
Hossam H. Azzaz
Department of Dairy Science, National Research Centre, Dokki, Giza, Egypt
Hussein A. Murad
Department of Dairy Science, National Research Centre, Dokki, Giza, Egypt
Tarek A. Morsy
Department of Dairy Science, National Research Centre, Dokki, Giza, Egypt
Asian Journal of Animal Sciences
Year: 2015 | Volume: 9 | Issue: 6 | Page No.: 254-265







ABSTRACT
Ionophores are feed additives that alter rumen microbial populations through ion transfer across cell membranes. Although ionophores have been used widely in the ruminant’s nutrition for improved feed efficiency and control of coccidiosis. Ionophores have been studied regarding several potential effects on ruminant animals, including influences on health, meat production, milk production and reproduction. This review will provide practitioners with relevant references in the published literature regarding ionophore use in ruminants' nutrition. It should also give some guidance as to what effects might be anticipated with the use of ionophores in ruminant animals.
PDF Abstract XML References Citation
Received: August 01, 2015;
Accepted: September 07, 2015;
Published: September 18, 2015
How to cite this article
Hossam H. Azzaz, Hussein A. Murad and Tarek A. Morsy, 2015. Utility of Ionophores for Ruminant Animals: A Review. Asian Journal of Animal Sciences, 9: 254-265.
DOI: 10.3923/ajas.2015.254.265
URL: https://scialert.net/abstract/?doi=ajas.2015.254.265
DOI: 10.3923/ajas.2015.254.265
URL: https://scialert.net/abstract/?doi=ajas.2015.254.265
INTRODUCTION
A long cherished dream of ruminant nutritionists has been to manipulate and improve the efficiency of ruminal fermentation. In particular, this has meant to increase ruminal propionic acid yield, depress methanogenesis and depress rapid ruminal proteolysis and deamination of dietary proteins. Research is continued to find a sole feed supplement that can improve the rate, efficiency and/or quality of gain, production, reproduction, to prevent certain diseases or preserve feeds (Azzaz et al., 2012). Several strategies have been used to achieve some or all of the above objectives by rumen ecology manipulation using some dietary additives and/or supplements including buffers, anabolic hormones, feed enzymes, synthetic amino acids, essential oil and microbial feed supplements (Azzaz et al., 2013, 2015a, b; Murad and Azzaz, 2013; Morsy et al., 2012; Kholif et al., 2012). In the mid-1970s a number of active compounds have been discovered that when fed, can improve ruminant production efficiency. One such class of these compounds is carboxylic polyether ionophore antibiotics which form lipid-soluble complexes with certain cations and facilitate their transport across biological membranes (Pressman, 1976). These ionophores are produced by various strains of Streptomyces and include monensin (RumensinTM), lasalocid (BovatecTM), laidlomycin propionate (CattlystTM), salinomycin and narasin. Monensin is polyether antibiotic which is orally fed as a sodium salt and consider the most widely used preparation of the different kinds of ionophores (Yang et al., 2007). Bergen and Bates (1984) reported that benefits derived by ruminant from the biological actions of ionophores were classified into three areas of effects as follows:
| • | Increased efficiency of energy metabolism of rumen bacteria and (and/or) the animal |
| • | Improved nitrogen metabolism of rumen bacteria and (or) the animal |
| • | Retardation of digestive disorders resulting from abnormal rumen fermentation |
Each action provides nutritional and metabolic advantages to the ionophore-supplemented animal over a non-supplemented animal. The animal transforms these into increased production or improved efficiency. Economic benefits derived from feeding ionophores include improved feed efficiency, increased weight gain and a reduction in morbidity and mortality rate (Van Nevel and Demeyer, 1988). In addition, McGuffey et al. (2001) reported that cows fed monensin had higher average milk yield (1.3 kg day–1), higher milk protein yield (26 g day–1) and lower milk fat and protein percent while, milk fat yield and dry matter intake were not different for control and monensin fed cows. Also, Symanowski et al. (1999) detected improvement in milk production efficiency by 3.6%, for monensin premix treated cows while, McGuffey and Giner-Chavez (1998) reported that monensin premix supplemented cows showed enhancement in milk production efficiency by 7.0%. Ionophores also help reduce the amount of excreta and gaseous emissions from animals. Thus, a positive effect on the environment occurs when ionophores are fed. This review will describe the general effect of ionophores on ruminal ecology and on cellular responses of ruminal anaerobes.
General properties of ionophores: Ionophores are highly lipophilic substances capable of interacting stoichiometrically with metal ions, thereby serving as a carrier by which these ions can be transported across a bimolecular lipid membrane (Ovchinnikov, 1979). Ionophores are toxic to many bacteria, protozoa, fungi and higher organisms and thus fit the classical definition of antibiotics (Pressman, 1976). The exterior of the molecule is hydrophobic, while the interior is hydrophilic and able to bind cations. Some ionophores bind only a single cation (uniporters), but others are able to bind more than one cation (antiporters). Because cell membranes are composed of lipid bilayers, high activation energy is needed to translocate ions. Ionophores are able to shield and delocalize the charge of ions and facilitate their movement across membranes.
