
E.V. Ikpeme
Department of Genetics and Biotechnology, University of Calabar, PMB1115, Calabar, Nigeria
O.U. Udensi
Department of Genetics and Biotechnology, University of Calabar, PMB1115, Calabar, Nigeria
LiveDNA: 234.3756
M.C. Okolo
Department of Genetics and Biotechnology, University of Calabar, PMB1115, Calabar, Nigeria
F.U. Ogban
Department of Computer Science, University of Calabar, PMB1115, Calabar, Nigeria
N.G. Ufford
Department of Genetics and Biotechnology, University of Calabar, PMB1115, Calabar, Nigeria
E.U. Odo
Department of Genetics and Biotechnology, University of Calabar, PMB1115, Calabar, Nigeria
B.O. Asuquo
Department of Animal Science, University of Calabar, PMB1115, Calabar, Nigeria
Asian Journal of Animal Sciences
Year: 2016 | Volume: 10 | Issue: 2 | Page No.: 131-138







ABSTRACT
Genetic diversity in germplasm is cardinal for breeders as it provides potential genotype (s) for breeding and improvement. Sixty C. gariepinus samples were obtained from the wild and cultured populations in Cross River, Nigeria. Twenty one morphometric traits were taken from each fish sample and were later prepared for proximate and mineral composition analyses. Predictive Analytics Software (PASW) version 20.0 was used for data analyses. Results showed that the standard length, pre-dorsal and pre-anal distances, pre-pectoral distance, head length, head width, eye diameter, distance between occipital process and caudal fin of C. garipeinus from the wild populations were significantly higher than those from the cultured populations. Calcium and magnesium contents were high in C. garipeinus from the wild populations while protein content was higher in the cultured C. garipeinus. The PC1 and PC2 contributed 44.89 and 14.00% to the total variability of 84.17%. From PC1, standard length (0.924), pre-dorsal distance (0.856), pre-anal distance (0.941), dorsal fin length (0.890), anal fin length (0.839), head length (0.946), head width (0.863), inter-orbital distance (0.820) and eye diameter (0.896) contributed significantly to the total variability observed in C. gariepinus populations. Cluster analysis revealed two major clusters for both morphometric and proximate composition, which were largely population-dependent. These suggest that C. gariepinus breeders and farmers should source C. gariepinus species from the wild in order to genetically enrich the gene pool. Additionally, head region-based morphometric traits might be more informative in distinguishing wild and cultured populations of C. gariepinus.
PDF Abstract XML References Citation
Received: September 18, 2015;
Accepted: December 19, 2015;
Published: February 15, 2016
How to cite this article
E.V. Ikpeme, O.U. Udensi, M.C. Okolo, F.U. Ogban, N.G. Ufford, E.U. Odo and B.O. Asuquo, 2016. Genetic Relatedness of Clarias gariepinus (L.) from Cultured and Wild Populations Using Multivariate Analyses. Asian Journal of Animal Sciences, 10: 131-138.
DOI: 10.3923/ajas.2016.131.138
URL: https://scialert.net/abstract/?doi=ajas.2016.131.138
DOI: 10.3923/ajas.2016.131.138
URL: https://scialert.net/abstract/?doi=ajas.2016.131.138
INTRODUCTION
Clarias gariepinus (African Catfish) is a fresh water species that belongs to the family Clarridae (order: Siluriformes) and consists 14 genera and 92 species (Teugels, 1986). This family of catfish is distributed all over Africa and Southeast Asia (Skelton, 2001). Clarias gariepinus species can be recognized by their scales, elongated body, long dorsal and anal fins and flattened bony head. They also possess a terminal broad mouth with four pairs of barbell and a large accessory breathing organ (Heok-Hee, 2003). Dorsally, they exhibit varying colours from dark to light brown often mottled with shades of grey and olive and pale cream to white underside (Skelton, 2001), which might be purely due to environmental variations.
