumed its final shape which is elongated. The spermatids in these steps are also characterized by the formation of annulus and manchette.
The annulus had not commenced its caudal migration and the mitochondria were not yet arranged helically around the axial filament (Fig. 3-6).
 | |
| Fig. 6: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 1: Spherical nucleus and a single round nucleolus, Step 9: Different stages of step 9, Note the condensation of the chromosome, nucleus closer to the plasma membrane, Note early step 10, Ch: Chromatin, G: Golgi, N: Nucleus, NO: Nucleolus and Mt: Mitochondria |
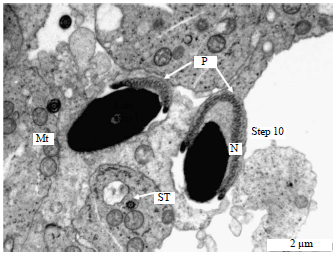 | |
| Fig. 7: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 10: Presence of the perforatorium aroung the nucleus of the spermatid, Mt: Mitochondria, N: Nucleus, P: Perforatorium and ST: Sperm tail |
Steps 10-12 (spermatids): These are the final steps of the spermatid development. Steps 10-11 spermatids are characterized by the formation of the residual body. Spermiation occurs during in the late step 11. During the final development of the sperm tail, prior to the caudal migration of the annulus in steps 10-12 spermatids, a swelling of the posterior middle piece became apparent.
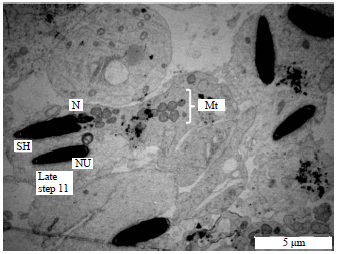 | |
| Fig. 8: | Electron micrograph of the spermatid showing the process of spermiogenesi in the vervet monkey, Chlorocebus aethiops, Step 11: Changes in the nucleus of the spermatid, Mt: Mitochondria, N: Nucleus and SH: Spermatid head |
 | |
| Fig. 9: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 11: Changes in the nucleus of the spermatid, L: Lipid droplet, Mt: Mitochondria and SH: Spermatid head |
Mitochondrial sheath, midpiece and the fibrous sheath are formed (Fig. 7-11). Figure 10 also illustrates step 3 of spermatid development.
DISCUSSION
The process of spermiogenesis is a complex event which involves morphological and ultrastructural changes of the developing germ cells. This process has been described by many researchers as a sub phase of the spermatogenesis15. During spermiogenesis, spermatids undergo specific morphological and functional modifications which involve nuclear elongation, chromatin condensation, acrosomal formation and tail development.
 | |
| Fig. 10: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 3: Round nucleus and the presence of the Golgi, Step 9: Migration of the nucleus to the plasma membrane almost filling the open space, Step 12: Elongated spermatid head, almost a complete mature spermatozoon, G: Golgi and Mt: Mitochondria |
 | |
| Fig. 11: | Electron micrograph of the spermatid showing the process of spermiogenesis in the vervet monkey, Chlorocebus aethiops, Step 1: Spherical nucleus of the spermatid, Step 12: Elongated spermatid head, Mt: Mitochondria and SH: Spermatid head |
To understand the process of spermatogenesis, the ultrastructural changes that occur in the developing spermatids need to be investigated and thoroughly described as illustrated in this study. To this knowledge, this is the 1st study to investigate the ultrastructural changes in the process of spermiogenesis in the vervet monkey.
In the present study, the spermiogenesis of the vervet monkey was found to be divided into 12 steps. This finding was similar to those reported in other mammals such as the common marmoset, broad-nosed bat16,17. The number of steps found in the vervet monkey is less than those observed in the Mongolian gerbil where 15 steps were identified18. Nineteen steps of spermatid development were observed in the spermiogenesis of the rat which was the higher number of steps compared to the number of spermatid development steps observed in this study19. It was reported that the number of steps in the differentiation of the spermatid development varies in different species and several studies have described between 6-16 steps16,20-22. This variation of the number of steps could be as a result of the final stages of the development of spermatozoon head of a specific species and the methods used by different researchers in analyzing the spermiogenesis process16.
Events which characterize the process of spermiogenesis, such as chromatin development, formation of the acrosome, elimination of the residual cytoplasm and the development of the flagellum occur across the animal kingdom20. The process of spermiogenesis was described in many animal species such as common tree shrews23, freshwater sting rays24, soft-shelled turtle25 and domestic fowls26 where basic developmental steps were found to be almost similar except for some species-specific processes. In this study, similar processes were also observed in the developmental stages of the vervet monkey spermatids and significant ultrastructure changes were identified.