Painter et al. (1982) explain the action of ionophore through the cation-proton exchange, which is mediated by a carboxylic acid ionophore (Fig. 1). The transport cycle begins with the anionic form of the ionophore confined to the membrane interface where it is stabilized by the polar environment characteristic to the surface of a membrane. As an anion, the ionophore is capable of ion pairing with a metal cation either at the terminal carboxylic acid moiety or at other internal sites. The binding of a cation initiates the formation of a lipophilic, cyclic cation-ionophore complex that can diffuse through the interior of the biomolecular membrane structure. Ultimately, the complex reaches the opposite face of the membrane where it is again subjected to a polar environment. The electrostatic forces that had stabilized the complex are no longer greater than the unfavorable AG (Gibbs free energy change) of cyclization and the ionophore releases its enclosed cation and reverts to the low energy acyclic conformation. There it awaits the next phase of the transport cycle. Both monensin and lasalocid fall into this classification of ionophores (Pressman, 1976). A salient feature of this model is that the ionophore must be in the anionic form before it is capable of binding a metal cation (M*). Furthermore, diffusion across the membrane cannot occur unless the ionophore exists in the protonated form (ionophore I-g) or as a zwitterion (M* and anionic form of ionophore). Ionophores as a carriers are selective for specific ions and this selectivity is an index of ion-binding preference, but the final distribution of ions and direction of movement is ultimately determined by the initial concentrations of ions across the cell membrane and proton motive force (Painter et al., 1982).
Mechanism of ionophore action in ruminal bacteria: Ruminal bacteria maintain high intracellular potassium and low intracellular sodium concentrations (Chow and Russell, 1992) and conversely, the ruminal environment contains high sodium and low potassium concentrations.
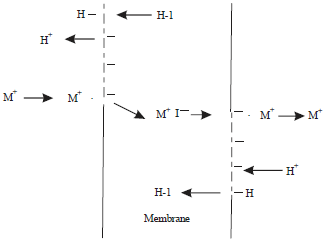 | |
| Fig. 1: | Carboxylic ionophore mediated cation transfer across a bimolecular lipid membrane, M+: Metal cation, I: Ionophore, H+: Proton, H-I: Protonated ionophore, M+I‾: Zwitterion of metal cation and anionic form of ionophore |
Thus, ruminal bacteria rely heavily upon ion gradients (both K+ and Na+ gradients) to take up nutrients and to establish a Proton Motive Force (PMF) (Rosen, 1986; Dawson and Boling, 1987; Van Kessel and Russell, 1992). Ruminal pH is somewhat acidic due to VFA concentrations; however the intracellular pH of many ruminal bacteria is near neutral, thus creating an inwardly directed proton gradient (Russell and Strobel, 1989).
Monensin is a metal/proton antiporter that can exchange H+ for either Na+ or K+ (Pressman, 1976; Russell and Strobel, 1989). Once inserted in the membrane, monensin exchanges intracellular potassium ions for extracellular protons or extracellular sodium for intracellular protons (Russell, 1987) (Fig. 2). Because the potassium gradient is greater than the sodium gradient, protons accumulate inside the bacterium (Chow et al., 1994). The bacterium reacts to this cytoplasmic acidification by activating a reversible ATPase to pump these protons out of the cell (Booth, 1985).
Additionally, other ATP-utilizing primary pumps for Na+ removal and K+ uptake are activated to reestablish ion gradients; resulting in the uncoupling of ATP hydrolysis for growth, thereby decreasing intracellular ATP pools, leading to cellular death (Russell, 1987; Russell and Strobel, 1989).
Effects of ionophores on energetics of ruminal fermentation: Biological actions of ionophores in ruminant are limited to the gastrointestinal tract. Anaerobic fermentation in the rumen derives energy from substrate oxidation by the transfer of electrons (and hydrogen) to acceptors other than oxygen. The reduced compounds formed are mainly VFA and methane. Fermentation balance requires that an increase in propionate production must be accompanied by a decrease in methane production (Chalupa, 1977; Demeyer and van Nevel, 1975). Up to 12% of the gross energy of feeds can be lost as eructated methane. Diverting hydrogen to other end products captures more digestible energy from fermented OM, resulting in more efficient use of feed energy. It also lessens the contribution of cattle to atmospheric methane accumulation. Raun (1974) discovered that monensin enhances rumen propionate production and was able to implement this biological action into the first commercial application of ionophores for the cattle industry in 1975.
Numerous studies have demonstrated that the addition of ionophores to mixed cultures of rumen microbes’ in vitro increases the production of propionic acid and reduces production of methane.
