
T. Prebavathy
Department of Veterinary Biochemistry,
J. Thanislass
Department of Veterinary Biochemistry,
Lydia Dhanammal
Department of Animal Genetics and Breeding,
R. Ganesan
Department of Animal Genetics and Breeding,
H.K. Mukhopadhyay
Department of Veterinary Microbiology, Rajiv Gandhi Institute of Veterinary Education and Research, Puducherry, 605 009, India
Asian Journal of Animal Sciences
Year: 2015 | Volume: 9 | Issue: 2 | Page No.: 45-56







ABSTRACT
The objective of this study was to understand the association between SNPs reported in the TLR2 gene of cattle and bovine mastitis. Allele Specific-PCR (AS-PCR) was developed for the detection of 6 SNPs (rs55617172, rs111026127, rs68268256, rs68268260, rs68343170 and rs68268268) which were reported to be responsible for change in amino acid present on the LRR-functional domain of TLR2 gene. Fifty well characterized mastitis cases in terms of California Mastitis Test, bacterial culture and PCR and fifty age-matched controls confirmed to be free from mastitis were selected from Puducherry region, India. The DNA was isolated from blood samples of the above animals. AS-PCR was performed with the custom designed primers and genotypes determined. The genotypes detected were further confirmed by sequencing and sequence analysis which had proved the efficiency of AS-PCR developed for the detection of SNPs in TLR2 gene. Statistical analysis of association between genotypes detected with the cases and control resulted in the identification of association (p = 0.0328) between TT genotype for SNP T→G at 385 mRNA position with the control and heterozygous genotype, CT for SNP C→T at 2010 mRNA position (p = 0.0006) with the mastitis. Odds Ratio (OR) analysis with 95% Confidence Intervals (CI) further confirmed significant (OR = 5.76, 95% CI = 2.07-15.97) association between the CT (C→T at 2010 mRNA position) heterozygous genotype and mastitis.
PDF Abstract XML References Citation
Received: December 08, 2014;
Accepted: March 07, 2015;
Published: April 13, 2015
How to cite this article
T. Prebavathy, J. Thanislass, Lydia Dhanammal, R. Ganesan and H.K. Mukhopadhyay, 2015. Association between SNPs in TLR2 Gene Segment Corresponding to LRR Functional Domain of TLR2 Receptor and Bovine Mastitis. Asian Journal of Animal Sciences, 9: 45-56.
DOI: 10.3923/ajas.2015.45.56
URL: https://scialert.net/abstract/?doi=ajas.2015.45.56
DOI: 10.3923/ajas.2015.45.56
URL: https://scialert.net/abstract/?doi=ajas.2015.45.56
INTRODUCTION
Mastitis is defined as an inflammatory reaction of the mammary gland. It is of great economic importance because it affects milk quality and production and can also spread to other cows in the herd (Lee et al., 2008). Mastitis can be either clinical or subclinical. In the absence of visible signs of infection, it is referred to as subclinical mastitis and which is the most prevalent form of mastitis (Akers, 2002). According to Shearer and Harris (1992) subclinical mastitis is important as it is 15-40 times more prevalent than the clinical form. In India too, the incidence of subclinical mastitis cases was were found to be greater (10-50%) than clinical mastitis (1-10%) (Joshi and Gokhale, 2006). Epidemiologic analyses revealed that the most prevalent agents associated with subclinical mastitis infections are Staphylococcus aureus (S. aureus), Streptococcus strains and Escherichia coli (E. coli) (Chaneton et al., 2008).
The incidence of subclinical mastitis has been found highest in purebred Holsteins and Jerseys and lowest in local cattle and buffaloes (Joshi and Gokhale, 2006) suggesting the role of genetic factors associated with the host to mastitis susceptibility. Identification of genetic factors responsible for mastitis will aid in selection of animals resistant to mastitis. Many genes like β-defensin 5, lactoferrin, lysozyme and lysostaphin (Cao et al., 2005) and Major Histocompatibility Complex (MHC) (Gao et al., 2006) have been researched for their association with mastitis. Describing genetic variation in these loci in relation to resistance against specific diseases may be useful in selection of animals for disease resistance/susceptibility.
