
Abdel Dayem Zakaria
Department of Physiology, Biochemistry and Pharmacology, College of Veterinary Medicine and Animal Resources, King Faisal University, Al-Asha, P.O. Box 400, (31982), Saudi Arabia
Department of Physiology, Faculty of Veterinary Medicine, Alexandria University, Rosetta Line, Behera Province 22758, Egypt
LiveDNA: 20.14957
Aida El-Sayed Bayad
Veterinary Services Center, Faculty of Veterinary Medicine, Alexandria University, Rosetta Line, Behera Province 22758, Egypt
LiveDNA: 20.15148
Sherief M. Abdel-Raheem
Department of Veterinary Public Health and Animal Husbandry, College of Veterinary Medicine and Animal Resources, King Faisal University, Al-Asha, Saudi Arabia
Department of Animal Nutrition and Clinical Nutrition, Faculty of Veterinary Medicine, Assiut University, 71526, Egypt
LiveDNA: 20.13959
Khalid A. Al-Busadah
Department of Physiology, Biochemistry and Pharmacology, College of Veterinary Medicine and Animal Resources, King Faisal University, Al-Asha, P.O. Box 400, (31982), Saudi Arabia
Ibrahim Albokhadaim
Department of Physiology, Biochemistry and Pharmacology, College of Veterinary Medicine and Animal Resources, King Faisal University, Al-Asha, P.O. Box 400, (31982), Saudi Arabia
Asian Journal of Animal Sciences
Year: 2016 | Volume: 10 | Issue: 2 | Page No.: 139-146







ABSTRACT
The present investigation was conducted to examine the role of camel’s milk on semen characteristic in immobilization stressed rats. The rats were divided into four groups; control (untreated), camel’s milk treated, immobilization stressed and camel’s milk treated immobilization stressed group. Immobilization stress resulted in significant decrease in both body and reproductive organs weight, sperm count, alive sperm percentage, mass motility percentage, plasma testosterone level and antioxidative stress parameters, while, it caused significant increase in adrenal gland weight, sperm abnormalities percentage, plasma glucose, corticosterone and malondialdehyde levels. Administration of camel’s milk to rats exposed to immobilization stress significantly amended the estimated parameters although not all were similar to control levels. It could be concluded from the present study that camel’s milk administration before immobilization stress improves semen characteristics in rats.
PDF Abstract XML References Citation
Received: December 09, 2015;
Accepted: January 05, 2016;
Published: February 15, 2016
How to cite this article
Abdel Dayem Zakaria, Aida El-Sayed Bayad, Sherief M. Abdel-Raheem, Khalid A. Al-Busadah and Ibrahim Albokhadaim, 2016. Camel’s Milk Improves the Semen Characteristic in Immobilization Stressed Rats. Asian Journal of Animal Sciences, 10: 139-146.
DOI: 10.3923/ajas.2016.139.146
URL: https://scialert.net/abstract/?doi=ajas.2016.139.146
DOI: 10.3923/ajas.2016.139.146
URL: https://scialert.net/abstract/?doi=ajas.2016.139.146
INTRODUCTION
Day by day, the stress is increasing in our life and it is responsible for calamitous results on the health, life quality and broad prevalence reason of disease. Steinberger (1978) stated that many forms of stress including physical and psychological can affect male fertility and reproduction. The stressor stimulates of the hypothalamo-hypophyseal-adrenal axis is a supposable mechanism for the suppression of male reproductive tasks out of inhibition in the hypothalamic-pituitary-testicular axis (Rivier and Rivest, 1991). Acute and chronic immobilization attained in studies of experimental stress can decrease testosterone secretion in adult rats (Demura et al., 1989; Orr and Mann, 1992). Exposure to stress is known to change the prooxidant-antioxidant equilibrium, which might lead to the expansion of different pathological cases. The restraint stress model has been shown to produce robust increases in basal oxidative stress (Zafir and Banu, 2009). Induction of restraint stress to rats was shown to cause oxidative stress which was accompanied with significantly reduced activities of antioxidant enzymes; SOD, GST, CAT and GR (Khataibeh, 2013) and increased Reactive Oxygen Species (ROS) e.g., hydrogen peroxide, superoxide anion radical and hydroxyl radical that result peroxidation of the lipid, particularly in membranes and can perform an substantial role in tissue injury. It has been proposed that chronic stress and elevation of adrenal glucocorticoids, which secreted during stress, affect various processes including ROS and elevate ROS by about 10% basically (Kovacs et al., 1996). The membrane injury produces distubrance of the tissue (Bagchi et al., 1999; Cochrane, 1991; McIntosh and SapoIsky, 1996).