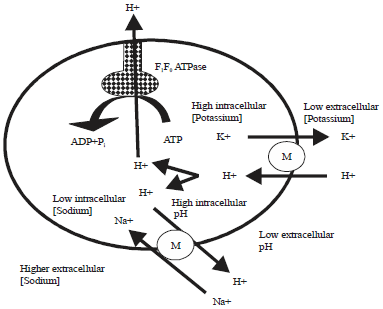 | |
| Fig. 2: | Gradient dissipating mechanism of monensin |
Similar effects were demonstrated in vivo by the studies of Richardson et al. (1976). Increases in the molar proportion of propionate in many studies were smaller than the changes in production rates, indicating that measurements of VFA concentration in rumen fluid can underestimate the effects of monensin on VFA production.
Improved energetics of rumen fermentation caused by monensin is illustrated by the work of Rogers and Davis (1982). Steers were fed a basal diet of 50% corn silage and 50% concentrate, either with or without monensin (33 mg kg–1 of DM). Monensin increased the daily ruminal production of acetic, propionic and total acids per kilogram of DM consumed by 29, 64 and 35%, respectively. Total VFA energy produced in the rumen per kilogram of DM consumed was increased from 0.852 Mcal kg–1 of DM for control steers to 1.137 Mcal kg–1 of DM for steers fed monensin, a 33% increase in ruminal digestible energy.
Ionophores inhibit methanogenesis by lowering the availability of hydrogen and formate, the primary substrates for methanogens. Bacteria that produce these substrates are sensitive to ionophores, whereas methanogens are more resistant (Chen and Wolin, 1979). Additional evidence for this mechanism is that, in the presence of monensin, methane production by mixed cultures of rumen microbes can be increased by adding hydrogen gas (Van Nevel and Demeyer, 1977). Bacteria that reduce succinate to propionate are resistant to ionophores, so propionate production increases. Defaunation by ionophores may also be partly responsible for the effect because protozoa produce hydrogen and are colonized by methanogens (Russell and Strobel, 1989). Similarly, rumen fungi, which also produce hydrogen, are sensitive to monensin in vitro (Marounek and Hodrova, 1989). Effects of ionophores have also been noted in ruminant hindgut fermentation (Marounek et al., 1990). However, Yokoyama et al. (1985) did not observe significant changes in ruminant hindgut fermentation resulting from ionophore treatment. Feeding of ionophores changes the site of digestion of dietary carbohydrate fractions. Ruminal digestion of starch may be decreased, but postruminally starch digestion is increased to the extent that total tract digestibility is unchanged (Funk et al., 1986; Muntifering et al., 1981). Fiber digestion is largely unaffected by ionophores (Allen and Harrison, 1979). Increased numbers of ionophore-resistant, fibrolytic bacteria, such as F. succinogenes may offset the reduced numbers of ionophore-sensitive ruminococci. Additionally, the somewhat longer rumen retention time caused by ionophores may contribute to maintenance of normal fiber digestion (Lemenager et al., 1978). Feeding monensin to lactating cows decreased ruminal digestion of OM, acid detergent fiber and starch (Haimoud et al., 1995). Total tract digestibility of these components was not different between control and monensin diets.
Effects of ionophores on nitrogen metabolism in the rumen: Ionophores have significant effects on nitrogen metabolism in the rumen. Research to characterize the effects of monensin on nitrogen metabolism showed that protein degradation, ammonia accumulation and microbial nitrogen were reduced in vitro with monensin (Van Nevel and Demeyer, 1977; Whetstone et al., 1981). Accumulation of α-amino nitrogen and peptides suggested that monensin had a greater inhibition on deamination than proteolysis. Net microbial growth was reduced by monensin, although overall fermentation as measured by fermentation end products was not affected (Muntifering et al., 1981; Faulkner et al., 1985). The observations of decreased ruminal protein degradation when ionophores were fed stimulated Russell’s group in the 1980s to investigate the actions of monensin on bacteria with known preference for utilizing protein as an energy substrate in the rumen ecosystem. Changes in overall nitrogen metabolism resulting from monensin feeding could not be accounted for quantitatively with the existing knowledge of ruminal bacteria activity. For example, pure cultures of rumen bacteria with the highest ammonia production from protein were gram negative and produced only 75-80% of the ammonia produced by mixed rumen bacteria. Russell hypothesized that within the rumen unknown strains of bacteria that have high specific rates of ammonia production must exist.
Using classical isolation techniques, Russell et al. (1988) were able to identify two new bacteria that had 18-39 times higher ammonia producing ability than previously known species of bacteria in the rumen. These bacteria were identified as a Peptostreptococcus species and a Clostridium species. Both were gram positive, required an amino acid source for growth and were monensin sensitive. Characterization of these two bacteria plus an additional Clostridium species is detailed in Paster et al. (1993). Using mixed ruminal bacteria in vitro, Chen and Russell (1991) demonstrated that monensin had very little effect on ruminal proteolysis but caused a reduction in ammonia production. These workers demonstrated that the reduction in ammonia in the presence of monensin occurred because of an accumulation of nitrogenous compounds in the form of non ammonia non protein nitrogen (peptides and amino acids). In a subsequent study with Holstein cows, Yang and Russell (1993a, b) demonstrated that the decrease in rumen ammonia caused by monensin was associated with a 10-fold decrease in ruminal bacteria that use amino acids and peptides as an energy source for growth.