Reduction in abundance of this catfish species in the natural water bodies has been reported (Islam et al., 2012; Garg et al., 2009), which might be due to overfishing, aquatic pollution, habitat degradation and other anthropogenic activities (King, 2007; Asuqwo and Udoh, 2002; Nwafili et al., 2012). This reduction in population size may lead to decreased genetic variation and loss of biological potentials of a stock (Islam et al., 2012) with the implication of creating genetic erosion. Unfortunately, majority of fish needed to meet consumers’ demands are obtained from the natural water bodies (Haruna et al., 2006) that has been reported to have lost its naturality with the resultant effect on genetic diversity. In Nigeria, C. gariepinus has gained popularity and attracted the interest of the aqua-culturists because of its high resistance to diseases, fast growth rate, high fecundity, palatability, high stocking densities under culture conditions and ability to tolerate a wide range of environmental conditions (Eyo et al., 2012; Olubunmi et al., 2009; Babalola and Apata, 2006). This notwithstanding, the cultured catfish populations is faced with series of challenges such as inbreeding depression, founder effects, genetic drift (Van Der Walt et al., 1993) that reduces genetic diversity. Genetic diversity in a germplasm is very cardinal and strategic to breeders as the greater the genetic diversity, the better for breeding and improvement.
Morphometric analysis has proven to be useful in species, races and population differentiation and has been widely employed in identification of different fish stock (Turan et al., 2004, 2005) for breeding and genetic manipulation purposes. Fishes exhibit high phenotypic plasticity and quickly adapt themselves to environmental changes by changing certain morphometric traits (Murta, 2000). Therefore, knowledge of the biology and population structure of any species according to Turan et al. (2006) is important for developing management and conservation plans of that species and may be used to study short term environmentally induced variations.
Presently, there is paucity of information on the morphometric variation of C. gariepinus in Nigeria. Hence, the present study, which aimed at evaluating the genetic diversity in C. gariepinus using multivariate analytical approach will however, provide preliminary information on the population structure of C. gariepinus in Nigeria.
MATERIALS AND METHODS
Sample collection: A total of 60 fish samples weighing approximately one kilogram were obtained from two wild and two cultured populations of C. gariepinus in Calabar for diversity assessment. Locations for the wild populations include Lemna River (LMN) and Okurikang Beach (OKU) while the cultured populations were Molecular Laboratory, 127 MCC Rd. Calabar (MCC) and University of Calabar Fish Farm (UFF). 15 fish samples of C. gariepinus were obtained from each of these sample locations. The fish samples were transported in 20 L capacity containers to the laboratory for analyses.
Morphometric measurements of fish: The following morphometric measurements were taken from each fish samples: Standard Length (SL), predorsal distance (PDD), preanal distance (PAD), preventral distance (PVD), prepectoral distance (PPD), Dorsal Fin Length (DFL), Anal Fin Length (AFL), Pectoral Fin Length (PFL), Pectoral Spine Length (PSL), distance between dorsal and caudal fin (DDCF), distance between occipital process and dorsal fin (DODF), Caudal Peduncle Depth (CPD), Body Depth at Anus (BDA), Head Length (HL), Head Width (HW), Snout Length (SNL), Interorbital Distance (ID), Eye Diameter (ED), Occipital Fontanelle Length (OFL), Width of occipital fontanelle (OFW), distance between snout and occipital process (DSO) using the method of Teugels (1982) (Fig. 1).
Proximate and mineral composition analyses: The fish samples were washed and weighed using electronic weighing scale (Model Kerro BL 20001). The samples were transferred into an air circulating oven and dried at temperature of 200̊C for 48 h. Dried samples were then cleaned, filleted and pulverized using electric blender to obtain a fine homogenous powder, which was then transferred into well labelled sterile bottles and stored in the refrigerator for proximate and mineral composition determination in triplicates. The proximate composition (moisture, ash, fat, fibre, protein and carbohydrate) were determined according to the official Methods of Analysis of the Association of Analytical Chemists (AOAC., 2005).