Oxidative stress induces the range of pathologies that may be affect the reproductive function (Maneesh et al., 2005). The generated ROS has a relationship with infertility (Shiraishi and Naito, 2007; Agarwal et al., 2008). Great attention have been paid to generate ROS because they have toxic action on sperm function and quality when they are generated at high level (Padron et al., 1997). The spermatozoa plasma membrane contains large amounts of polyunsaturated fatty acids so it is very susceptible to OS-induced damage (Alvarez and Storey, 1995) and the scavenging enzymes concentrations are low at their cytoplasm (Sharma and Agarwal, 1996). The cell membrane polyunsaturated fatty acids are the most target of the ROS leading to lipid oxidation or lipid peroxidation (Makker et al., 2009). Aldehydes such as malondialdehyde (MDA) are the results of lipid peroxides degeneration which in turn reacting with lipids, proteins and nucleic acids causing damage of the cell (Sim et al., 2003). In addition, because the shortage of intracellular antioxidant enzymes that give limited protection to the cell membrane the sperm depend on the protection afforded by the seminal plasma (Zini et al., 1993). There are inverse relationship between the ROS levels and the sperm motility (Iwasaki and Gagnon, 1992).
Camel’s milk is diverse from other ruminant milk it contains high minerals; vitamins and insulin contents but have low protein; cholesterol and sugar (Knoess, 1979). This may give it its medicinal properties. In Sahara, butter of fresh camel’s milk is overwhelmingly used as a base for medicines. Cosmetics or pharmaceuticals can be develop from camel’s milk. Camel’s milk can treated a series of metabolic and autoimmune diseases. Chronic pulmonary tuberculosis can be treated with raw camel’s milk (Agarwal et al., 2008). Agarwal et al. (2008) found that the required daily dose of insulin for diabetic patients with type 1, reduced by 30-35% in response to treatment with camel’s milk.
The unrivaled constituent of camel’s milk has been evidenced to trench the immunity. The immunity components have manifested to be efficient in destructing microbial agents, both bacterial and viral and act as safeguard to the human body against many diseases and treats several types of cancer.
Camel’s milk reduced free radicals production and oxidative stress status in cadmium induced anemia in rats (Dallak, 2009) and toxicity in rats (Al-Hashem, 2009a). Oral administration of camel’s milk before administration of aluminum chloride alleviates lipid peroxidation and oxidative stress in the red blood cells (Al-Hashem, 2009a) and rat testes (Al-Hashem, 2009b) liver and kidney of rat (Al-Hashem, 2009a). Camel’s milk administration to diabetic patients improved the diabetes-induced oxidative stress (El-Said et al., 2010). Moreover, camel’s milk lessen injury induced by alcohol in rat’s liver and protects hepatic tissue from alcohol-induced toxicity (Darwish et al., 2012).
In our mopping, we didn’t meet any research study the effect of camel’s milk against immobilization stress. Hence, in the present study, the conservative efficiency of camel’s milk was assessed against the deleterious effect induced by immobilization stress on semen characteristics.
MATERIALS AND METHODS
Milk sample: Early in the morning, camel’s milk samples were collected daily from Experimental Veterinary and Agriculture Station belonging to King Faisal University, at AL Asha, Saudi Arabia. Milk was collected by hand milking from camels .The samples were collected aseptically in sterile screw-capped plastic containers, placed in a cool box containing ice packs and immediately transported to the laboratory.
Experimental animals and protocol: Twenty four adult male Wister rats, weighing 200-250 g were obtained from the Experimental Veterinary and Agriculture Station belonging to King Faisal University, at AL Asha, Saudi Arabia, housed in the Physiology Laboratory in plastic cages. The rats were adapted in controlled environment (temperature: 20-25°C with equal of light and dark). The rats were fed with standard food pellets and water ad libitum, proximate analysis of food pellets performed according to the guidelines of AOAC (2005), is presented in Table 1. In this study, the experiment conducted on the animals were in accordance with King Faisal University’s guidance from the Ethical Committee for Research on Laboratory Animals. After adaptation for 1 week, the animal were divided randomly into 4 groups (6 rats each) as the following. (I) Unstressed group (Control): The animals in this group were unstressed and they received intra-gastric 2 mL saline daily (using stomach tube), (2) Camel’s milk group: Consisted of unstressed rats and they received daily intra-gastric 2 mL camel’s milk (using stomach tube), (3) Immobilization-stressed group: the animal in this group were subjected to immobilization stress for 6 h/day, 6 days/week (Almeida et al., 1998) and were daily administered intra-gastric 2 mL saline before exposure to immobilization stress by 30 min and (4) Camel’s milk-treated immobilization-stressed group: The rats in this group were daily administered intra-gastric 2 mL camel’s milk before exposure to immobilization stress by 30 min. The restraint device was a transparent plastic tube, the animal faced the apex of the tube, which had an opening 1 cm in diameter approximately 1 cm from the animal’s nose and the tube was closed at the base with the tape. The size of the tube sufficient to induce stress without unnecessary pain. To avoid familiarization of animals to the daily procedure, immobilization times were arbitrarily rotated within certain time periods. The experiment was conducted for 8 weeks to cover complete spermatogenic cycle (Clermont and Harvey, 1965).