A greater proportion of the nitrogen that reaches the abomasum is of dietary origin when monensin is included in the diet (Faulkner et al., 1985; Muntifering et al., 1981). The relative decrease in bacterial nitrogen originates from a significant reduction in ruminal bacterial protein synthesis. Efficiency of ruminal bacterial protein synthesis was generally unchanged (Faulkner et al., 1985; Muntifering et al., 1981). Flow of total essential and total nonessential amino acids to the abomasum was greater when monensin was fed with preformed protein (Faulkner et al., 1985). Quantity of amino acids reaching the abomasum was not affected by monensin when urea was the principal nitrogen source (Poos et al., 1979).
Much of the research on ionophores and protein metabolism was conducted using animals at maintenance intake or slightly above maintenance intake. Studies in lactating dairy cows have shown similar results. Haimoud et al. (1995) reported reduced rumen ammonia and reduced ruminal breakdown of dietary protein in dairy cows fed monensin. More protein of dietary origin reached the lower gut. Bacterial protein synthesis was unaffected. Total tract nitrogen digestibility was increased significantly by approximately 8% between 3 and 9 days postpartum. Nitrogen balance during this time was improved by monensin supplementation (Plaizier et al., 2000). Spears (1990) summarized the effects of ionophores on apparent digestibility of nitrogen in cattle. Improvements in nitrogen digestibility averaged about 3.5 percentage points in cattle. In most studies, retained nitrogen as a percent of nitrogen intake or as a percent of nitrogen absorbed was increased with ionophores (Muntifering et al., 1981; Poos et al., 1979). A higher proportion of plant nitrogen to microbial nitrogen reaching the small intestine may explain the improved nitrogen digestibility and retention with ionophore feeding because of the typically higher digestibility of plant nitrogen and lack of nutrient value of nucleic acid nitrogen from bacteria (Rasmussen, 1993).
Ionophores and the control of rumen-related digestive disorders: Certain animal conditions, e.g., bloat and acidosis are caused by or related to disturbances in ruminal fermentation. These conditions are attenuated or reduced when ionophores are fed because of a specific effect on a particular bacterial species, changes in eating behavior or changes in end products of fermentation.
Bloat: Bloat result from excess production of stable foam in the rumen (Bartley et al., 1983). Gas becomes entrapped within the reticulo-rumen and failure of the eructation mechanism causes acute abdominal distension. The acute nature of the disorder often leads to death within hours after the ingestion of the bloat provocative meal. Viscosity of rumen fluid from bloat susceptible steers was higher than in normal steers fed a feedlot type diet (Sakauchi and Hoshino, 1981). Incidence of bloat was 86.3% in bloat susceptible steers during a 30 days control feeding period. Subsequent feeding of monensin (40 mg kg–1) reduced the incidence of bloat to 4.2% over the following 36 days. Removal of monensin from the diet caused incidence of bloat to increase to 24.3% for the next 36 days. During the feeding of monensin, viscosity of rumen fluid from bloat-susceptible steers decreased to values similar to normal steers. Similar effects of monensin on viscosity of rumen fluid were noted in sheep (Usagawa, 1992).
Acidosis: Current levels of milk production and the relative cost of grain to forage have resulted in an increased proportion of rapidly fermentable carbohydrate in the diet of dairy cows. By default, effective fiber in the diet is often deficient. Consumption of rapidly fermentable diets places the dairy cow at risk for acidosis. Ionophores have the potential to ameliorate the disease of acidosis by two distinct mechanisms.
The first mechanism is through ionophore effects on lactic acid producing strains of bacteria such as Streptococcus bovis. Dennis et al. (1981) reported that lasalocid and monensin inhibit many of the major strains of rumen bacteria that produce lactic acid. They also reported that the major strains of lactate fermenting bacteria were resistant to ionophores. Colony counts of S. bovis and Lactobacillus (lactate producing gram positive bacteria) were reduced in rumen fluid taken from cattle infused intraruminally with glucose and ionophore. Conversely, colony counts of lactate utilizing bacteria (gram negative) were unaffected by presence of ionophore.
The second mechanism that ionophores may impact acidosis is through changes in eating dynamics of cattle fed ionophore containing diets. Ionophores change the eating behavior of cattle and this may cause a reduction in digestive conditions, including acidosis and death (Black and McQuilken, 1980; Cooper et al., 1999).
