
Zongyue Liu
Institute of Special Economic Animal and Plant Sciences, The Chinese Academy of Agricultural Sciences, Jilin Provincial Key Laboratory for Molecular Biology of Special Economic Animals, State Key Laboratory for Molecular Biology of Special Economical Animals, Changchun, 132112, China
Erjun Ren
Institute of Special Economic Animal and Plant Sciences, The Chinese Academy of Agricultural Sciences, Jilin Provincial Key Laboratory for Molecular Biology of Special Economic Animals, State Key Laboratory for Molecular Biology of Special Economical Animals, Changchun, 132112, China
Shiyong Wang
Institute of Special Economic Animal and Plant Sciences, The Chinese Academy of Agricultural Sciences, Jilin Provincial Key Laboratory for Molecular Biology of Special Economic Animals, State Key Laboratory for Molecular Biology of Special Economical Animals, Changchun, 132112, China
Xiumei Xing
Institute of Special Economic Animal and Plant Sciences, The Chinese Academy of Agricultural Sciences, Jilin Provincial Key Laboratory for Molecular Biology of Special Economic Animals, State Key Laboratory for Molecular Biology of Special Economical Animals, Changchun, 132112, China
Fuhe Yang
Institute of Special Economic Animal and Plant Sciences, The Chinese Academy of Agricultural Sciences, Jilin Provincial Key Laboratory for Molecular Biology of Special Economic Animals, State Key Laboratory for Molecular Biology of Special Economical Animals, Changchun, 132112, China
Asian Journal of Animal Sciences
Year: 2015 | Volume: 9 | Issue: 2 | Page No.: 73-79







ABSTRACT
The aim of this study was to use defined molecular markers to diagnose the self-biting behavior in blue fox. A single RAPD marker, S414, amplified a specific band of 768 bp in the stereotyped behavior group and this amplicon was designated SRS414. The sequences of SRS414 exhibited a 70% similarity to the Macaca mulatta MHC class I region. The RAPD marker was converted into a Sequence-Characterized Amplified Regions (SCAR) marker via cloning and sequencing of the RAPD amplification product and subsequent design of corresponding 18-mer oligonucleotide primers. The RAPD and SCAR markers were then validated in the two blue fox groups. The results of a χ2 test revealed a significant difference (p<0.05) between the detection rate of the two groups. This indicated that SCAR-S414 can be used as a positive marker to detect on farm stereotyped behavior.
PDF Abstract XML References Citation
Received: February 28, 2014;
Accepted: February 14, 2015;
Published: April 13, 2015
How to cite this article
Zongyue Liu, Erjun Ren, Shiyong Wang, Xiumei Xing and Fuhe Yang, 2015. Application of Sequence-Characterized Amplified Regions to the Detection of Self-biting Behavior in the Blue Fox. Asian Journal of Animal Sciences, 9: 73-79.
DOI: 10.3923/ajas.2015.73.79
URL: https://scialert.net/abstract/?doi=ajas.2015.73.79
DOI: 10.3923/ajas.2015.73.79
URL: https://scialert.net/abstract/?doi=ajas.2015.73.79
INTRODUCTION
Stereotypes are found in captive animals but are rare in the wild (Mason, 1991). The cage environment of farmed fur animals is sometimes considered to be a poor living condition but although more attention is generally paid to environmental impacts on farmed animals, the role of genetic origins cannot be overlooked. Stereotyped characteristics have been found to be heritable in bank voles (Clethrionomys glareolus) (Schoenecker and Heller, 2000) and african striped mice (Rhabdomys pumilio) (Schwaibold and Pillay, 2001) and the importance of genetic transmission in this regard has also been indicated in fur animals, such as mink (Hansen, 1993). Smith (1984) suggests that the occurrence of stereotypes in thoroughbred racehorses may be of genetic origin, as indicated by evidence from stereotyping stallions and producing stereotyped offspring. A similarly positive correlation between the occurrence of stereotypes in parents and their offspring has also been indicated in other species (Kiley, 1977; Hansen, 1993).