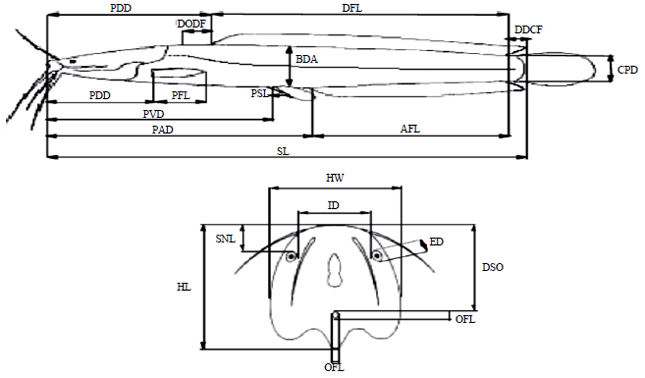 | |
| Fig. 1: | Location of measurements on Clarias gariepinus |
The protein content was determined using micro Kjeldhal method (% N X 6.25) while the carbohydrate content was estimated by the difference obtained after subtracting the total organic nitrogen, protein, lipid, ash and fibre from the total dry matter and expressing as percentage. For the mineral compositions, sodium and potassium were estimated using flame photometric technique while zinc, magnesium, calcium and iron contents were determined by atomic absorption spectrophotometric method (AOAC., 2005).
Statistics analysis: obtained were subjected to the analysis of variance (ANOVA), principal component analysis as well as cluster analyses using the computer software, Predictive Analytics Software (PASW) version 20.0.
RESULTS
Analysis of variance: There was significant differences (p<0.05) observed in the morphometric traits in the different C. gariepinus obtained from the two populations. The result revealed that C. gariepinus from the wild had higher morphometric measurements than the cultured, especially in their standard length, pre-dorsal and pre-anal distances, pre-pectoral distance, head length, head width, eye diameter, distance between occipital process and caudal fin as well as anal fin length (Table 1). The result also showed that there were significant differences (p<0.05) observed in the proximate and mineral compositions of C. gariepinus from the two populations. For instance, calcium and magnesium were higher in C. gariepinus from Itu River (wild) followed by C. gariepinus obtained from UNICAL fish farm. This was the same trend for other mineral elements, especially potassium and iron. For the proximate composition, C. gariepinus from UNICAL fish farm contain more protein followed by those selected from Lemna River. However, lipids were significantly higher in C. gariepinus from Itu River followed by those from MCC fish population. Other nutrients showed significantly varying levels but generally the cultured fish populations gave higher nutrient contents when compared with those from the wild (Fig. 2-4).
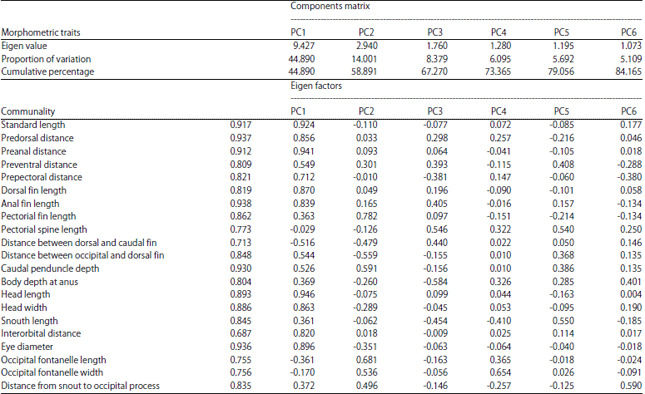
Principal component analyses for morphometric traits in Clarias gariepinus: Principal component analysis was carried out for 21 morphometric characters. Six principal components were extracted that had Eigen values greater than 1 (Eigen>1). PC1 and PC2 contributed 44.89 and 14.00%, respectively to the total variability of 84.17%. Morphometric traits with more than 90% communalities were standard length (92%), pre-dorsal distance (94%), pre-anal distance (91%), anal fin length (94%), caudal penduncle depth (93%) and eye diameter (94%) though other characters revealed high communalities greater than 50%.