Rumen microorganisms and their sensitivity-resistant to ionophores: Soon after antibiotics became commercially available, resistant bacterial mutants were observed. While the array of antibiotics is great, resistance can be categorized by three general schemes (Chopra and Ball, 1982): (i) Synthesis of enzymes which degrade the antibiotic (e.g., 1-lactamases), (ii) Alteration in the cellular target (e.g., ribosomes) and (iii) Change in cellular permeability (e.g., streptomycin uptake). Because genes encoding resistance factors often are readily transferred from one strain (or species) to another, the effectiveness of many antibiotic treatments has been reduced. Yet after more than 30 years of widespread use, ionophores continue to improve the efficiency of animal performance. This observation would suggest that the sensitivity of ruminal microorganisms is relatively stable and that the pattern of resistance is due to a fundamental difference between cells.
Major rumen organisms which have been reported to be resistant or sensitive to rumensin are presented in Fig. 3. The sensitive bacteria are predominantly Gram+ and are acetic acid, butyric acid, lactic acid and methane producers. The monensin-resistant bacteria are Gram- and tend to be succinate and propionate producers.
The differences between rumensin-sensitive and rumensin-resistant bacteria are probably largely due to the inherent differences between Gram‾ and Gram+ organisms. The outer membrane of gram negative bacteria is impermeable to many macromolecules and solute movement is mediated by porins. Porins form hydrophilic channels through the hydrophobic outer membrane with an exclusion limit of approximately 600 daltons (Nikaido and Nakae, 1979). Because ionophores are extremely hydrophobic and have molecular sizes of greater than 500 daltons, the outer membrane should serve as a protective barrier. Indeed, gram negative bacteria generally are ionophore resistant. Gram positive bacteria, which lack a protective outer membrane, are usually sensitive to ionophores. Some bacteria which stain gram negative actually have gram positive cell wall structure (e.g., Butyrivibrio fibrisolvens (Cheng and Costerton, 1977) and ruminococci (Hungate, 1966). These organisms are also sensitive to ionophores. There have been relatively few enumerations of monensin-sensitive bacteria in animals fed monensin (Dawson and Boling, 1983), but in some cases the numbers of gram positive species decreased (Nagaraja et al., 1982). Protozoa and fungi lack an outer membrane and in vitro, they are also sensitive to monensin (Dennis et al., 1986; Hino, 1981; Stewart et al., 1987).
 | |
| Fig. 3: | Response of rumen bacteria to rumensin |
While the hypothesis that the bacterial outer membrane provides a general pattern of ionophore resistance is attractive, there are several observations which need to be reconciled: (i) Some gram negative species are not resistant to high concentrations of ionophores (Dawson and Boling, 1987; Nagaraja and Taylor, 1987), (ii) Ionophores increased ion flux in some gram negative bacteria (Dawson and Boling, 1987; Nagaraja and Taylor, 1987), (iii) Gram negative species that were originally sensitive to ionophores can adapt (Newbold et al., 1988), (iv) There have been reports that gram positive bacteria can develop resistance to ionophores (Dawson and Boling, 1987; Dennis et al., 1981; Liu, 1982), (v) Certain ciliate protozoa (Newbold et al., 1988) and fungi (Stewart et al., 1987) were relatively insensitive to ionophores. Hence, the presence of an outer membrane is not an absolute criterion for resistance.
At this time, it is impossible to exclude the involvement of extracellular polysaccharide in ionophore resistance. Because ionophore sensitivity is related to ion movement, increased ion pump activity might also provide a mechanism of resistance. The deposition of polysaccharide does not entail large energy expenditure (Stouthamer, 1973), but mechanisms to counteract futile ion cycles could be costly. Since the rumen is a highly competitive and usually energy-limited environment, the survival of resistant organisms in vitro would not necessarily guarantee their persistence in vivo.
CONCLUSION
Ionophores are organic compounds mainly from Streptomyces spp. that facilitate selective transportation of ions across the outer cell membrane. The primary way in which ionophores modify rumen function is by decreasing the ruminal population of gram positive bacteria relative to that of gram negative bacteria. Gram positive bacteria lack the complex cell wall of gram negative bacteria and the associated lipopolysaccharide layer with its protein channels (porins) that have a size exclusion limit (600 Da) that is impervious to ionophores. Consequently, ionophores successfully infiltrate the outer cell membrane of gram positive bacteria and rapidly and repeatedly cause efflux of intracellular K+ from the cell and influx of extracellular protons (Na+ and H+). To overcome the resulting acidity and the depletion of K+, which inhibits protein synthesis, ATPase pumps are elicited to eject the protons but this depletes energy reserves for bacterial growth. The cytoplasmic acidity culminates in cell death.