Body and reproductive organ weight: All rats were weighed individually twice at the beginning and at the end of the experiment, sacrificed by decapitation immediately after the end of the last immobilization stress session. Immediately after blood collection, a midline abdominal incision was made. The testes; epididymis; prostate and seminal vesicles glands were dissected free, blot dry and weighed. The Index Weight (IW) was estimated for each organ according to Matousek (1969).
| Table 1: | Chemical analysis of the basal diet |
 | |
Biochemical analysis and hormonal assay: Fresh plasma were used for glucose estimation enzymatically as described by Trinder (1969) using Glucofix kits (Menarine Diagnostic, Italy), testosterone determination was done according to the method described by Orr and Mann (1992) using RIA kits from Diagnostic Products Cooperation (Los Angeles, California) with a sensitivity of 0.2 mg mL–1 and intra-assay variation coefficient was 12.8%. Corticosterone level was measured according to Ortiz et al. (1999) using RIA kit (ICN Biomedicals, Costa Mesa, CA). The inter and intrassay coefficients of variation were 3.6 and 4.3%, respectively. The plasma was used for determination of GSH (Sedlak and Lindsay, 1968), SOD (Nishikimi et al., 1972) and CAT (Aebi, 1984), MDA (Ohkawa et al., 1979) using commercial available colorimetric assay kits (Bio diagnostic, Egypt) according to the manufacturer guides.
Sperm concentration: Epididymal spermatozoa concentration were estimated by some modification of Yokoi et al. (2003) method. The epididymis was cut into head; body and tail, then they minced in 5 mL phosphate buffer (pH 7.4), after that they shake vigorously for homogeneity and dispersal of sperm cells. An aliquot (10 μL) of epididymal sperm suspension was placed in the hemocytometer and was pliable to stand for 5 min to be count by 200× magnification of light microscope. The heads of sperm were estimated and expressed as million per milliliter.
Alive sperm percent: One drop of epididymal content of each animal was mixed with one drop of eosin-nigrosin stain and a thin film was made on a clean slide. The average viability percent was determined from 200 sperms examined per slide (Bearden and Fuquary, 1980).
Sperm motility: The progressive motility was evaluated according to Sonmez et al. (2005). The content of cauda epididymis was obtained with a pipette and 2 mL tris buffer solution was added to it. The average final motility score was estimated from the three different fields in each sample. The motility percentage was determined at 400× magnification.
Abnormality percentage: The epididymal sperm morphology was determined according to Evans and Maxwell (1987). Briefly, 40 μL of sperm suspension was mixed with 10 μL of 1% eosin and nigrosine, 200 sperm were examined on each slide using a light microscope at 400× magnification and the percentages of abnormal spermatozoa morphology were recorded.
Statistical analysis: The obtained data were expressed as means±standard errors. The significance of differences between means was calculated by one-way analysis of variance followed by Duncan’s multiple range test (SAS., 2001). The difference between means was considered significant when p<0.05.
RESULTS
Body weight and food intake: Table 2 showed that the body weights and food intake significantly decreased in immobilization-stressed and camel’s milk-treated immobilization-stressed groups compared to that of control group (p≤0.05). The food intake significantly decrease in immobilization-stressed compared to camel’s milk-treated immobilization-stressed group.
Reproductive organ and adrenal gland weights: As observed in Table 2, the index weight of the epididymis, testes, prostate and seminal vesicle significantly decreased (p≤0.05) in immobilized stressed rats and those immobilized stressed and treated with camel’s milk compared to the control group. Also, the index weight of the epididymis, testes, prostate and seminal vesicle significantly decreased (p≤0.05) in immobilized stressed rats in comparison with those of immobilized stressed treated with camel’s milk. The adrenal gland index weight increased significantly (p≤0.05) in immobilized stressed rats and those immobilized stressed and treated with camel’s milk compared to the control. The adrenal gland index weight significantly increased in immobilized stressed rats compared with that of immobilized stressed and treated with camel’s milk.
