
Irma Isnafia Arief
Department of Animal Production and Technology, Faculty of Animal Science, Bogor Agricultural University, Indonesia
Betty Sri Laksmi Jenie
Department of Food Science and Technology, Faculty of Agricultural Technology, Bogor Agricultural University, Kampus IPB Darmaga Bogor, Bogor, 16680, Indonesia
Made Astawan
Department of Food Science and Technology, Faculty of Agricultural Technology, Bogor Agricultural University, Kampus IPB Darmaga Bogor, Bogor, 16680, Indonesia
Kazuhito Fujiyama
International Center for Biotechnology, Osaka University, Japan 2-1, Yamadaoka, Suita, Osaka 565-0871, Japan
Arief Budi Witarto
Faculty of Biotechnology, Sumbawa University of Technology, Jl. Raya Olat Maras, Moyo Hulu, Kab. Sumbawa Besar, Nusa Tenggara Barat, Indonesia
Asian Journal of Animal Sciences
Year: 2015 | Volume: 9 | Issue: 1 | Page No.: 25-36







ABSTRACT
Functional foods are food that can improve health by providing benefit beyond that of the basic nutritional value. Probiotics are included as component of functional foods. Probiotics are living microorganisms which when administered in adequate amounts confer a healthy benefit on the host. The research aim was to identify and characterize as probiotic of Indonesian Lactic Acid Bacteria (LAB) that isolated from fresh beef derived from traditional market around Bogor, West Java, Indonesia. The results showed that the 16S rRNA gene could be amplified by PCR (Polymerase Chain Reaction) to get single band with designed primer. Identification by 16S rRNA gene sequencing of 20 isolates showed that 11 isolates were identified as Lactobacillus plantarum, 6 isolates as Lactobacillus acidophilus, 2 isolates as Pediococcus pentosaceus and 1 isolate as Enterococcus faecium. The twenty LAB isolates showed different ability to survive under simulated gastrointestinal conditions. Only 10 isolates could survive well at pH 2.0 (survival rate over than 50%) and the population increased at pH 2.5, 3.2 and 7.2. The ten LAB isolates i.e., 7 strains of L. plantarum 1B1, 1A5, 2D1, 1C4, 2B1, 2B2, 2C12 and 3 strains of L. acidophilus 2B4, 2C2 and 1A32 demonstrated other important probiotic characteristics including survival ability at 0.5% bile salt, antibacterial activities and had good coaggregationcapabilities against the tested pathogenic bacteria (Escherichia coli ATCC 25922, Salmonella typhimurium ATCC 14028, enteropathogenic Escherichia coli/EPEC and Staphylococcus aureus ATCC 25923) by in vitro analysis.
PDF Abstract XML References Citation
Received: June 10, 2014;
Accepted: December 12, 2014;
Published: January 24, 2015
How to cite this article
Irma Isnafia Arief, Betty Sri Laksmi Jenie, Made Astawan, Kazuhito Fujiyama and Arief Budi Witarto, 2015. Identification and Probiotic Characteristics of Lactic Acid Bacteria Isolated from Indonesian Local Beef. Asian Journal of Animal Sciences, 9: 25-36.
DOI: 10.3923/ajas.2015.25.36
URL: https://scialert.net/abstract/?doi=ajas.2015.25.36
DOI: 10.3923/ajas.2015.25.36
URL: https://scialert.net/abstract/?doi=ajas.2015.25.36
INTRODUCTION
Recently, development of Lactic Acid Bacteria (LAB) as culture starter for functional food currently is a trend in food processing technology. Lactic acid bacteria as probiotic is beneficial for human health. Probiotic defined as additional nutrient consisted of living microorganism that if it was consumed by human or animal in sufficient amount will confer health benefit to the host (FAO. and WHO., 2002). Probiotics can be defined as live microbial food supplements with health benefits to the host by improving the intestinal microbiota. Many reports showed the usefulness of LAB as probiotics for human (Brady et al., 2000; Herich and Levkut, 2002; Kimoto-Nira et al., 2007; Zoumpopoulou et al., 2008). It has been reported that basic conditions for LAB strains to be used as probiotics minimally include the following: (1) They should be generally recognized as safe (GRAS), (2) They should be tolerant to acid and bile and (3) They have antagonistic activity against bacterial pathogenics (FAO. and WHO., 2002; Pennacchia et al., 2006).
Beef can become source of LAB. Peranakan Ongole (PO) beef is Indonesian local beef that domesticated in Indonesia and mostly is consumed by Indonesian people. PO beef is sold at many traditional market in Indonesia. The need for new strains of LAB that carry probiotic characteristics for development of probiotic food is increasing, recently. Exploration of functional characteristic of LAB isolated from Indonesian local beef is interesting. Further research is needed to find out more information related to functional characteristics as probiotic, especially to identify accurate species and strain from isolated LAB in order to meet FAO. and WHO (2002) requirements. Molecular identification of LAB by Polymerase Chain Reaction (PCR) followed by 16S rRNA genes sequencing is well known method that can be applied for this purpose. The aims of this study were (a) To identify isolates of indigenous LAB isolated from Indonesian local beef (PO beef) and (b) To characterize the strains as probiotic by in vitro analysis.