In recent years, attention is being focused on the genes responsible for innate immunity and for their association with mastitis (Jault et al., 2004; Nonaka and Kimura, 2006). Among these, the gene responsible for TLR is important as it is responsible for receptors that recognize Pathogen Associated Molecular Patterns (PAMPs) on the surface of microorganisms. Several studies have shown that mutations in the TLR may reduce the ability of the protein to recognize PAMPs and therefore interfere with innate immune activation. Single Nucleotide Polymorphisms (SNPs) within TLR genes in humans have been found to be associated with susceptibility to specific infectious diseases (Pandey and Agrawal, 2006). A recent study revealed that two missense mutations in TLR4 Leucine Rich Repeat (LRR) domain were associated with Mycobacterium aviumpara tuberculosis (MAP) infection (Mucha et al., 2009) suggesting an association between TLR4 mutations and increased susceptibility to MAP infection in cattle.
Toll-like receptor 2 (TLR2) a member of TLR is strongly expressed during mastitis caused by S. aureus (Goldammer et al., 2004) and is uniquely capable of recognizing the peptidoglycan and lipoteichoic acid from S. aureus and other gram-positive bacteria (Bannerman et al., 2004). Bovine TLR2 can properly transduce signals from S. aureus and E. coli (Yang et al., 2008). Bovine TLR2 gene located at the proximal end of BTA 17 contains 2 exons and encodes 784 amino acids (White et al., 2003). Studies have identified polymorphic sites in the bovine TLR2 gene in Bos indicus and Bos taurus cattle breeds (Jann et al., 2008; Opsal et al., 2008; Mariotti et al., 2009; Seabury et al., 2010). Jann et al. (2008) observed that three SNPs of TLR2 gene were localized at positions shaped by positive selection in the ruminant dataset (Leu227Phe, His305Pro, His326Gln) and in domains involved in the recognition of ligands and which could be considered as candidate SNPs for immune related traits. Polymorphisms in the TLR2 gene encoding receptor are likely to contribute to the overall variation in the resistance or susceptibility to mastitis in dairy cattle. Thus, TLR2 appears to be an attractive candidate gene for S. aureus associated mastitis. Therefore the present study is planned to study the association between SNPs reported earlier present in the functional domain of TLR2 gene and bovine mastitis caused by S. aureus.
MATERIALS AND METHODS
Collection of samples: Crossbred cattle between 6-8 years of age were chosen from six private dairy farms in Puducherry region, India. Milk and blood samples were collected in sterile vials from 94 animals with a recurrent history of sub-clinical mastitis and from 72 apparently healthy animals with no history of mastitis. Udder was washed with potassium permanganate solution and teats wiped with 70% alcohol before the collection of the milk samples. One or two streams of milk were discarded before collecting samples from each quarter. Using the Californian Mastitis Test (CMT) reagent, the milk samples were screened for Somatic Cell Count (SCC) and thereby the stage of sub-clinical mastitis condition was ascertained. Blood samples were collected from jugular vein aseptically. Acid Citrate Dextrose (ACD) solution was used as an anticoagulant (2/10 mL blood). The samples were transported in an ice pack and kept at 4°C for further processing.
Isolation and identification of causative agents of mastitis: Milk samples were mixed well and two or three loop full of milk were streaked on Mueller Hinton Agar with 7% sodium chloride which inhibit the growth of other organisms and facilitate the growth of the genus Staphylococcus and incubated at 37°C for 24-48 h. The cultured organisms were then subjected to bacteriological analysis. A minimum of five colonies of the same type was recorded as causative agent and growth of more than one type of colonies was determined as mixed growth.