Epididymal sperm characteristics: Immobilization stress and immobilization stress plus camel’s milk caused significant reduction (p≤0.05) in epididymal sperm concentration; alive sperm and mass motility percentages, while increase sperm abnormalities percent liken to the control. The decrease was the minimal in immobilization stressed rats treated with camel’s milk (Table 3).
Biochemical and hormonal analysis: Table 4 shows the plasma CAT, SOD and GSH and testosterone (Table 5) levels were significantly decreased (p≤0.05) in immobilization stressed and the immobilization stressed plus camel’s milk treated groups in comparison with the control group. The decline was less obvious in the immobilization stress plus camel’s milk treated group.
| Table 2: | Effect of immobilization stress and immobilization stress plus camel’s milk on body weight, sex organs and adrenal gland index weight |
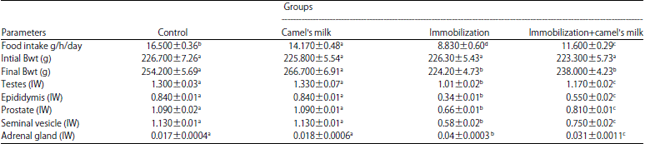 | |
Mean±SE, IW: Index weight, N = 6, immobilization was done for 6 h daily for 8 weeks, camel’s milk was administered intra gastric 30 min before immobilization, means followed by similar letter are not significantly different at p≤0.05 | |
| Table 3: | Effect of immobilization stress and immobilization stress plus camel’s milk on epididymal sperm characteristics |
 | |
Means±SE, N = 6, immobilization was done for 6 h daily for 8 weeks, camel’s milk was administered intra gastric 30 min before immobilization, means followed by similar letter are not significantly different at p≤0.05 | |
| Table 4: | Effect of immobilization stress and camel’s milk on plasma oxidative stress parameter levels |
 | |
Mean±SE, N = 6, immobilization was done for 6 h daily for 8 weeks, camel’s milk was administered intra gastric 30 min before immobilization, means followed by similar letter are not significantly different at p≤0.05, MDA: Malondialdehyde, CAT: Catalase, SOD: Superoxide dismutase, GSH: Glutathione | |
| Table 5: | Effect of immobilization stress and camel’s milk on plasma glucose, testosteroneand corticosterone levels |
 | |
Means±SE, N = 6, immobilization was done for 6 h daily for 8 weeks, camel’s milk was administered intra gastric 30 min before immobilization, means followed by similar letter are not significantly different at p≤0.05 | |
Furthermore, plasma glucose, corticosterone (Table 5) and MDA (Table 4) increased significantly (p≤0.05) in the immobilization stress and the immobilization stress plus camel’s milk treated groups compared to the control. Moreover, (Table 4 and 5) shows that the greatest increase in plasma glucose, corticosterone and MDA was observed in immobilization stress rats.
DISCUSSION
In the present study immobilization stress significantly decreased the body weight and food intake as compared to control group, this effect agree with other reports (Herman et al., 1995; Nayanatara et al., 2012) which have found that weight loss has occurred as a result of stress. The weight loss is explained as a result of reduced feed and water consumption or exhaustion of body reserves as a result of increased metabolic activity (Herman et al., 1995). Suppressed appetite in repeated stress might be due to adrenocorticotrophic releasing hormone which is released during stress (Nayanatara et al., 2005). In the present study the adrenal gland weight and corticosterone were increased in the immobilization stressed group, one of the of assessment of the initial responses of the organisms to stress of any kind is hypertrophy and hyperplasia of the adrenal gland (Alario et al., 1987; Marti et al., 1993). The release of adrenaline and glucocorticoid from the adrenal gland begins the biological reactions allowing the organism to overcome the adverse physiological and psychological stressor (Wong, 2006).