The metabolic effects of ionophores on the rumen fermentation can be summarized in this point:
| • | Shift in acetate-propionate ratio toward more propionate |
| • | Some increase of lactate to propionate production via the accelerate pathway |
| • | Decreased ruminal protein breakdown and deamination; lower ruminal ammonia-N |
| • | Primary H+ or formate producers, gram positive organisms, are inhibited |
| • | Decrease in methane production primarily due to lowered availability of H2 and formate and depressed interspecies H2 transfer |
| • | Depression of lactic acid production under acidosis inducing conditions |
| • | Gram negative organisms, of which many produce succinate (source of propionate) or possess capacity for the reductive tricarboxylic acid cycle to use bacterial reducing power, survive |
| • | Some evidence for depressed rumen content turnover |
| • | A mild inhibition of protozoa |
| • | Decrease in rumen fluid viscosity in bloated animals |
| • | Depressed growth yield efficiency of the ruminal microbes |
REFERENCES
- Haimoud, D.A., M. Vernay, C. Bayourthe and R. Moncoulon, 1995. Avoparcin and monensin effects on the digestion of nutrients in dairy cows fed a mixed diet. Can. J. Anim. Sci., 75: 379-385.
CrossRefDirect Link - Azzaz, H.H., H.M. Ebeid, T.A. Morsy and S.M. Kholif, 2015. Impact of feeding yeast culture or yeast culture and propionibacteria 169 on the productive performance of lactating buffaloes. Int. J. Dairy Sci., 10: 107-116.
CrossRefDirect Link - Azzaz, H.H., H.A. Aziz, E.S.A. Farahat and H.A. Murad, 2015. Impact of microbial feed supplements on the productive performance of lactating nubian goats. Global Vet., 14: 567-575.
CrossRefDirect Link - Azzaz, H.H., A.M. Kholif, H.A. Murad, M.A. Hanfy and M.H. Abdel Gawad, 2012. Utilization of cellulolytic enzymes to improve the nutritive value of banana wastes and performance of lactating goats. Asian J. Anim. Vet. Adv., 7: 664-673.
CrossRefDirect Link - Azzaz, H.H., H.A. Murad, A.M. Kholif, T.A. Morsy, A.M. Mansour and H.M. El-Sayed, 2013. Increasing nutrients bioavailability by using fibrolytic enzymes in dairy buffaloes feeding. J. Biol. Sci., 13: 234-241.
CrossRefDirect Link - Bartley, E.E., T.G. Nagaraja, E.S. Pressman, A.D. Dayton, M.P. Katz and L.P. Fina, 1983. Effects of lasalocid or monensin on legume or grain (FeedLot) bloat. J. Anim. Sci., 56: 1400-1406.
PubMedDirect Link - Bergen, W.G. and D.B. Bates, 1984. Ionophores: Their effect on production efficiency and mode of action. J. Anim. Sci., 58: 1465-1483.
PubMedDirect Link - Booth, I.R., 1985. Regulation of cytoplasmic pH in bacteria. Microbiol. Rev., 49: 359-378.
Direct Link - Chen, G.J. and J.B. Russell, 1991. Effect of monensin and a protonophore on protein degradation, peptide accumulation and deamination by mixed ruminal microorganisms in vitro. J. Anim. Sci., 69: 2196-2203.
PubMedDirect Link - Chen, M. and M.J. Wolin, 1979. Effect of monensin and lasalocid-sodium on the growth of methanogenic and rumen saccharolytic bacteria. Applied Environ. Microbiol., 38: 72-77.
PubMedDirect Link - Cheng, K.J. and J.W. Costerton, 1977. Ultrastructure of Butyrivibrio fibrisolvens: A gram-positive bacterium? J. Bacteriol., 129: 1506-1512.
Direct Link - Chopra, I. and P. Ball, 1982. Transport of antibiotics into bacteria. Adv. Microbiol. Physiol., 23: 183-240.
PubMedDirect Link - Chow, J.M. and J.B. Russell, 1992. Effect of pH and monensin on glucose transport by Fibrobacter succinogenes, a cellulolytic ruminal bacterium. Applied Environ. Microbiol., 58: 1115-1120.
Direct Link - Chow, J.M., J.A. van Kessel and J.B. Russell, 1994. Binding of radiolabeled monensin and lasalocid to ruminal microorganisms and feed. J. Anim. Sci., 72: 1630-1635.
PubMedDirect Link - Cooper, R.J., T.J. Klopfenstein, R.A. Stock, C.T. Milton, D.W. Herold and J.C. Parrott, 1999. Effects of imposed feed intake variation on acidosis and performance of finishing steers. J. Anim. Sci., 77: 1093-1099.
PubMedDirect Link - Dawson, K.A. and J.A. Boling, 1983. Monensin-resistant bacteria in the rumens of calves on monensin-containing and unmedicated diets. Applied Environ. Microbiol., 46: 160-164.
Direct Link - Dawson, K.A. and J.A. Boling, 1987. Effects of potassium ion concentrations on the antimicrobial activities of ionophores against ruminal anaerobes. Applied Environ. Microbiol., 53: 2363-2367.