Identification of Staphylococcus sp.: Individual colonies were Gram stained. Only Gram positive cocci which were arranged in clusters were considered and the same individual colonies from the culture plates were streaked on Mueller Hinton Agar and incubated overnight at 37°C. A loop full of obtained culture was inoculated in 2 mL Luria Broth, incubated overnight at 37°C and the culture was used in the identification of S. aureus by PCR (Brakstad et al., 1992).
Nonsynonymous SNPs in TLR2 and primer designing: The region of six nonsynonymous SNPs corresponding to the functional domain of TLR2 i.e., LRR and TIR were obtained from the NCBI (www.ncbi.nlm.nih.gov). The corresponding messenger RNA sequence in FASTA format was downloaded and subjected to Batch Primer 3 to obtain allele specific primers (Table 1). Further, six sets of primers were designed for the detection of wild type genotype using Primer 3 (Table 2). These two different sets of primers were used for allele specific PCR for the detection of SNPs.
| Table 1: | AS-PCR-Allele specific primers for mutants |
 | |
| Table 2: | AS-PCR-Primers for the detection of wild type genotype |
 | |
| Table 3: | Sequencing primers |
 | |
Allele-specific polymerase chain reaction (AS-PCR): The DNA was isolated as per the method of Sambrook and Russel (2001). The quality of DNA isolated was determined using agarose gel electrophoresis and the ratio of absorbance at 260/280 nm. DNA with A260 and A280 ratio at = 1.8 were utilized in the study. The DNA samples were adjusted to a uniform concentration of 150 ng/5 μL and subjected to AS-PCR for chosen SNPs in TLR2 gene using custom designed primers.
Two reaction mixtures (20 μL each) were prepared for AS-PCR with two different forward primers (one wild forward primer and another mutant forward primer at 20 pmoles concentration) along with the regular ingredients of PCR at standard concentrations. Standard PCR conditions were followed with standardized annealing temperature of 58°C. The amplified products were tested on 2% agarose gel electrophoresis.
Sequencing and sequence analysis: The SNPs detected by AS-PCR were confirmed by sequencing and sequence analysis. Sets of primers shown in Table 3 were used to amplify the region of DNA containing the selected six different candidate SNPs present in the functional domain of TLR2 gene for sequencing. The amplified PCR products were custom sequenced. The specificity of the sequence obtained and the nucleotide variations with respect to the Bos taurus TLR2 sequence were determined using "BLAST" (Basic Local Alignment Search Tool) (www.ncbi.nlm.nih.gov).
Statistical analysis: Fisher’s exact test was used to study the case-control association using GraphPad software. A probability level (p) of less than 0.05 was considered as statistically significant. Odds Ratio (OR) with 95% confidence intervals was determined for each heterozygous allele and genotype frequency. OR with 95% confidence intervals were calculated using Medcalc Software (version 12.3.0) and the values were considered significant if the 95% confidence interval excludes the value of 1.
RESULTS
Collection and screening of samples: A total of 94 milk and blood samples were collected from cattle affected with a history of recurrent subclinical mastitis infection. The subclinical form was detected by CMT. Out of 94 milk samples, 82 were found positive for CMT which were also found positive for bacterial culture and of which 50 samples were found infected only with S. aureus, confirmed by S. aureus specific PCR. The samples were included under case group. Among milk samples collected from 72 apparently healthy animals with no history of mastitis infection, 50 samples were found negative for both CMT and bacterial culture and thus grouped as controls.