In our study, chronic immobilization resulted in a significant decrease in the reproductive organs weight which could be attributed to decrease in plasma testosterone levels. These results are in line with the results of Rai et al. (2003) who concluded that the immobilization stress causes reduction in testes size and weight and atrophy of the seminal vesicles and prostate gland. The mass of differentiated spermatogenic cells is the main factor affect the testes weight, the decrease in the testes weight could be attributed to inhibition of steroidogenic enzyme activity; spermatogenesis and reduction of the number of germ cells (Takahashi and Oishi, 2001). Our results revealed that immobilized stress rats had diminished sperm quality (Kini et al., 2009). The decrease in sperm count could be attributed to the bad effect of immobilization stress on spermatogenesis. The stress induced lipid peroxidation or decrease in serum testosterone levels which could be the mechanisms by which the stress induced its harmful effect of on spermatogenesis. The changes in epididymal weights was consistent with the reduction in epididymal sperm count. In males, one of the first signs of stress is the decrease of serum testosterone (Fenster et al., 1997). Spermatogenesis and maintenance of the histology and function of the accessory sex glands in the male need testosterone (Boockfor and Blake, 1997). Serum testosterone level significantly reduced in immobilized stress rats and that lipid peroxidation induced by stress (Liu et al., 1996). The increased lipid peroxidation increase the alteration of sperm membrane functions, impair development of sperm and reduced sperm motility beside oxidative damage to sperm DNA (Aitken et al., 1989). In the present study, the highest concentration of MDA was observed in immobilized stress rats which could be lead to loss of membrane structure and function. These results were similar to those of Ahmed (2012).
It was found that immobilized stress rats demonstrated increase in the plasma glucose level. These findings are in line with the study of Thomas et al. (2002) and Ahmed (2012) who observed a significant inverse correlation between fasting blood glucose and testosterone level. Also, Muthusamy et al. (2009) found that the level of glucose was increased and the serum insulin and glucose uptake decreased after castration and administration of physiological doses of testosterone reverse the situation. Many studies on animals and human stated that hyperglycemia is associated with stress (Surwit et al., 1992). Plasma testosterone in the current work declined upon exposure to immobilization stress and this result is in line with the previous studies (Norman and Smith, 1992). Testosterone administration to immobilization-stressed rats resulted in decrease in blood glucose level (Ahmed, 2012).
Immobilization stress in the present study, led to increase of MDA and decreased the GSH, CAT and SOD levels, which suggests that the increased lipid peroxidation and per oxidative damage resulted from stress protocol. Oxidative stress and the changes in biochemical parameters produced in different organs of the rats can be corrected by camel’s milk administration (Al-Hashem, 2009a). In this study, feeding camel’s milk prior to immobilization stress resulted in significant decrease in MDA and glucose levels; increase GSH, SOD and CAT, improve in semen characteristics and increase body and reproductive organ weights compared to rats exposed to immobilization stress alone. Camel’s milk has antioxidant properties since it contains several antioxidant vitamins in high concentrations such as vitamin B2, C, E and A and very rich in many trace elements e.g., magnesium and zinc (Yousef, 2004). Vitamin E in camel’s milk acts as antioxidant molecules and as free radical scavenger, also it is necessary for normal activity of oxidative enzymes (Palamanda and Kehrer, 1993). In addition, vitamin C in camel’s milk scavengers superoxide, H2O2 and hydroxyl radicals so it prevents agglutination of sperm, also it helps in vitamin E recycle and prevents lipid peroxidation and protects against DNA damage induced by H2O2 radical (Guney et al., 2007). Magnesium in camel’s milk helps in the absorption and metabolism of different vitamins e.g., C, B and E and plays as antioxidant (Barbagallo et al., 1999). Additionally, another essential trace element for the activity of large number of enzymes and for the living organisms and found in large quantity in camel’s milk is Zinc (Yousef, 2004). Notably, Zn can block cellular deterioration by antioxidant system activation (Powell, 2000; Ozturk et al., 2003; Ozdemir and Inanc, 2005). Sperm cells are metabolically active and generate large numbers of free radicals during their development. Zinc dependent enzyme systems help neutralize these free radicals thereby improving sperm quality. Inferior semen quality, idiopathic male infertility and improper development of secondary sex characters and hypogonadism associated accompanied with zinc deficiency and low zinc concentration in the semen (Endre et al., 1990; Prasad, 1991; Haas, 2006; Colagar et al., 2009).
CONCLUSION
It could be concluded from the present study that camel’s milk administration to rats before exposed to immobilization stress corrected the estimated parameters and semen characteristics although they not reach to the control levels.
ACKNOWLEDGMENTS
The researchers are thankful to the Camel Research Centre, King Faisal University, especially Professor Dr. M. M. Al-Eknah for his support and help during the period this research took place and for enabling us to obtain camel’s milk.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Agarwal, A., K. Makker and R. Sharma, 2008. Clinical relevance of oxidative stress in male factor infertility: An update. Am. J. Reprod. Immunol., 59: 2-11.
CrossRefPubMedDirect Link - Ahmed, M.A., 2012. Role of testosterone in glucose homeostasis in immobilization stressed rats. J. Am. Sci., 8: 628-637.
Direct Link - Aitken, R.J., J.S. Clarkson and S. Fishel, 1989. Generation of reactive oxygen species, lipid peroxidation and human sperm function. Biol. Reprod., 41: 183-187.