| Table 1: | Pooled morphological traits obtained from 60 Clarias gariepinus sampled in the four populations |
 | |
*Means followed with the same case letter along horizontal array indicates no significance difference (p>0.05) | |
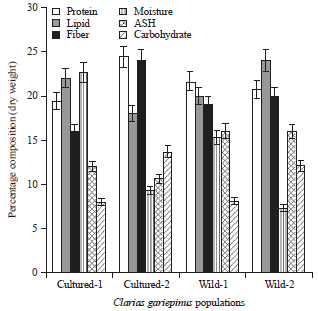 | |
| Fig. 2: | Percentage proximate composition of Clarias gariepinus in cultured and wild population |
From PC1, standard length of C. gariepinus, pre-dorsal and pre-anal distances, dorsal fin and anal fin lengths, head length, head width, inter-orbital distance and eye diameter contributed significantly to the total variability observed in C. gariepinus populations investigated. In PC2, pectoral fin length (0.782), caudal penduncle depth (0.591), occipital fontanelle length (0.681) and occipital fontanelle width (0.536) contributed highly to the total variability. Generally, however, in other PCs, there were some characters that contributed greater than 40% to the variability in the C. gariepinus populations (Table 2).
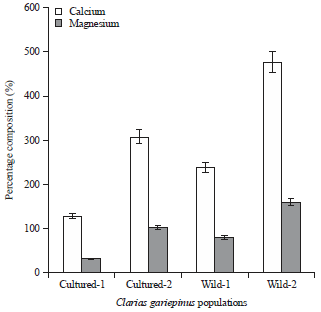 | |
| Fig. 3: | Percentage mineral composition of Clarias gariepinus in cultured and wild population |
Cluster analysis for morphometric, proximate and mineral composition in Clarias gariepinus: For the morphometric characters, the dendrogram revealed two major clusters, cluster-1 (1) and cluster-2 (19), where cluster-2 was further sub-clustered into cluster-2A (10) and cluster-2B (9), respectively. The result revealed that though cluster analysis pooled C. gariepinus species from wild and cultured populations together in the major cluster (Cluster-2), its sub-clusters-2A and 2B separated the clusters into wild and cultured (Fig. 5). Additionally, when the C. gariepinus populations were clustered based on proximate and mineral compositions, two major clusters were generated (Fig. 6).
| Table 2: | Pricipal components (PCs) for 21 morphometric traits in 60 samples of the four Clarias gariepinus populations |
 | |
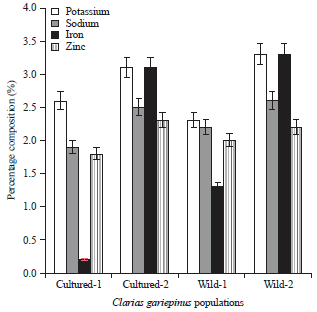 | |
| Fig. 4: | Percentage mineral composition of Clarias gariepinus in cultured and wild population |
Clarias gariepinus from different populations were pooled together in cluster-2, species in cluster-1 that was from the wild population notwithstanding. In the cluster-2 sub-clusters, there was no specific trend followed in the mini-clusters.
DISCUSSION
Genetic differences among species of organisms could be in terms of DNA sequences, biochemical (protein structure and isoenzymes properties) and physiological as well as in morphological traits. Danish et al. (2012) observed that genetic variability in fishes is very critical for aqua-culture and fishery management, stock identification and selection for breeding programmes. It also aids ecology restoration as well as estimation of genetic contribution in stock. According to Ikpeme et al. (2015), Calabar is seemingly the fastest growing farm and market for C. gariepinus with the implication that farmers need superior genotypes as to maximize productivity and profitability. This could be a leeway to combating the devastating challenge in the Sub-Saharan African countries, which is poverty amidst scourging malnutrition (Ikpeme et al., 2015). In Calabar, catfish species are usually called "Point and Kill", implying that consumers have certain criteria for selection, which are morphological through mere inspection. The result showed that C. gariepinus sampled from the wild significantly (p<0.05) performed better morphometrically than those from the cultured population. The result revealed that the standard length, head length, head width, eye diameter, anal fin length, pre-dorsal and pre-anal distances, etc. of C. gariepinus from the wild were significantly (p<0.05) greater than those from the cultured.