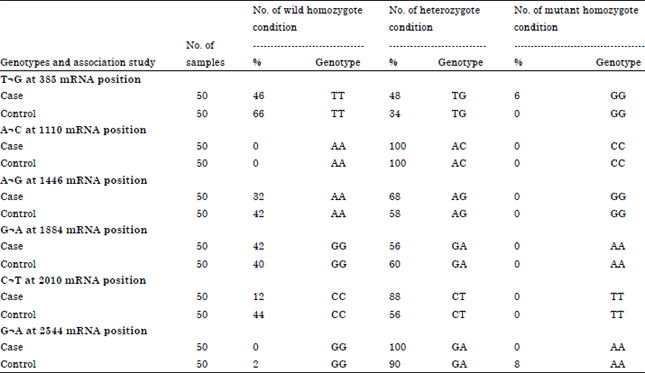
AS-PCR for the detection of genotype of SNPs: AS-PCR was conducted to genotype the SNPs selected. For which each sample was tested in two reactions, Reaction 1 with wild type forward primer and Reaction 2 with mutant type forward primer. The reverse primer was common for both. If the amplification was observed only in reaction 1, it was considered as a wild homozygote. If amplification was in reaction 2, it was considered a mutant homozygote. Amplification in both the reactions indicated a heterozygotic genotype. The results obtained are presented in Table 4. Agarose gel electrophoresis of PCR products obtained in AS-PCR is shown in Fig. 1.
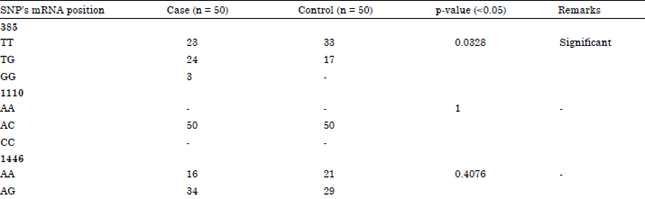
Case and control study: Association between SNPs and disease: A total of 100 samples were screened for 6 candidate SNPs in the case and the control groups (50 samples in each group). Based on the ‘p’ value obtained, the SNP T→G at mRNA position of 385 showed statistically significant association with the control. The SNP C→T at mRNA position of 2010 was having highly significant association with the cases (Table 5). The other SNPs did not show any significant association.
| Table 4: | Case-Control Study-Genotype frequency of SNPs studied |
 | |
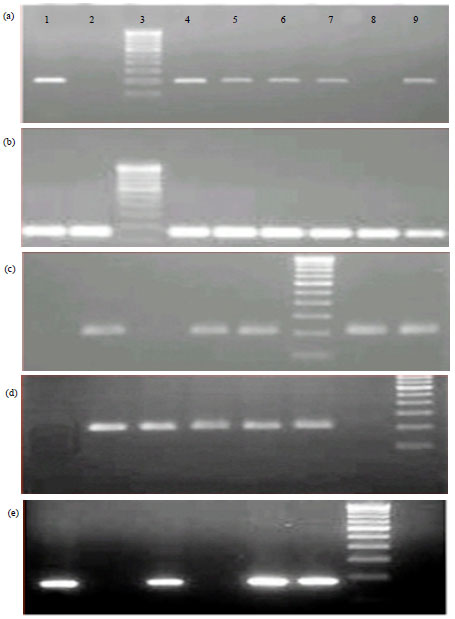 | |
| Fig. 1(a-e): | Agarose gel electrophoresis of AS-PCR product of SNP (T→G) at 385 mRNA position (a) Control, SNP (A→C) at 1110 mRNA position (b) Mastitis, SNP (A→G) at 1446 mRNA position (c) Control, SNP (G→A) at 2544 mRNA position (d) Mastitis and SNP (C→T) at 2010 mRNA position and (e) Mastitis. Samples are loaded in pair (Wild and Mutant). Amplification in one reaction mixture indicates homozygotic and in both indicates heterozygotic genotypes |
Confirmation of presence of SNPs by sequencing and sequence analysis: To identify the presence of selected SNPs and to confirm the specificity of the primer designed, PCR products of TLR2 gene which included the nucleotide region of 6 different selected SNPs were sent for sequencing. Six samples (3-Case and 3-Control) for each genotype were sequenced and the chromatograms showing the nucleotide variations are presented below. The genotypes of SNPs detected in the homozygous and heterozygous form were evident in the chromatograms (Fig. 2).