Direct Link - Al-Hashem, F.H., 2009. Camel's milk protects against aluminum chloride-induced toxicity in the liver and kidney of white albino rats. Am. J. Biochem. Biotehcnol., 5: 98-108.
CrossRefDirect Link - Al-Hashem, F.H., 2009. Camel's milk alleviates oxidative stress and lipid peroxidation induced by chronic aluminum chloride exposure in rat's testes. Am. J. Applied Sci., 6: 1868-1875.
CrossRefDirect Link - Almeida, S.A., S.O. Petenusci, J.A. Anselmo-Franci, A.A. Rosa-e-Silva and T.L. Lamano-Carvalho, 1998. Decreased spermatogenic and androgenic testicular functions in adult rats submitted to immobilization-induced stress from prepuberty. Braz. J. Med. Biol. Res., 31: 1443-1448.
PubMedDirect Link - Alario, P., A. Gamallo, M.J. Beato and G. Trancho, 1987. Body weight gain, food intake and adrenal development in chronic noise stressed rats. Physiol. Behav., 40: 29-32.
CrossRefPubMedDirect Link - Alvarez, J.G. and B.T. Storey, 1995. Differential incorporation of fatty acids into and peroxidative loss of fatty acids from phospholipids of human spermatozoa. Mol. Reprod. Dev., 42: 334-346.
PubMedDirect Link - Barbagallo, M., L.J. Dominguez, M.R. Tagliamonte, L.M. Resnick and G. Paolisso, 1999. Effects of vitamin E and glutathione on glucose metabolism: Role of magnesium. Hypertension, 34: 1002-1006.
CrossRefPubMedDirect Link - Bagchi, D., O.R. Carryl, M.X. Tran, M. Bagchi and A. Garg et al., 1999. Acute and chronic stress-induced oxidative gastrointestinal mucosal injury in rats and protection by bismuth subsalicylate. Mol. Cell Biochem., 196: 109-116.
PubMedDirect Link - Boockfor, F.R. and C.A. Blake, 1997. Chronic administration of 4-tert-octylphenol to adult male rats causes shrinkage of the testes and male accessory sex organs, disrupts spermatogenesis and increases the incidence of sperm deformities. Biol. Reprod., 57: 267-277.
Direct Link - Clermont, Y. and S.C. Harvey, 1965. Duration of the cycle of the seminiferous epithelium of normal, hypophysectomized and hypophysectomized-hormone treated albino rats. Endocrinology, 79: 80-89.
PubMedDirect Link - Cochrane, C.G., 1991. Mechanisms of oxidant injury of cells. Mol. Aspects Med., 12: 137-147.
CrossRefDirect Link - Colagar, A.H., E.T. Marzony and M.J. Chaichi, 2009. Zinc levels in seminal plasma are associated with sperm quality in fertile and infertile men. Nutr. Res., 29: 82-88.
CrossRefDirect Link - Dallak, M., 2009. Camel's milk protects against cadmium chloride-induced hypochromic microcytic anemia and oxidative stress in red blood cells of white albino rats. Am. J. Pharmacol. Toxicol., 4: 136-143.
Direct Link - Darwish, H.A., N.R. Abd Raboh and A. Mahdy, 2012. Camel's milk alleviates alcohol-induced liver injury in rats. Food Chem. Toxicol., 50: 1377-1383.
CrossRefPubMedDirect Link - Demura, R., T. Suzuki, S. Nakamura, H. Komatsu, E. Odagiri and H. Demura, 1989. Effect of immobilization stress on testosterone and inhibin in male rats. J. Androl., 10: 210-213.
CrossRefPubMedDirect Link - El-Said, E.E., G.R. El-Sayed and E. Tantawy, 2010. Effect of camel milk on oxidative stresses in experimentally induced diabetic rabbits. Vet. Res. Forum, 1: 30-43.
Direct Link - Fenster, L., D.F. Katz, A.J. Wyrobek, C. Pieper, D.M. Rempel, D. Oman and S.H. Swan, 1997. Effects of psychological stress on human semen quality. J. Androl., 18: 194-202.
Direct Link - Guney, M., B. Oral, H. Demirin, N. Karahan, T. Mungan and N. Delibas, 2007. Protective effects of vitamins C and E against endometrial damage and oxidative stress in fluoride intoxication. Clin. Exp. Pharmacol. Physiol., 34: 467-474.
CrossRefDirect Link - Herman, J.P., D. Adams and C. Prewitt, 1995. Regulatory changes in neuroendocrine stress-integrative circuitry produced by a variable stress paradigm. Neuroendocrinology, 61: 180-190.