Studies on the identification of spermatid developmental steps have been done on several vertebrates and in most instances the analysis was based on the acrosome development12. Steps 1-3 were characterized by presence of proacrosomal vesicle situated near the highly developed Golgi apparatus. These steps are the same with all other mammalian studies17. The same features were observed in the study of Mongolian gerbil18. The endoplasmic reticulum was observed situated near the Golgi apparatus and numerous vesicles were found situated between the two organelles. Golgi apparatus consisted of several lamellae arranged in crescent with its concavity facing the acrosome. Steps 4-5 spermatids showed flattening of the proacrosomal vesicle against one pole of the spermatid nucleus and synthesis of acrosomal contents. In the study by Segatelli et al.18 Mongolian gerbil showed similar features. The nuclear shape was still spherical and the chromatin was beginning to condense. The findings from this study showed similar features of the presence of Golgi apparatus in close proximity to the acrosomal system as reported in the broad-nosed bat and Mongolian gerbil16,18.
In steps 6-9, the spermatid’s nuclei become flattened and some condensation of the chromatin granules occurs. The principal feature in this group of spermatids is the condensation of acrosome and the formation of a cap, covering a large surface of the nucleus. The annulus and manchette also start to develop during this period. The same features were identified in the Mongolian gerbil16,18. Nuclear condensation continues in the steps 10-12 when it is virtually complex and the spermatids head had assumed its final shape. The head of the spermatozoon of the vervet monkey is similar to all other mammals with a tapered and dorsoventrally flattened head27,28. Steps 10-12 were characterized by the formation of the residual body and spermiation occurs at this stage. During final development of the sperm tail, prior to the caudal migration of the annulus in steps 10-12 and a swelling of the posterior middle piece become apparent. These steps were also described by Holt and Moore17 in their study on the common marmoset.
The ultrastructural changes occurring during the process of spermiogenesis of the vervet monkey are found to have similar features with processes found in seminiferous epithelium of other mammals and humans. The four major phases of spermiogenesis with distinguishing features which characterized them were observed in this study. It is important to note that these phases tend to overlap and it is quite convenient to label them as early, middle and late phase as described for baboons by Afzelius et al.29. These studies findings are very important in providing information about possible defects in sperm structure such as abnormal sperm morphology. With humans being exposed to various endocrine disruptors and early detection of defects in the process of spermiogenesis or spermiation and could assist in the potential reproductive toxicant administration.
CONCLUSION
In conclusion, the spermiogenesis and the ultrastructural changes that occur in the vervet monkey follow the general pattern that has been described for other mammalian species including humans and other non-human primates. It can be concluded that the vervet monkey being an Old World Monkey could be regarded as a suitable model for studies in spermiogenesis and other reproductive studies such as toxicology and male infertility.
ACKNOWLEDGMENTS
The authors would like to acknowledge NRF for funding this project. We would also like to thank Dr Jurgen Seier and staff of the Primate Unit of the MRC, Parow, Cape Town, for assistance with sampling and Dr C. Baker and her staff members in the Electron Microscopic Unit, Sefako Makgatho Health Sciences University, Pretoria for their assistance in tissue processing.
REFERENCES
- O'Donnell, L., 2014. Mechanisms of spermiogenesis and spermiation and how they are disturbed. Spermatogenesis, Vol. 4.
CrossRefDirect Link - Behr, R., N. Hunt, R. Ivell, J. Wessels and G.F. Weinbauer, 2000. Cloning and expression analysis of testis-specific cyclic 3′, 5′-adenosine monophosphate-responsive element modulator activators in the nonhuman primate (Macaca fascicularis): Comparison with other primate and rodent species. Biol. Reprod., 62: 1344-1351.
PubMedDirect Link - Rathke, C., W.M. Baarends, S. Awe and R. Renkawitz-Pohl, 2014. Chromatin dynamics during spermiogenesis. Biochimica Biophysica Acta (BBA)-Gene Regulatory Mech., 1839: 155-168.
CrossRefDirect Link - Majhi, R.K., A. Kumar, M. Yadav, P. Kumar, A. Maity, S.C. Giri and C. Goswami, 2016. Light and electron microscopic study of mature spermatozoa from White Pekin duck (Anas platyrhynchos): An ultrastructural and molecular analysis. Andrology, 4: 232-244.
CrossRefDirect Link - Soley, T.J., 2016. A comparative overview of the sperm centriolar complex in mammals and birds: Variations on a theme. Anim. Reprod. Sci., 169: 14-23.
CrossRefDirect Link - Moreno, R.D., J. Ramalho-Santos, P. Sutovsky, E.K. Chan and G. Schatten, 2000. Vesicular traffic and Golgi apparatus dynamics during mammalian spermatogenesis: Implications for acrosome architecture. Biol. Reprod., 63: 89-98.