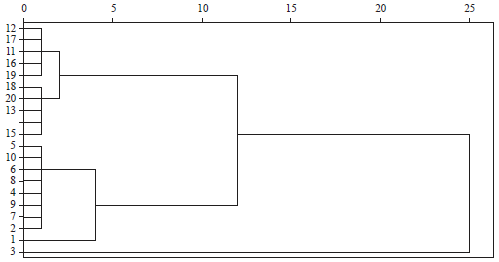 | |
| Fig. 5: | Ward method-based dendrogram of morphometric traits of Clarias gariepinus obtained from 4 populations [1-5 from OKU (wild), 6-10 from LMN (wild), 11-15 from MCC (cultured) and 16 - 20 from UFF (cultured)] |
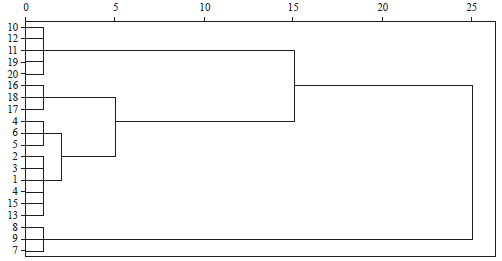 | |
| Fig. 6: | Wards method-based dendogram on proximate and mineral compositions of Clarias gariepinus obtained from 4 populations [1-5 from OKU (wild), 6-10 from LMN (wild), 11-15 from MCC (cultured), an d 16-20 from UFF (cultured)] |
This was similarly reported by Turan et al. (2005) and Saini et al. (2008) in separate experiments. According to Murta (2000), these morphological differences could either be attributed to genetic factors or associated with phenotypic plasticity in response to different environmental factors in each area. Specifically, the eye diameter of C. gariepinus from the wild populations is wider than those from the cultured. Matthews (1998) attributes this to turbidity differentials between the water bodies. It should be understood that the divergence in the eye diameter is an adaptive feature of the catfish in the wild to be able to see far objects. This morphometric divergence in eye diameter of C. gariepinus was also reported by Turan et al. (2005) in Turkey. Ikpeme et al. (2015) earlier findings using RAPD molecular markers in the same populations showed that there was wider genetic diversity among C. gariepinus in the wild than those in the cultured. The implication is that there exist relationship between genetic diversity as revealed by molecular markers based analysis and that using morphological technique. This feature might have been reduced in the catfish in the cultured population probably due to inbreeding depression, founder effect as well as genetic drift (Van Der Walt et al., 1993). This resultant low level of morphological diversity in the cultured population obviously has negative correlation with the potential for adaptation to changing environmental conditions (Leow, 2000).
It was also observed that there was significant differences (p<0.05) between the wild and cultured C. gariepinus population as regards to proximate and mineral compositions. For instance, the protein content in C. gariepinus from UNICAL farm (cultured) was significantly higher than that from Lemna River (wild). The differential in the protein content between the C. gariepinus from the wild and cultured populations could be attributed to the type of feed administered. Because the farmer wants to make quick returns on investment, the tendency is that the farmer feeds the fish with high/good quality feed, which may have manifested in the protein level. Omoniyi et al. (2013) reported similar finding. On the contrary, the mineral composition such as calcium and magnesium were higher in C. gariepinus from Itu River (wild) as compared with these mineral elements in C. gariepinus in the cultured. Omoniyi et al. (2013) observed that the mineral composition in the wild C. gariepinus were higher than in cultured C. gariepinus, especially in Mg, Mn and Cu while the reverse was the case with C. gariepinus in the cultured for Ca, P and Na. The obvious possibility is that C. gariepinus in the wild aquatic ecosystem may have absorbed or accumulated these mineral elements from the environment. It is therefore clear that the mineral element state of an aquatic body might determine the level of the said element (s) in the tissues of the aquatics. These variations, especially in proximate and mineral compositions in the wild and cultured C. gariepinus populations might be attributed to habitat change (Omoniyi et al., 2013; Onyia et al., 2013). It is suggested that the proximate and mineral composition in C. gariepinus might be largely environmental rather than genetic.