PubMedDirect Link - Iwasaki, A. and C. Gagnon, 1992. Formation of reactive oxygen species in spermatozoa of infertile patients. Fertil. Steril., 57: 409-416.
PubMedDirect Link - Rai, J., S.N. Pandey and R.K. Srivastava, 2003. Effect of immobilization stress on spermatogenesis of albino rats. J. Anat. Soc. India, 52: 52-57.
Direct Link - Khataibeh, M., 2013. Cinnamon modulates biochemical alterations in rats loaded with acute restraint stress. J. Saudi Chem. Soc.
CrossRef - Kini, R.D., A.K. Nayanatara, S.R. Pai, C. Ramswamy, M.R. Bhat and V.S. Mantur, 2009. Infertility in male wistar rats induced by cadmium chloride: Role of ascorbic acid. J. Chinese Clin. Med., 4: 616-621.
Direct Link - Kovacs, P., I. Juranek, T. Stankovicova and P. Svec, 1996. Lipid peroxidation during acute stress. Die Pharmazie, 51: 51-53.
PubMedDirect Link - Makker, K., A. Agarwal and R. Sharma, 2009. Oxidative stress and male infertility. Indian J. Med. Res., 129: 357-367.
PubMed - Maneesh, M., H. Jayalekshmi, S. Dutta, A. Chakrabarti, D.M. Vasudevan, 2005. Effect of chronic ethanol administration on testicular antioxidant system and steroidogenic enzyme activity in rats. Indian J. Exp. Biol., 43: 445-449.
PubMedDirect Link - Marti, O., A. Gavalda, T. Jolin and A. Armario, 1993. Effect of regularity of exposure to chronic immobilization stress on the circadian pattern of pituitary adrenal hormones, growth hormone and thyroid stimulating hormone in the adult male rat. Psychoneuroendocrinology, 18: 67-77.
CrossRefDirect Link - Matousek, J., 1969. Effects on spermatogenesis in guinea-pigs, rabbits and sheep after their immunization with sexual organ fluids of bulls. J. Reprod. Fertil., 19: 63-72.
CrossRefDirect Link - McIntosh, L.J. and R.M. Sapolsky, 1996. Glucocorticoids increase the accumulation of reactive oxygen species and enhance adriamycin-induced toxicity in neuronal culture. Exp. Neurol., 141: 201-206.
CrossRefDirect Link - Muthusamy, T., P. Murugesan and K. Balasubramanian, 2009. Sex steroids deficiency impairs glucose transporter 4 expression and its translocation through defective Akt phosphorylation in target tissues of adult male rat. Metabolism, 58: 1581-1592.
CrossRefDirect Link - Nayanatara, A.K., H.S. Nagaraja and B.K. Anupama, 2005. The effect of repeated swimming stress on organ weights and lipid peroxidation in rats. Thai. J. Pharm. Sci., 18: 3-9.
Direct Link - Nayanatara, A.K., Y. Tripathi, H.S. Nagaraja, P.S. Jeganathan, C. Ramaswamy, B. Ganaraja and S.R. Pai, 2012. Effect of chronic immobilization stress on some selected physiological, biochemical and lipid parameters in Wistar albino rats. Res. J. Pharm. Biol. Chem. Sci., 3: 34-42.
Direct Link - Nishikimi, M., N.A. Rao and K. Yagi, 1972. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun., 46: 849-854.
CrossRefPubMedDirect Link - Norman, R.L. and C.J. Smith, 1992. Restraint inhibits luteinizing hormone and testosterone secretion in intact male rhesus macaques: Effects of concurrent naloxone administration. Neuroendocrinology, 55: 405-415.
CrossRefDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Orr, T.E. and D.R. Mann, 1992. Role of glucocorticoids in the stress-induced suppression of testicular steroidogenesis in adult male rats. Hormone Behav., 26: 350-363.
CrossRefPubMedDirect Link - Ortiz, R.M., T. Wang and C.E. Wade, 1999. Influence of centrifugation and hindlimb suspension on testosterone and corticosterone excretion in rats. Aviation Space Environ. Med., 70: 499-504.
PubMedDirect Link - Ozdemir, G. and F. Inanc, 2005. Zinc may protect remote ocular injury caused by intestinal ischemia reperfusion in rats. Tohoku J. Exp. Med., 206: 247-251.