MATERIALS AND METHODS
Isolation and identification of LAB
LAB isolation, morphological, physiological and biochemical tests: To isolate the LAB, 25 g of each beef sample were taken aseptically from traditional markets at Bogor, West Java, Indonesia, diluted in 225 mL buffer peptone water (BPW, Oxoid) and homogenized by stomacher blender. Serial 10-fold dilution of both beef samples were prepared with buffer peptone water (BPW, Oxoid) and inoculated on de Man Rogosa Sharp agar (MRS-agar, Oxoid). All media were incubated for 48 h at 37°C. About 50% of typical colonies from each medium were randomly selected and streaked onto MRS agar media to obtain pure cultures.
Gram staining, morphology, catalase activity, NH3 and gas production from glucose were determined as described by Cai et al. (1999). Growth at different temperatures was observed in MRS broth after incubation at 15 and 45°C for 48 h. Salt tolerance of LAB was tested by using MRS broth containing 6.5% (w/v) NaCl at 37°C for 48 h. For testing the sugar fermentations, strains were cultivated in different media containing sugar.
Identification of LAB by polymerase chain reaction and 16S rRNA genes sequencing genome extraction: Extraction of genome was performed using kits from Promega, the Wizard ® SV Genomic Purification System Catalog No. A2360. Genomic extracts obtained was analyzed for its concentration of genomic DNA using a spectrophotometer (Gene Quant) at 260 and 280 nm.
Amplification of 16S rRNA genes by PCR: 16S rRNA gene amplification procedure was performed according to the method of Islam et al. (2008), with slight modification on the PCR condition. Briefly, reaction amplification of DNA samples were carried out in 0.2 mL PCR tubes. Each PCR reaction tube contained Eq Takara Taq (5 units mL-1) of 0.25 mL, 10 x Eq Taq Buffer 5 mL, dNTP mixture (2.5 mM) of 4 mL, universal Primer 9 F (5'-GAGTTTGATCCTGGCTCAG-3') and universal primer 1541 R (5'AGGAGGTGATCCAGCC-3') of 2.5 mL each of 20 μM, in increments of 1 mg of genomic extract and distilled water was added until 50 mL of volume. Primers were purchased from the company’s primary Gene Design, Inc. Japan. The PCR amplification was performed to obtain a single band which was the 16S rRNA gene amplification. In this study, there were three conditions for the PCR reaction as follows: (1) 15 isolates (1C6, 1D1, 1A6, 2C12, 2B2, 2B4, 1A5, 1A32, 1B1, 2B1, 1C4, 2D1, 2B3, 1C3 and 1C1) were treated with PCR conditions of initial denaturation of 95°C 5 min, 30 cycles of annealing and extension at 95°C for 1 min, 50°C for 1 min, 72°C for 2 min and 72°C for 7 min final stages. (2) Three isolates (2D2, 1A1, 2A2) with PCR conditions of initial denaturation at 95°C for 5 min, followed by 30 cycles of 95°C for 1 min and 45 sec 51°C, 72°C for 2 min and the final stage 72°C for 7 min. (3) Two isolates (2C2 and 1A2) with PCR conditions of initial denaturation at 95°C for 5 min, 30 cycles of 95°C for 1 min, 45 sec 52°C, 72°C for 2 min and 72°C for 7 min final stage. The PCR products were examined by using 1% agarose electrophoresis and photographed using a UV camera and the results were printed on Polaroid film.
16S rRNA gene sequencing: Primers were universal primers 785F (5'-GGATTAGATACCCTGGTAGTC-3') with the amplicon in bases 800-1400 and 802 R (5'-TACCAGGGTATCTAATCC-3') with amplicon bases to 1-800, by Weisburg et al. (1991) with slight modification on primer sequences. Additional primer sequences were Primer 1R (5'-GGGCATGATGATTTGACGTC-3') for amplifying sequence of base number 600-1200, 2F primer (5'-GTGAGACTGCCGGTGACAAA-3') for the 1150-1500 base amplicon and primer 3R (5'-ATCAGACTTAAAAAACCGCC-3') for the 1-600 base amplicon to get a complete 16S rRNA gene sequence. Gene sequencing were performed using the purified extract genomic DNA Purification Kit Monofast I (protocol according to company instructions). Big Dye sequencing reagent was ver 3.1. Terminator Cycle Sequencing Kit (Applied Biosystem, Japan). Sequencer tool used was the ABI Prism 3100 Genetic Analyzer (Applied Biosystem, Japan).
Sequence similarity searches were performed in the GenBank data library using the BLAST program. The sequence information was then imported into Genetic 7 software program for assembly and CLUSTAL W software program for alignment. Phylogenetic trees were constructed by the neighbor-joining method using MEGA 4 software program.
Characterization of LAB as probiotic by in vitro analysis
Bacterial strains and media: The twenty isolates of LAB were preserved at -30°C in MRS broth (Oxoid) with 20% (v/v) glycerol added. Before experimental use, the cultures were propagated twice in MRS (Oxoid) at 37°C.