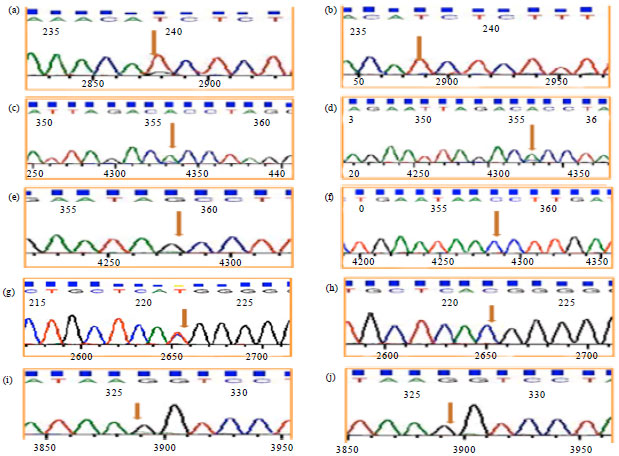 | |
| Fig. 2(a-j): | Chromatogram showing heterozygote condition (TG) (a, b) At 239 which corresponds to 385 position and homozygote for wild condition (TT) at 237 which corresponds to 385 position, (c, d) Heterozygote condition (AC) at 356 which corresponds to 1110 position, (e, f) Heterozygote condition (AG) at 358 which corresponds to 1446 position and wild homozygote condition (AA) at 357 which corresponds to 1446 position, (g, h) Heterozygote condition (CT) at 221 which corresponds to 2010 position and homozygote for wild condition (CC) at 222 which corresponds to 2010 position and (i, j) Heterozygote condition (GA) at 326 which corresponds to 2544 position and homozygote for wild condition (GG) at 326 which corresponds to 2544 position |
| Table 5: | Fischer’s exact analysis of genotype identified in cases and controls |
  | |
| Table 6: | Analysis of OR at 95% confidence interval for the detection of association between genotype and disease |
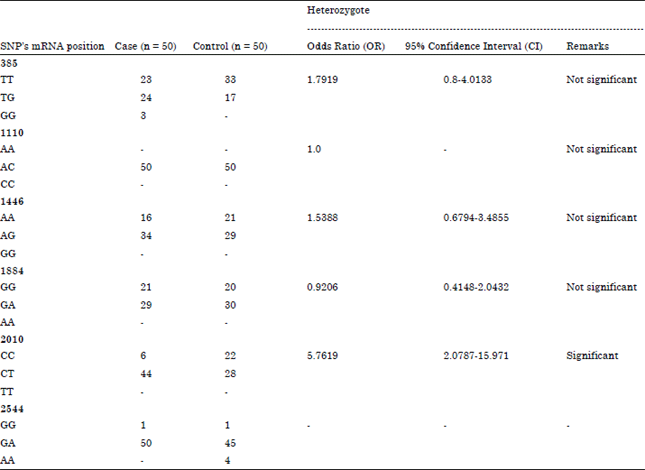 | |
In order to further validate the association between genotype and disease, the genotype frequencies were subjected for OR analysis at 95% Confidence Interval (CI). The heterozygotic genotype of SNP C→T at mRNA position of 2010 was found to have significant association with disease (Table 6).
DISCUSSION
Several studies have shown that mutations in the TLR may reduce the ability of the protein to recognize PAMPs and therefore interfere with innate immune activation. Toll-like receptors are Type I trans-membrane proteins. The extra-cellular domain (Ectodomain) of TLR contains Leucine Rich Repeats (LRR) and all TLRs have a single trans-membrane domain and a cytoplasmic domain that is homologous to the cytoplasmic tail of the IL-1 receptor (Medzhitov et al., 1997). The LRR domain plays a very important role in recognizing Pathogen Associated Molecular Patterns (PAMP’s) of the invading microbes. The LRR consensus motif forms the inner core of horseshoe structured ectodomain, while extra LRR regions forms convex surface. Mutations in convex surface may affect PAMP binding onto the TLR horseshoe (Bhide et al., 2009) and makes the animals susceptible for disease conditions.