Genomic analysis generally proceeds along a specific pathway of investigations in order to identify genes involved in specific traits (Andersson, 2001) and determining the underlying mechanisms. Li et al. (2008) have reported that different fragments were amplified in the healthy and stereotyped mink by Random Amplified Polymorphic DNA (RAPD) primer S356 and that a fragment of 1000 bp was only amplified in the stereotyped behavior of mink groups. Liu et al. (2011) have also reported that SCAR markers (SRA8-250) are useful in detecting self-biting behavior in minks. Bulked Sergeant Analysis (BSA) is a rapid method for identifying markers linked to any specific gene or genomic region. This method has now been used widely in combination with the amplified fragment polymorphisms (AFLP)/RAPD method and has proven to be a very powerful and efficient technique for identifying markers linked to a specific gene (Wicks et al., 2001; Wang et al., 2001; Agrama et al., 2002; Zhang et al., 2006). Notably, some AFLP and RAPD markers linked to genes of interest have been converted into Sequence-Characterized Amplified Regions (SCAR) markers (Wang et al., 2001; Agrama et al., 2002; Zhang et al., 2006). Specific SCAR markers are developed with a pair of longer primers (usually the extended sequence of a RAPD primer) of approximately 20 bases. Compared with universal primers, unique primers for special regions prevent sit-competition and render the results less sensitive to reaction conditions and therefore more reproducible with enhanced specificity (Hernandez et al., 1999).
In present study, we employed BSA in combination with RAPD was employed to identify specific DNA sequences associated with stereotyped behavior and to then convert them into SCAR markers to distinguish between stereotyped and healthy animals.
MATERIALS AND METHODS
Sample collection: Experiments were carried out at the Fur Animals Experiment Station of the Institute of Special Economic Animals and Plants of the Chinese Academy of Agricultural Sciences in Northeast China. The healthy and stereotyped blue foxes were housed in standard roofed sheds with open sides in individual standard rearing cages (100x70x70 cm). Total genomic DNAs were extracted from blood samples using a phenol-chloroform procedure. Healthy and stereotyped blue fox DNA pools were prepared to examine the RAPD marker by BSA, each containing an equivalent amount of total DNA from each group.
RAPD-PCR analysis of genomic DNA: One hundred primers were sourced from the Shanghai Sangon Biological Engineering Technology Company for use in the RAPD-PCR analysis of the two sets of total DNA pools. PCR amplification were preformed in an Eppendorf AG thermal cycler (Gene Co., Ltd., Hamburg, Germany) in a 15 μL total volume containing 1U TaKaRa Taq DNA polymerase (TaKaRa Biotechnology Co., Ltd., Dalian, China), 2.5 μL of 10xPCR buffer (containing 500 mM KCl and 15 mM MgCl2), 0.2 mM of each dNTP, 6 μM of each primers and 2 μL of DNA template. The PCR cycling parameters were as follows: 30 cycles of 94°C for 30 sec, 60°C for 1 min and 72°C for 1.5 min with a initial hot start at 94°C for 5 min and a final extension at 72°C for 5 min. The PCR products were detected by 2% agarose gel electrophoresis. Additional healthy and stereotyped blue fox genomic DNA samples were used to test the RAPD marker, each group including 30 individuals.
Cloning and sequencing of the RAPD fragment: Once the RAPD fragment presented as polymorphic between the two sample pools, the band of amplified DNA corresponding to this segment was excised from the agarose gel, purified using an Agarose Gel DNA extraction Kit and cloned into the PMD-18T vector. The sequences obtained were analyzed using the BLAST program for a similarity search.
Design of primers and analysis of the SCAR marker: The SCAR primers (sense: 5' AACGGCCAGTGATTCGA 3' and anti-sense: 5’ CATTACCAGGCATCCATTCC 3’) were against the RAPD fragment sequence using Primer 5 software. The PCR conditions used for SCAR amplification were similar to those that were used in the RAPD analysis. Amplification products were all resolved electrophoretically on 2% agarose gels. Healthy and stereotype behavior individuals (15 of each) from another blue fox farm was tested for the SCAR marker. A χ2 test for independence in a 2x2 table was performed to determine whether a relationship exists between the RAPD and SCAR markers of the two groups. SAS8.0 software was used to analyze these data.
RESULT
RAPD-PCR analysis: High-molecular weight genomic DNA was isolated from fresh blood samples. To identify DNA markers linked to the stereotype behavior of the blue fox, 100 RAPD primers were screened using pooled total DNA from both the healthy and stereotyped groups of animals. The random primer S414 amplified an approximately 800 bp fragment (denoted SRS414-800) from the stereotype behavior group. This fragment was not detectable in the healthy group and was thus regarded as being specific to stereotype behavior (Fig. 1). The validity of the primer pair S414 in detecting a stereotyped fox was tested using 30 stereotyped and healthy foxes. A χ2 test indicated a significant difference (p<0.01) between the two groups (Table 1).
Sequencing analysis: The SRS414-800 fragment was excised from an agarose gel and then purified and sequenced. The sequence was found to be 781 bp in length and exhibited a 70% similarity to the Macaca mulatta MHC class I region. Query coverage and E values ranged from 33-38% and from 1x10-31 to 2x10-28, respectively.