Principal component analysis showed that six principal components were extracted, which accounted for 84.10% variability with PC1 and PC2 contributing 44.89 and 14.00%, respectively. It was observed that the major morphological traits that contributed significantly to the variability were standard length, dorsal and anal fin lengths, head length, head width, eye diameter and pectoral fin length. Principal components revealed that morphometric differentiation between C. gariepinus from the wild and cultured populations were largely located in the head region. This suggests that these morphological traits might be useful in selection for breeding purposes. The above results as corroborated by Ikpeme et al. (2015) earlier report using RAPD markers’ technique could be the underlying genetic differences between C. gariepinus in the wild and cultured populations. This means that morphological or other types of genetic markers other than DNA markers could be very informative in estimating genetic diversity in species. Cluster analysis revealed population-based clustering, especially at the sub-cluster level. This showed that there exists morphological diversity between the C. gariepinus in the wild and those from the cultured, which was corroborated with the earlier report of Ikpeme et al. (2015) on the same population.
Giri et al. (2012) observed that the link between species conservation and efficient utilization of genetic resources is germplasm characterization. Understandably, breeders are more interested in the extent of genetic diversity presumably after germplasm characterization in any given population. This gives the extent to which he can appropriate selection for effective species development and improvement. There is more genetic diversity in C. gariepinus from the wild, which could be harnessed for breeding and genetic improvement purposes.
CONCLUSION
The present result suggests that C. gariepinus breeders and farmers should source C. gariepinus species from the wild in order to genetically enrich the gene pool. Additionally, head region-based morphometric traits might be more informative in distinguishing wild and cultured populations of C. gariepinus.
ACKNOWLEDGMENTS
The authors are grateful to the department of Genetics and Biotechnology, University of Calabar, Nigeria for providing counterpart funding for this research. We are sincerely grateful to Department of Animal Science, University of Calabar, Nigeria for her assistance.
REFERENCES
- Asuqwo, F.E. and J.P. Udoh, 2002. Patterns of total hydrocarbon, copper and iron in some fish from Cross River Estuary, Nigeria. West Afr. J. Applied Ecol., 3: 91-97.
CrossRefDirect Link - Babalola, T.O.O. and D.F. Apata, 2006. Effects of dietary protein and lipid levels on growth performance and body composition of African catfish Heterobranchus longifilis (Valenciennes, 1840) fingerlings. J. Anim. Vet. Adv., 5: 1073-1079.
Direct Link - Danish, M., I.J. Singh, P. Giri and C.P. Singh, 2012. Molecular characterization of two populations of catfish Clarias batrachus L. using random amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol., 11: 14217-14226.
Direct Link - Garg, R.K., P. Sairkar, N. Silawat, N. Vijay, N. Batav and N.N. Mehrotra, 2009. Genetic diversity between two populations of Heteropneustes fossilis (Bloch) using RAPD profile. Int. J. Zool. Res., 5: 171-177.
CrossRefDirect Link - Giri, P., G. Taj and H.S. Ginwal, 2012. Molecular characterization of six populations of Acorus calamus L. using randomly amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol., 11: 9522-9526.