Polymorphisms, in particular, nonsynonymous SNPs in the TLR2 gene are likely to contribute to the overall variation in the resistance or susceptibility to mastitis in dairy cattle. So far, 29 and 83 polymorphisms with 15 and 10 nonsynonymous SNPs have been reported that are located within the predicted functional domains of TLR2 and TLR6, respectively. Prediction and comparison of protein domain architectures for TLR2 and TLR6 revealed 6 regions of LRR patterning that were conserved among multiple species. The average polymorphism in bovine TLR2 gene was 1 SNP per 75 bp sequenced (Seabury and Womack, 2008).
Out of 15 nonsynonymous SNPs reported in bovine TLR2, 12 were present on the LRR and 2 were present on the Toll-Interleukin-Resistance (TIR) homologous domain and 1 present on the transmembrane domain in normal animals. Among the 12 SNPs, six nonsynonymous SNPs namely rs55617172, rs111026127, rs68268256, rs68268260, rs68343170 and rs68268268 resulting in the change in amino acids Asp63Glu, His305Pro, Asn417Ser, Arg563His, Thr605Met and Arg783Lys respectively, They were selected randomly for the association study. AS-PCR for easy genotyping of these 6 SNPs was developed.
SNPs were detected using AS-PCR primers which were designed in such a way that the 3’ nucleotide of a primer corresponded to the site of the SNP (Ugozzoli and Wallace, 1991). Thus, the allele-specific primer matches perfectly with one allele (the specific allele) and has a 3’ mismatch with the nonspecific allele. Mismatched 3’ termini were extended by DNA polymerases with much lower efficiency than correctly matched termini (Petruska et al., 1988). Thus AS-PCR could detect the given SNP. The advantage of AS-PCR is that it is methodologically simple and does not require special machines except a thermal cycler. Once the optimal conditions (i.e., optimal annealing temperature) were standardized, the genotyping of the specific SNP can be easily done by AS-PCR (Muneta et al., 2012). Allele-specific PCR procedures have the additional benefit that restriction enzyme digestion is not necessary after PCR amplification for SNPs detection.
Six sets of primers (each set with 2 forward primers and 1 reverse primer) were designed for AS-PCR for the detection of 6 SNPs. Two reactions were carried out for each sample for one SNP, one with wild type forward primer and reverse primer and the other with mutant type forward primer and reverse primer. The DNA isolated from totally 100 samples (50 mastitis cases and 50 age matched controls) were subjected to AS-PCR with the primers as mentioned above. The primers had efficiently detected the genotype of SNPs as homozygotic wild type, homozygotic mutant type and heterozygotic type. The genotype detected by the AS-PCR developed was, further confirmed by sequencing and sequence analysis. PCR products of representative samples (four samples for each genotype) were custom sequenced. The specificity of the sequences was confirmed by BLAST analysis. Further, the chromatogram of each sequence was checked for the presence of single peak (homozygous) or overlapping peaks (heterozygous) at the position of SNPs reported. The chromatograms obtained clearly supported the type of genotype of SNPs detected by the developed AS-PCR, thereby supporting the efficiency of AS-PCR for the detection of genotype of SNPs.
In order to study the association between genotype detected and mastitis, genotype frequency of each SNP’s was subjected for Fisher’s exact test with a probability level (p) of less than 0.05, considered statistically significant. Odds Ratio (OR) with 95% confidence intervals were determined for each heterozygous allele for its association with disease.