Conversion of the RAPD marker into a SCAR marker: Based on the SRS414-800 sequence, a pair of primers designated SCAR-A8 was designed to convert the RAPD marker into a single locus SCAR marker. As expected, the primer pair amplified a 772 bp fragment from genomic DNA of the stereotype behavior group but not from the healthy animals (Fig. 2).
| Table 1: | Distribution of RAPD markers among healthy and stereotyped blue fox groups |
 | |
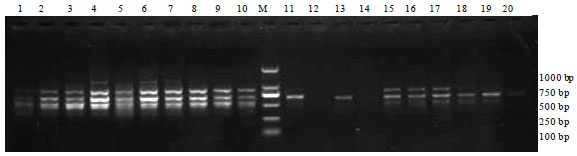 | |
| Fig. 1: | PCR results for healthy and stereotyped fox genetic DNA obtained using an RAPD marker. Lane M, DNA size markers; lanes 1~10, stereotyped blue foxes; lanes 11~20, healthy blue foxes |
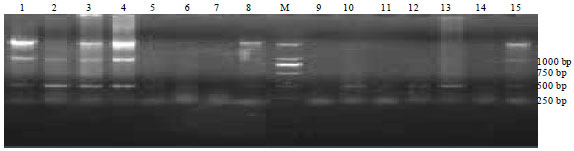 | |
| Fig. 2: | PCR results for healthy and stereotyped fox genetic DNA obtained using an SCAR marker. Lane M, DNA size markers; lanes 1~8, stereotyped blue foxes; lanes 9~15, healthy blue foxes |
| Table 2: | Distribution of SCAR markers among healthy and stereotyped blue fox groups |
 | |
The validity of the SCAR-S414 primer pair was tested using the DNA of blue fox from another farm. The χ2 test indicated significant difference (0.01<p<0.05) between the two groups (Table 2).
DISCUSSION
Stereotypic behavior or self-biting etiologies in farmed animals are usually multifarious, involving complex interactions among genetic, environmental, neurological, physiological and endocrinological and social factors (De Bellis et al., 1999a, b; Schore, 2002; Siegel, 1999; Van der Kolk, 1994). In the wild, fur animals are considered to be relatively social in their behaviors. The caged environment of farmed fur animals often considered to be an inferior living environment to natural habitats and to coincide with the emergence of stereotyped behaviors, can possibly be enriched by reconstructing the normal social systems for such animals (Ahola et al., 2007). In any event, careful analysis of animal emotions and behavior is an important approach to the improvement of animal welfare in applied ethological studies. The previous report of Moe et al. (2006) on anticipatory behavior may be useful for the future development of indicators of positive emotional states and, thus improved welfare, in farmed silver foxes. Correlations among behavioral problems and neurotransmitters, most notably plasma and platelet concentrations of serotonin (5-HT), dopamine (DA) and norepinephrine (NE), also have been found in some species, such as rats, rabbits, humans and dogs (Higley et al., 1992, 1996; Rogeness et al., 1992; Reisner et al., 1996; Horwitz and Mills, 2002). There is increasing evidence also that molecular methods or genes controlling aggressive behavior can be utilized to search for allelic differences which will assist in explaining phenotypic differences. Fewer data are available however on impulsive/compulsive behaviors, partly because the study of these characteristics is less unified across species.
Nevertheless, clear genetic effects do exist in relation to such traits. For instance, the strain type affects barbering by mice and tail biting by pigs (Breuer et al., 2003) and it is possible to breed high- and low-feather-pecking strains of laying hens (Kjaer et al., 2001). Lin and Bai (2008) have shown in this regard that polymorphisms in the 5-hydroxytryptamine 1A receptor gene and dopamine receptor D1 and D2 genes, had a direct association with stereotyped behaviors in the mink. Li et al. (2008) and Liu et al. (2011) have previously used the RAPD and SCAR methods to amply different fragments in healthy and stereotyped mink groups. Among these methods, PCR-based DNA markers are particularly desirable because methodologically they provide ease of use and fast and reproducible results. This strategy was employed in this study by combining bulked sergeant analysis and the RAPD method to rapidly identify molecular markers that are strictly linked to self-biting minks. The RAPD method on its own is easy to perform but is relatively poor in term of repeatability. Hence, the self-biting specific marker we identified was converted into a stable SCAR marker.