Direct Link - Dennis, S.M., T.G. Nagaraja and A.D. Dayton, 1986. Effect of lasalocid, monensin and thiopeptin on rumen protozoa. Res. Vet. Sci., 41: 251-256.
PubMedDirect Link - Dennis, S.M., T.G. Nagaraja and E.E. Bartley, 1981. Effects of lasalocid or monensin on lactate-producing or -using rumen bacteria. J. Anim. Sci., 52: 418-426.
PubMedDirect Link - Faulkner, D.B., T.J. Klopfenstein, T.N. Trotter and R.A. Britton, 1985. Moneimsin effects on digestibility, ruminal protein escape and microbial protein synthesis on high-fiber diets. J. Anim. Sci., 61: 654-660.
Direct Link - Funk, M.A., M.L. Galyean and T.T. Ross, 1986. Potassium and lasalocid effects on performance and digestion in lambs. J. Anim. Sci., 63: 685-691.
PubMedDirect Link - Lemenager, R.P., F.N. Owens, B.J. Shockey, K. S. Lusby and R. Totusek, 1978. Monensin effects on rumen turnover rate, twenty-four hour VFA pattern, nitrogen components and cellulose disappearance. J. Anim. Sci., 47: 255-261.
Direct Link - Marounek, M. and B. Hodrova, 1989. Susceptibility and resistance of anaerobic rumen fungi to antimicrobial feed additives. Lett. Applied Microbiol., 9: 173-175.
CrossRefDirect Link - Marounek, M., O. Petr and L. Machanova, 1990. Effect of monensin on in vitro fermentation of maize starch by hindgut contents of cattle. J. Agric. Sci., 115: 389-392.
CrossRefDirect Link - McGuffey, R.K., L.F. Richardson and J.I.D. Wilkinson, 2001. Ionophores for dairy cattle: Current status and future outlook. J. Dairy Sci., 84: E194-E203.
CrossRefDirect Link - Muntifering, R.B., B. Theurer and T.H. Noon, 1981. Effects of monensin on site and extent of whole corn digestion and bacterial protein synthesis in beef steers. J. Anim. Sci., 53: 1565-1573.
PubMedDirect Link - Murad, H.A. and H.H. Azzaz, 2013. Cellulase production from rice straw by Aspergillus flavus NRRL 5521. Sci. Int., 1: 103-107.
CrossRefDirect Link - Nagaraja, T.G. and M.B. Taylor, 1987. Susceptibility and resistance of ruminal bacteria to antimicrobial feed additives. Applied Environ. Microbiol., 53: 1620-1625.
Direct Link - Nagaraja, T.G., T.B. Avery, E.E. Bartley, S.J. Galitzer and A.D. Dayton, 1982. Effect of lasalocid, monensin or thiopeptin on lactic acidosis in cattle. J. Anim. Sci., 54: 649-658.
CrossRefPubMedDirect Link - Newbold, C.J., R.J. Wallace, N.D. Watt and A.J. Richardson, 1988. Effect of the novel ionophore tetronasin (ICI 139603) on ruminal microorganisms. Applied Environ. Microbiol., 54: 544-547.
Direct Link - Nikaido, H. and T. Nakae, 1979. The outer membrane of Gram-negative bacteria. Adv. Microb. Physiol., 20: 163-250.
CrossRefDirect Link - Ovchinnikov, Y.A., 1979. Physico-chemical basis of ion transport through biological membranes: Ionophores and ion channels. Eur. J. Biochem., 94: 321-336.
Direct Link - Painter, G.R., R. Pollack and B.C. Pressman, 1982. Conformational dynamics of the carboxylic ionophore lasalocid A underlying cation complexation-decomplexation and membrane transport. Biochemistry, 21: 5613-5620.
CrossRefDirect Link - Paster, B.J., J.B. Russell, C.M.J. Yang, J.M. Chow, C.R. Woese and R. Tanner, 1993. Phylogeny of the ammonia-producing ruminal bacteria Peptostreptococcus anaerobius, Clostridium sticklandii and Clostridium aminophilum sp. nov. Int. J. Syst. Bacteriol., 43: 107-110.
CrossRefDirect Link - Plaizier, J.C., A. Martin, T. Duffield, R. Bagg, P. Dick and B.W. McBride, 2000. Effect of a prepartum administration of monensin in a controlled-release capsule on apparent digestibilities and nitrogen utilization in transition dairy cows. J. Dairy Sci., 83: 2918-2925.
CrossRefPubMedDirect Link - Poos, M.I., T.L. Hanson and T.J. Klopfenstein, 1979. Monensin effects on diet digestibility, ruminal protein bypass and microbial protein synthesis. J. Anim. Sci., 8: 1516-1524.
PubMedDirect Link - Pressman, B.C., 1976. Biological applications of ionophores. Ann. Rev. Biochem., 45: 501-530.