Among the different genotypes, TT homozygous genotype identified for the SNP T→G at 385 mRNA position (rs55617172) was significantly (p = 0.0328) associated with the control animals whereas CT heterozygous genotype for the SNP C→T at 2010 mRNA position (rs68343170 was found to be significantly (p = 0.0006) associated with mastitis cases. Further analysis of association between heterozygous genotype frequencies and mastitis in cases and control using OR with 95% confidence intervals, revealed strong association (OR = 5.76, 95% CI = 2.07-15.97) between CT heterozygous genotype at 2010 mRNA position with mastitis. The SNP C→T is responsible for change in amino acid from Threonine→Methionine i.e., polar to non-polar amino acid which could have structural consequences thereby which may predispose the animal to mastitis. Therefore the CT genotype for the SNP C→T at 2010 mRNA position can be considered as a Type I DNA marker for susceptibility to bovine mastitis. SNP T→G at 385 mRNA position can be used as a DNA marker for mastitis resistance. The effects of TLR2 polymorphisms on Somatic Cell Score (SCS) were analyzed and significant association was found between T385G and SCS. The mean of genotype GG was significantly lower than those of genotype TT and TG. No significant associations were found with SCS for G398A and G1884A (Zhang et al., 2009). Huang et al. (2011) conducted a case-control study and association analysis of SNPs identified in the 5’ upstream of TLR2 gene and revealed that the cow with the genotype TT had significantly lower somatic cell score than that with genotype TG or GG in Chinese Holstein cattle.
REFERENCES
- Bannerman, D.D., M.J. Paape, J.W. Lee, X. Zhao, J.C. Hope and P. Rainard, 2004. Escherichia coli and Staphylococcus aureus elicit differential innate immune responses following intramammary infection. Clin. Diagn. Lab. Immunol., 11: 463-472.
PubMed - Bhide, M.R., R. Mucha, I. Mikula, L. Kisova, R. Skrabana, M. Novak and I. Mikula, 2009. Novel mutations in TLR genes cause hyporesponsiveness to Mycobacterium avium subsp. paratuberculosis infection. BMC Genet., Vol. 10.
CrossRefDirect Link - Brakstad, O.G., K. Aasbakk and J.A. Maeland, 1992. Detection of Staphylococcus aureus by polymerase chain reaction amplification of the nuc gene. J. Clin. Microbiol., 30: 1654-1660.
PubMedDirect Link - Cao, S.Z., H.B. Li, A.H. Wang, X.X. Zhao and L.X. Du, 2005. Construction of mastitis resistant DNA library in dairy cows using suppression subtractive hybridization. Acta Veterinaria Zootechnica Sinica, 36: 526-530.
Direct Link - Chaneton, L., L. Tirante, J. Maito, J. Chaves and L.E. Bussmann, 2008. Relationship between milk lactoferrin and etiological agent in the mastitic bovine mammary gland. J. Dairy Sci., 91: 1865-1873.
CrossRef - Gao, S.X., S.Z. Xu, J. Li, X. Gao, H.Y. Ren, J.B. Chen and Y. Ma, 2006. Analysis of polymorphisms of BoLA-DQA and DRB3 * exon2 and the relationship between the gene and mastitis in dairy cattle. Acta Veterinaria Zootechnica Sinica, 37: 317-320.
Direct Link - Goldammer, T., H. Zerbe, A. Molenaar, H.J. Schuberth, R.M. Brunner, S.R. Kata and H.M. Seyfert, 2004. Mastitis increases mammary mRNA abundance of β-defensin 5, Toll-Like-Receptor 2 (TLR2) and TLR4 but not TLR9 in cattle. Clin. Diagn. Lab. Immunol., 11: 174-185.
CrossRefDirect Link - Jann, O.C., D. Werling, J.S. Chang, D. Haig and E.J. Glass, 2008. Molecular evolution of bovine Toll-like receptor 2 suggests substitutions of functional relevance. BMC Evol. Biol., Vol. 8.
CrossRefDirect Link - Jault, C., L. Pichon and J. Chluba, 2004. Toll-like receptor gene family and TIR-domain adapters in Danio rerio. Mol. Immunol., 40: 759-771.
CrossRefDirect Link - Huang, J., L. Liu, H. Wang, C. Zhang and Z. Ju et al., 2011. Variants and gene expression of the TLR2 gene and susceptibility to mastitis in cattle. Asian J. Anim. Vet. Adv., 6: 51-61.
CrossRefDirect Link - Lee, K.H., J.W. Lee, S.W. Wang, L.Y. Liu and M.F. Lee et al., 2008. Development of a novel biochip for rapid multiplex detection of seven mastitis-causing pathogens in bovine milk samples. J. Vet. Diagn. Invest., 20: 463-471.