In our current analysis, the SCAR marker derived from a RAPD fragment showed a 70% similarity to Macaca mulatta MHC class I region, a tightly linked cluster of genes encoding proteins that are essential for the establishment and regulation of the immune response in most vertebrates. The canine MHC or Dog Leukocyte Antigen (DLA) complex has been one of the most systematically studied to date and eight functional genes and five pseudogenes have been identified to date within the class I and II regions in this species. Of particular note, the MHC class II molecules in the dog are constitutively expressed on the surface of the dendritic cells (DCs), macrophages, B cells, T cells, thymocytes and vascular endothelium (Doveren et al., 1985; Mueller et al., 2004). DCs and MHC class II molecules play crucial roles in regulating immune responses in the skin (Delves and Roitt, 2000). Enhanced MHC class II expression also has been documented in dogs with pyoderma (Day, 1996), ectoparasitism (Stemmer et al., 1996), immune-mediated dermatoses (Jackson et al., 2004) and neoplasia (Affolter and Moore, 2002). On the other hand, genes in the class II region in humans have been associated with susceptibility and resistance to a variety of diseases ranging from autoimmune diseases such as diabetes mellitus and rheumatoid arthritis to allergic diseases such as atopic dermatitis. The association of this genomic region with skin disorders may also implicate the MHC gene in the self-biting traits of caged blue foxes. This is supported by the data generated by the use of the SCAR marker developed in our current study.
All of current and marginally positive findings must be regarded as quite preliminary due to some noteworthy limitations. The sample size, especially in relation to the SCAR test, is quite small by the standard of a genetic study and with a sufficient sample size in this test, we believe that the P values would have been lower than those obtained by RAPD analysis. It should be noted also that the SCAR marker was detected in about 20% of healthy blue foxes. This may indicate that there is a threshold in the blue fox central nervous system for the onset of self-biting behavior even with the presence of the self-biting gene. Otherwise, the SCAR marker is dominant and cannot show the interaction between alleles. Since behavior is normally inherited in a polygenic, additive manner, there are actually many genes to be identified and in the future we will seek to estimate the contribution of each of these genes to the phenotypic variations observed in the stereotyped behaviors of farmed animals.
ACKNOWLEDGMENTS
The project was supported by the Special Fund for Public Welfare Technology Research in the Agricultural Industry (200903014).
REFERENCES
- Andersson, L., 2001. Genetic dissection of phenotypic diversity in farm animals. Nat. Rev. Genet., 2: 130-138.
CrossRefPubMedDirect Link - Affolter, V.K. and P.F. Moore, 2002. Localized and disseminated histiocytic sarcoma of dendritic cell origin in dogs. Vet. Pathol., 39: 74-83.
CrossRefPubMedDirect Link - Agrama, H.A., S.F. Houssin and M.A. Tarek, 2002. Cloning of AFLP markers linked to resistance to Peronosclerospora sorghi in maize. Mol. Genet. Genomics, 267: 814-819.
CrossRef - Breuer, K., M.E.M. Sutcliffe, J.T. Mercer, K.A. Rance, V.E. Beattie, I.A. Sneddon and S.A. Edwards, 2003. The effect of breed on the development of adverse social behaviours in pigs. Applied Anim. Behav. Sci., 84: 59-74.
CrossRef - Day, M.J., 1996. Expression of major histocompatibility complex class II molecules by dermal inflammatory cells, epidermal Langerhans cells and keratinocytes in canine dermatological disease. J. Comp. Pathol., 115: 317-326.
CrossRef - De Bellis, M.D., A.S. Baum, B. Birmaher, M.S. Keshavan and C.H. Eccard et al., 1999. Developmental traumatology part I: Biological stress systems. Biol. Psychiatry, 45: 1259-1270.
CrossRef - De Bellis, M.D., M.S. Keshavan, D.B. Clark, B.J. Casey and J.N. Giedd et al., 1999. Developmental traumatology part II: Brain development. Biol. Psychiatry, 45: 1271-1284.
CrossRef - Delves, P.J. and I.M. Roitt, 2000. Advances in immunology: The immune system-Second of two parts. N. Engl. J. Med., 343: 108-117.
Direct Link - Doveren, R.F., W.A. Buurman, B. Schutte, G. Groenewegen and C.J. van der Linden, 1985. Class II antigens on canine T lymphocytes. Tissue. Antigens, 25: 255-265.
PubMed - Hansen, C.P.B., 1993. Stereotypies in ranch mink: The effect of genes, litter size and neighbours. Behav. Processes, 29: 165-177.
CrossRefDirect Link - Higley, J.D., P.T. Mehlman, D.M. Taub, S.B. Higley, S.J. Suomi, M. Linnoila and J.H. Vickers, 1992. Cerebrospinal fluid monoamine and adrenal correlates of aggression in free-ranging rhesus monkeys. Arch. Gen. Psychiatry, 49: 436-441.