Direct Link - Islam, M.N., A. Basak and M.S. Alam, 2012. Genetic diversity in wild and hatchery populations of stinging catfish (Heteropneustes fossilis Bloch) revealed by RAPD analysis. J. Bio-Sci., 19: 81-87.
Direct Link - Murta, A.G., 2000. Morphological variation of horse mackerel (Trachurus trachurus) in the Iberian and North African Atlantic: Implications for stock identification. ICES J. Mar. Sci., 57: 1240-1248.
CrossRefDirect Link - Nwafili, S.A., O.O. Soyinka and T.X. Gao, 2012. Levels and patterns of genetic diversity in wild Chrysichthys nigrodigitatusin Lagos lagoon complex. Afr. J. Biotechnol., 11: 15748-15754.
CrossRefDirect Link - Onyia, L.U., K.S. Michael, J.M. Manu and M. Sabo, 2013. Comparison of nutrient values of wild and cultured Heterobranchus bidorsalis and Clarias gariepinus. Nig. J. Fish. Aquacult., 1: 7-12.
Direct Link - Saini, A., A. Dua and V. Mohindra, 2008. Comparative morphometrics of two populations of giant river catfish (Mystus seenghala) from the Indus river system. Integrat. Zool., 3: 219-226.
CrossRefDirect Link - Teugels, G.G., 1982. Preliminary results of a morphological study of five African species of the subgenus Clarias (Clarias) (Pisces; Clariidae) J. Nat. Hist., 16: 439-464.
CrossRefDirect Link - Turan, C., D. Erguden, F. Turan and M. Gurlek, 2004. Genetic and morphologic structure of Liza abu (Heckel, 1843) populations from the rivers Orontes, Euphrates and Tigris. Turk. J. Vet. Anim. Sci., 28: 729-734.
Direct Link - Turan, C., M. Oral, B. Ozturk and E. Duzgunes, 2006. Morphometric and meristic variation between stocks of bluefish (Pomatomus saltatrix) in the Black, Marmara, Aegean and Northeastern Mediterranean seas. Fish. Res., 79: 139-147.
CrossRefDirect Link - Turan, C., S. Yalcin, F. Turan, E. Okur and I. Akyurt, 2005. Morphometric comparisons of African catfish, Clarias gariepinus, populations in Turkey. Folia Zool., 54: 165-172.
Direct Link - Van Der Walt, L.D., F.H Van Der Bank and G.J. Steyn, 1993. The suitability of using cryopreservation of spermatozoa for the conservation of genetic diversity in African catfish (Clarias gariepinus). Compa. Biochem. Psychol. Part A: Physiol., 106: 313-318.
CrossRefDirect Link - Omoniyi, P.M., F.E. Adedayo and A.J. Idowu, 2013. Variations in proximate and minerals composition of wild and cultivated Clarias gariepinus (burchell, 1822). Scient. J. Biol. Sci., 2: 219-226.
Direct Link - Ikpeme, E.V., O.U. Udensi, U.B. Ekaluo, M.E. Kooffreh, C.M. Okolo, P.B. Ekpo and N.C. Ogbonna, 2015. Unveiling the genetic diversity in Clarias gariepinus (Burchell, 1822) using Random Amplified Polymorphic DNA (RAPD) fingerprinting technique. Asian J. Anim. Sci., 9: 187-197.
CrossRefDirect Link - Olubunmi, A., A. Olateju, M. Latifat and A. Oluniyi, 2009. Production, growth and effect of varying stocking density of Clariobranchus fry. J. Fish. Int., 4: 73-78.
Direct Link - Eyo, V.O., A.P. Ekanem and A.I. Obiekezie, 2012. A comparative study of egg and sperm quality of Clarias gariepinus fed unical aqua feed and coppens commercial feed. Int. J. Biol. Pharm. Allied Sci., 1: 1598-1607.
Direct Link - Heok-Hee, 2003. Clarias insolitus: A new species of Clariid catfish (Teleostei: siluriformes) from Southern Borneo. Zootaxa, 284: 1-8.
Direct Link