PubMedDirect Link - Mariotti, M., J.L. Williams, S. Dunner, A Valentini and L. Pariset, 2009. Polymorphisms within the Toll-Like Receptor (TLR)-2, -4 and -6 genes in cattle. Diversity, 1: 7-18.
Direct Link - Medzhitov, R., P. Preston-Hurlburt and C.A. Janeway Jr., 1997. A human homologue of the drosophila toll protein signals activation of adaptive immunity. Nature, 388: 394-397.
PubMed - Mucha, R., M.R. Bhide, E.B. Chakurkar, M. Novak and I. Mikula Sr., 2009. Toll-like receptors TLR1, TLR2 and TLR4 gene mutations and natural resistance to Mycobacterium avium subsp. paratuberculosis infection in cattle. Vet. Immunol. Immunopathol., 128: 381-388.
CrossRef - Nonaka, M. and A. Kimura, 2006. Genomic view of the evolution of the complement system. Immunogenetics, 58: 701-713.
CrossRefDirect Link - Opsal, M.A., S. Lien, S. Brenna-Hansen, H.G. Olsen and D.I. Vage, 2008. Association analysis of the constructed linkage maps covering TLR2 and TLR4 with clinical mastitis in Norwegian Red cattle. J. Anim. Breed. Genet., 125: 110-118.
PubMed - Pandey, S. and D.K. Agrawal, 2006. Immunobiology of Toll-like receptors: Emerging trends. Immun. Cell. Biol., 84: 333-341.
CrossRefDirect Link - Petruska, J., M.F. Goodman, M.S. Boosalis, L.C. Sowers, C. Cheong and I. Tinoco, 1988. Comparison between DNA melting thermodynamics and DNA polymerase fidelity. Proc. Natl. Acad. Sci., 85: 6252-6256.
Direct Link - Muneta, Y., Y. Minagawa, M. Kusumoto, H. Shindai, H. Uenishi and I. Splichal, 2012. Development of allele-specific primer PCR for a swine TLR2 SNP and comparison of the frequency among several pig breeds of Japan and the Czech republic. J. Vet. Med. Sci., 74: 553-559.
Direct Link - Sambrook, J. and D.W. Russell, 2001. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory Press, New York, USA., ISBN-13: 9780879695774, Pages: 2344.
Direct Link - Seabury, C.M. and J.E. Womack, 2008. Analysis of sequence variability and protein domain architectures for bovine peptidoglycan recognition protein 1 and Toll-like receptors 2 and 6. Genomics, 92: 235-245.
PubMed - Seabury, C.M., P.M. Seabury, J.E. Decker, D.R. Schnabel, J.F. Taylor and J.E. Womack, 2010. Diversity and evolution of 11 innate immune genes in Bos taurus taurus and Bos taurus indicus cattle. Proc. Natl. Acad. Sci., 107: 151-156.
Direct Link - Ugozzoli, L. and R.B. Wallace, 1991. Allele-specific polymerase chain reaction. Methods, 2: 42-48.
CrossRefDirect Link - White, S.N., S.R. Kata and J.E. Womack, 2003. Comparative fine maps of bovine toll-like receptor 4 and toll-like receptor 2 regions. Mamm. Genome, 14: 149-155.
Direct Link - Yang, W., H. Zerbe, W. Petzl, R.M. Brunner and J. Gunther et al., 2008. Bovine TLR2 and TLR4 properly transduce signals from Staphylococcus aureus and E. coli, but S. aureus fails to both activate NF-κB in mammary epithelial cells and to quickly induce TNFα and interleukin-8 (CXCL8) expression in the udder. Mol. Immunol., 45: 1385-1397.
PubMed - Zhang, L.P., Q.F. Gan, T.H. Ma, H.D. Li and X.P. Wang et al., 2009. Toll-like receptor 2 gene polymorphism and its relationship with SCS in dairy cattle. Anim. Biotechnol., 20: 87-95.
CrossRefPubMedDirect Link