CrossRef - Higley, J.D., S.T. King Jr., M.F. Hasert, M. Champoux, S.J. Suomi and M. Linnoila, 1996. Stability of interindividual differences in serotonin function and its relationship to severe aggression and competent social behavior in rhesus macaque females. Neuropsychopharmacology, 14: 67-76.
CrossRefPubMedDirect Link - Hernandez, P., A. Martin and G. Dorado, 1999. Development of SCARs by direct sequencing of RAPD products: A practical tool for the introgression and marker-assisted selection of wheat. Mol. Breed., 5: 245-253.
CrossRefDirect Link - Jackson, H.A., T. Olivry, F. Berget, S.M. Dunston, C. Bonnefont and L. Chabanne, 2004. Immunopathology of vesicular cutaneous lupus erythematosus in the rough collie and Shetland sheepdog: A canine homologue of subacute cutaneous lupus erythematosus in humans. Vet. Dermatol., 15: 230-239.
CrossRefDirect Link - Li, Y., J. Yao, L. Ma, Z. Li and X. Bai, 2008. RAPD genetic analysis on etiological factor of mink self-biting disease. Sheng Wu Gong Cheng Xue Bao, 24: 563-568.
PubMed - Liu, Z.Y., F.Y. Ning, H.Y. Yang, L. Wei and X.J. Bai, 2011. Rapid detection of self-biting disease of mink by specific sequence-characterized amplified regions. J. Forest. Res., 22: 123-126.
CrossRef - Ahola, L., S. Hanninen and J. Mononen, 2007. A note on stereotyped behaviour in pair and group-housed farmed juvenile raccoon dogs. Applied Anim. Behav. Sci., 107: 174-180.
CrossRef - Reisner, I.R., J.J. Mann, M. Stanley, Y.Y. Huang and K.A. Houpt, 1996. Comparison of cerebrospinal fluid monoamine metabolite levels in dominant-aggressive and non-aggressive dogs. Brain Res., 714: 57-64.
CrossRefDirect Link - Rogeness, G.A., M.A. Javors and S.R. Pliszka, 1992. Neurochemistry and child and adolescent psychiatry. J. Am. Acad. Child Adolesc. Psychiatry, 31: 765-781.
CrossRef - Schore, A.N., 2002. Dysregulation of the right brain: A fundamental mechanism of traumatic attachment and the psychopatho genesis of posttraumatic stress disorder. Aust. N. Z. J. Psychiatry, 36: 9-30.
CrossRefPubMedDirect Link - Schoenecker, B. and K.E. Heller, 2000. Indication of a genetic basis of stereotypies in laboratory-bred bank voles (Clethrionomys glareolus). Applied Anim. Behav. Sci., 68: 339-347.
PubMed - Schwaibold, U. and N. Pillay, 2001. Stereotypic behaviour is genetically transmitted in the African striped mouse Rhabdomys pumilio. Applied Anim. Behav. Sci., 74: 273-280.
CrossRef - Smith, L.B., 1984. Electric field therapy for equine sterotypic behaviors. J. Biol. Phys., 12: 33-36.
CrossRef - Stemmer, B.L., L.G. Arlian, M.S. Morgan, C.M. Rapp and P.F. Moore, 1996. Characterization of antigen presenting cells and T-cells in progressing scabietic skin lesions. Vet Parasitol., 67: 247-258.
CrossRef - Wang, Z.L., L.X. Wang, J.R. Dai, B. Wang and X.Z. Li, 2001. Molecular identification and mapping of a maize gene (Rf3) in S-type CMS using AFLP, RFLP and SCAR techniques. J. Genet. Genomics, 28: 465-470.
PubMed - Zhang, Z.F., Y. Wang and Y.L. Zheng, 2006. AFLP and PCR-based markers linked to Rf3, a fertility restorer gene for S cytoplasmic male sterility in maize. Mol. Genet. Genomics, 276: 162-169.
CrossRef - Moe, R.O., M. Bakken, S. Kittilsen, H. Kingsley-Smith and B.M. Spruijt, 2006. A note on reward-related behaviour and emotional expressions in farmed silver foxes (Vulpes vulpes)-Basis for a novel tool to study animal welfare. Applied Anim. Behav. Sci., 101: 362-368.
CrossRef - Kjaer, J.B., P. Sorensen and G. Su, 2001. Divergent selection on feather pecking behaviour in laying hens (Gallus gallus domesticus). Applied Anim. Behav. Sci., 71: 229-239.
CrossRefDirect Link