CrossRefPubMedDirect Link - Ozturk, A., A.K. Baltaci, R. Mogulkoc, E. Oztekin, A. Sivrikaya, E. Kurtoglu and A. Kul, 2003. Effects of zinc deficiency and supplementation on malondialdehyde and glutathione levels in blood and tissues of rats performing swimming exercise. Biol. Trace Elem. Res., 94: 157-166.
CrossRefPubMedDirect Link - Padron, O.F., N.L. Brackett, R.K. Sharma, C.M. Lynne, A.J. Thomas and A. Agarwal, 1997. Seminal reactive oxygen species and sperm motility and morphology in men with spinal cord injury. Fertil. Steril., 67: 1115-1120.
CrossRefDirect Link - Palamanda, J.R. and J.P. Kehrer, 1993. Involvement of vitamin E and protein thiols in the inhibition of microsomal lipid peroxidation by glutathione. Lipids, 28: 427-431.
CrossRefDirect Link - Powell, S.R., 2000. The antioxidant properties of zinc. J. Nutr., 130: 1447S-1454S.
CrossRefDirect Link - Prasad, A.S., 1991. Discovery of human zinc deficiency and studies in an experimental human model. Am. J. Clin. Nutr., 53: 403-412.
Direct Link - Rivier, C. and S. Rivest, 1991. Effect of stress on the activity of the hypothalamic-pituitary-gonadal axis: Peripheral and central mechanisms. Biol. Reprod., 45: 523-532.
CrossRefDirect Link - Sedlak, J. and R.H. Lindsay, 1968. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal. Biochem., 25: 192-205.
CrossRefPubMedDirect Link - Sharma, R.K. and A. Agarwal, 1996. Role of reactive oxygen species in male infertility. Urology, 48: 835-850.
CrossRefPubMedDirect Link - Shiraishi, K. and K. Naito, 2007. Effects of 4-hydroxy-2-nonenal, a marker of oxidative stress, on spermatogenesis and expression of p53 protein in male infertility. J. Urol., 178: 1012-1017.
CrossRefDirect Link - Sim, A.S., C. Salonikas, D. Naidoo and D.E.L. Wilcken, 2003. Improved method for plasma malondialdehyde measurement by high-performance liquid chromatography using methyl malondialdehyde as an internal standard. J. Chromatography B, 785: 337-344.
CrossRefDirect Link - Sonmez, M., G. Turk and A. Yuce, 2005. The effect of ascorbic acid supplementation on sperm quality, lipid peroxidation and testosterone levels of male Wistar rats. Theriogenology, 63: 2063-2072.
CrossRefDirect Link - Steinberger, E., 1978. The etiology and pathophysiology of testicular dysfunction in man. Fertil. Steril., 29: 481-491.
PubMed - Surwit, R.S., M.S. Schneider and M.N. Feinglos, 1992. Stress and diabetes mellitus. Diabetes Care, 15: 1413-1422.
PubMedDirect Link - Takahashi, O. and S. Oishi, 2001. Testicular toxicity of dietary 2,2-bis(4-hydroxyphenyl)propane (bisphenol A) in F344 rats. Arch. Toxicol., 75: 42-51.
CrossRefDirect Link - Thomas, J.L., B.T. Quang, E. Ochsenbein and J.P. Vincent, 2002. Relationship between some biological parameters and plasma testosterone in institutionalized aging men. J. Nutr. Health Aging, 7: 437-439.
PubMedDirect Link - Trinder, P., 1969. Determination of blood glucose using 4-amino phenazone as oxygen acceptor. J. Clin. Pathol., 22: 246-246.
Direct Link - Wong, D.L., 2006. Epinephrine biosynthesis: Hormonal and neural control during stress. Cell. Mol. Neurobiol., 26: 889-898.
CrossRefDirect Link - Yokoi, K., E.O. Uthus and F.H. Nielsen, 2003. Nickel deficiency diminishes sperm quantity and movement in rats. Biol. Trace Elem. Res., 93: 141-154.
CrossRefPubMedDirect Link - Yousef, M.I., 2004. Aluminum-induced changes in hemato-biochemical parameters, lipid peroxidation and enzyme activities of male rabbits: Protective role of ascorbic acid. Toxicology, 199: 47-57.
CrossRefDirect Link - Zafir, A. and N. Banu, 2009. Induction of oxidative stress by restraint stress and corticosterone treatments in rats. Indian J. Biochem. Biophys., 46: 53-58.
PubMedDirect Link - Zini, A., E. de Laraminde and C. Gagnon, 1993. Reactive oxygen species in semen of infertile patients: Levels of superoxide dismutase-and catalase-like activities in seminal plasma and spermatozoa. Int. J. Androl., 16: 183-188.
CrossRefPubMedDirect Link