CrossRefDirect Link - Rasmussen, M.A., 1993. Isolation and characterization of Selenomonas ruminantium strains capable of 2-deoxyribose utilization. Applied Environ. Microbiol., 39: 2077-2081.
Direct Link - Richardson, L.E., A.P. Raun, E.L. Potter, C.O. Cooley and R.P. Rathmacher, 1976. Effect of monensin on rumen fermentation in vitro and in vivo. J. Anim. Sci., 43: 657-664.
Direct Link - Rogers, J.A. and C.L. Davis, 1982. Rumen volatile fatty acid production and nutrient utilization in steers fed a diet supplemented with sodium bicarbonate and monensin. J. Dairy Sci., 65: 944-952.
CrossRefPubMedDirect Link - Rosen, B.P., 1986. Recent advances in bacterial ion transport. Ann. Rev. Microbiol., 40: 263-286.
CrossRefDirect Link - Russell, J.B., H.J. Strobel and G.J. Chen, 1988. Enrichment and isolation of a ruminal bacterium with a very high specific activity of ammonia production. Applied Environ. Microbiol., 54: 872-877.
Direct Link - Russell, J.B., 1987. A proposed mechanism of monensin action in inhibiting ruminal bacterial growth: effects on ion flux and protonmotive force. J. Anim. Sci., 64: 1519-1525.
PubMedDirect Link - Russell, J.B. and H.J. Stroble, 1989. Effect of Ionophores on ruminal fermentation. Applied Environ. Microbiol., 55: 1-6.
Direct Link - Sakauchi, R. and S. Hoshino, 1981. Effects of monensin on ruminal fluid viscosity, pH, volatile fatty acids and ammonia levels and microbial activity and population in healthy and bloated feedlot steers. Zeitschrift Tierphysiologie Tierernahrung Futtermittelkunde, 46: 21-32.
CrossRefDirect Link - Spears, J.W., 1990. Ionophores and nutrient digestion and absorption in ruminants. J. Nutr., 120: 632-638.
PubMedDirect Link - Stewart, C.S., S.H. Duncan and K.N. Joblin, 1987. Antibiotic Manipulation of Rumen Microflora. The Effects of Avoparcin and Monensin on the Release of Tritium from Labelled Cellulose by Bacteroides succinogenes and the Rumen Fungus Neocallimastix frontalis. In: Recent Advances in Anaerobic Bacteriology: Proceedings of the fourth Anaerobic Discussion Group Symposium held at Churchill College, University of Cambridge, July 26-28, 1985, Borriello, S.P., J.M. Hardie, B.S. Drasar, B.I. Duerden, M.J. Hudson and R.J. Lysons (Eds.). Springer, Netherlands, ISBN-13: 978-94-009-3293-7, pp: 108-119.
- Stouthamer, A.H., 1973. A theoretical study on the amount of ATP required for synthesis of microbial cell material. Antonie Van Leeuwenhoek, 39: 545-565.
CrossRefDirect Link - Usagawa, T., 1992. Effects of monensin and salinomycin on the in vitro foam stability of sheep rumen fluid. Anim. Sci. Technol., 63: 16-20.
Direct Link - Van Kessel, J.S. and J.B. Russell, 1992. Energetics of arginine and lysine transport by whole cells and membrane vesicles of strain SR, a monensin-sensitive ruminal bacterium. Applied Environ. Microbiol., 58: 969-975.
PubMedDirect Link - Van Nevel, C.J. and D.I. Demeyer, 1977. Effect of monensin on rumen metabolism in vitro. Applied Environ. Microbiol., 34: 251-257.
Direct Link - Whetstone, H.D., C.L. Davis and M.P. Bryant, 1981. Effect of monensin on breakdown of protein by ruminal microorganisms in vitro. J. Am. Sci., 53: 803-809.
PubMedDirect Link - Yang, C.M. and J.B. Russell, 1993. Effect of monensin on the specific activity of ammonia production by ruminal bacteria and disappearance of amino nitrogen from the rumen. Applied Environ. Microbiol., 59: 3250-3254.
Direct Link - Yang, C.M. and J.B. Russell, 1993. The effect of monensin supplementation on ruminal ammonia accumulation in vivo and the numbers of amino acid-fermenting bacteria. J. Anim. Sci., 71: 3470-3476.
PubMedDirect Link - Yang, W.Z., C. Benchaar, B.N. Ametaj, A.V. Chaves, M.L. He and T.A. McAllister, 2007. Effects of garlic and juniper berry essential oils on ruminal fermentation and on the site and extent of digestion in lactating cows. J. Dairy Sci., 90: 5671-5681.
CrossRefPubMedDirect Link - Morsy, T.A., S.M. Kholif, O.H. Matloup, M.M. Abdo and M.H. El-Shafie, 2012. Impact of anise, clove and juniper oils as feed additives on the productive performance of lactating goats. Int. J. Dairy Sci., 7: 20-28.
CrossRef