CrossRefPubMedDirect Link - Niu, C.M., J.Q. Guo, H.T. Ma, Z. Zheng and Y. Zheng, 2016. [Sperm acrosome formation-associated genes in mice: Advances in studies]. Natl. J. Androl., 22: 72-76, (In Chinese).
PubMedDirect Link - Leblond, C.P. and Y. Clermont, 1952. Spermiogenesis of rat, mouse, hamster and guinea pig as revealed by the periodic acid-fuchsin sulfurous acid technique. Am. J. Anat., 90: 167-215.
CrossRefDirect Link - Kushner, H., N. Kraft-Schreyer, E.T. Angelakos and E.M. Wudarski, 1982. Analysis of reproductive data in a breeding colony of African green monkeys. J. Med. Primatol., 11: 77-84.
PubMedDirect Link - Seier, J.V., 1986. Breeding vervet monkeys in a closed environment. J. Med. Primatol., 15: 339-349.
PubMed - Beguelini, M.R., C.C.I. Puga, S.R. Taboga and E. Morielle-Versute, 2011. Ultrastructure of spermatogenesis in the white-lined broad-nosed bat, Platyrrhinus lineatus (Chiroptera: Phyllostomidae). Micron, 42: 586-599.
CrossRefDirect Link - Holt, W.V. and H.D. Moore, 1984. Ultrastructural aspects of spermatogenesis in the common marmoset (Callithrix jacchus). J. Anat., 138: 175-188.
Direct Link - Segatelli, T.M., C.C.D. Almedia, P.F.F. Pinheiro, M. Martinez, C.R. Padovani and F.E. Martinez, 2000. Ultrastructural study of acrosomeformation in mongolian gerbil (Meriones unguiculatus). Tissue Cell, 32: 508-517.
CrossRefDirect Link - Lalli, M. and Y. Clermont, 1981. Structural changes of the head components of the rat spermatid during late spermiogenesis. Am. J. Anat., 160: 419-434.
CrossRefDirect Link - Singwi, M.S. and S.B. Lall, 1983. Spermatogenesis in the non-scrotal bat-Rhinopoma kinneari Wroughton (Microchiroptera: Mammalia). Cells Tissues Organs, 116: 136-145.
CrossRefDirect Link - Segatelli, T.M., C.C.D. Almeida, P.F.F. Pinheiro, M. Martinez, C.R. Padovani and F.E. Martinez, 2002. Kinetics of spermatogenesis in the mongolian gerbil (Meriones unguiculatus). Tissue Cell, 34: 7-13.
CrossRefPubMedDirect Link - Nakai, M., J.K. van Cleeff and J.M. Bahr, 2004. Stages and duration of spermatogenesis in the domestic ferret (Mustela putorius furo). Tissue Cell, 36: 439-446.
CrossRefDirect Link - Suphamungmee, W., C. Wanichanon, R. Vanichviriyakit and P. Sobhon, 2008. Spermiogenesis and chromatin condensation in the common tree shrew, Tupaia glis. Cell Tissue Res., 331: 687-699.
CrossRefPubMedDirect Link - Chatchavalvanich, K., A. Thongpan and M. Nakai, 2005. Ultrastructure of spermiogenesis in a freshwater stingray, Himantura signifer. Ichthyol. Res., 52: 379-385.
CrossRefDirect Link - Zhang, L., X.K. Han, M.Y. Li, H.J. Bao and Q.S. Chen, 2007. Spermiogenesis in soft-shelled turtle, Pelodiscus sinensis. Anat. Rec., 290: 1213-1222.
CrossRefDirect Link - Gunawardana, V.K. and M.G. Scott, 1977. Ultrastructural studies on the differentiation of spermatids in the domestic fowl. J. Anat., 124: 741-755.
Direct Link - Johnston, S.D., L. Daddow, F.N. Carrick and B. Jamieson, 2004. Observations of spermiogenesis and epididymal sperm maturation in the rufous hare wallaby, Lagorchestes hirsutus (Metatheria, Mammalia). Acta Zool., 85: 53-58.
CrossRefDirect Link - Lloyd, S., F. Carrick and L. Hall, 2008. Unique features of spermiogenesis in the Musky Rat-kangaroo: Reflection of a basal lineage or a distinct fertilization process? J. Anat., 212: 257-274.
CrossRefDirect Link - Afzelius, B.A., R.E. Johnsonbaugh, J.W. Kim, L. Ploen and E.M. Ritzen, 1982. Spermiogenesis and testicular spermatozoa of the olive baboon (Papio anubis). J. Submicrosc. Cytol., 14: 627-639.
PubMedDirect Link