The indicator strains used are including Salmonella typhimurium ATCC 14028, Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 25923 and local isolate enteropathogenic Escherichia coli (EPEC). The EPEC strain was collection of Faculty of Veterinary Bogor Agricultural University, Indonesia.
Survival rates under simulated gastrointestinal conditions: This experiments had been done according to Lin et al. (2006). Three samples were assayed as replication. The initial population also were determined. Survival rates were counted by comparing between initial population and end population after incubation on different pH conditions. Effects of bile salt on the growth of LAB were observed by inoculating 1 mL of LAB in 9 mL MRS-broth containing 0.5% bile salt (Difco). They were incubated at 37°C for 6 h. Viable bacterial counts were determined by plating serial dilution with BPW on MRS-agar at 37°C for 48 h.
Antibacterial activity assay: The antibacterial activity was assayed using the agar well diffusion method described by Savadogo et al. (2004). Each assay was performed in triplicate.
Coaggregation capacity assay: Coaggregation capacity of LAB against pathogenic bacteria was performed according to the method of El-Naggar (2004). Experiments were conducted in three replication.
Statistical analysis: Experimental design used was Complete Randomized Design. Data was analyzed by ANOVA (Analysis of variance) (Steel et al., 1996). If significantly difference, tukey test was used.
RESULT AND DISCUSSION
Isolation and identification of LAB
Morphological, physiological and biochemical profile of LAB: The morphological, physiological and biochemical profile of LAB are shown in Table 1. All strains are Gram positive, catalase negative and homofermentatives.
Identification of LAB by polymerase chain reaction and 16S rRNA genes sequencing: All genome extracts had a value of genome comparison of absorbance 260/280 around 1.8 representing that it contained DNA with high purity. Gene amplification by PCR and 1% agarose electrophoresis managed to obtain a single band indicating that the 16S rRNA gene successfully amplified with primers 9F and 1541R (Fig. 1).
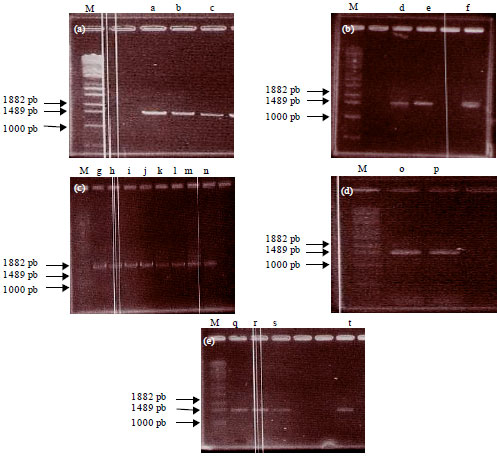 | |
| Fig. 1(a-e): | 1% agarose electrophoresis and 16S rRNA gene amplification by PCR (M = marker λEcoT141, a = 1C6, b = 1D1, c = 1A6, d = 2D2, e = 2A2, f = 1B1, g = 2B2, h = 2D1, i = 1A32, j = 2C12, k = 2B4, l = 1A1, m = 1C4, n = 2C2, o = 2C2, p = 1A2, q = 2B1, r = 2B3, s = 1C3, t = 1C1) |
| Table 1: | Morphological, physiological and biochemical profile of LAB |
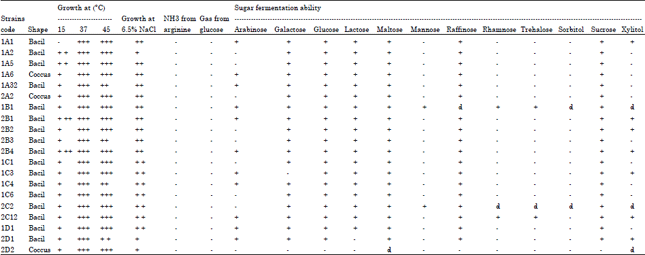 | |
| +: Weak (can ferment sugar), ++: Medium , +++: Strong, -: Can not produce NH3/gas/ferment sugar, d: Dubius (weak fermentation on sugar) | |
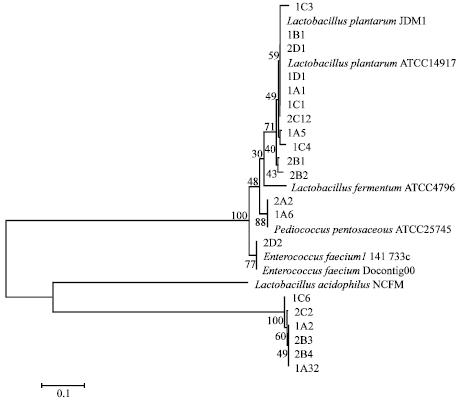 | |
| Fig. 2: | Phylogenetic tree Indonesian LAB isolates aligned with the closest international isolates (GenBank) |
Construction of phylogenetic trees showed that 20 LAB isolates were included in 4 species belonging to the Lactobacillus plantarum, Lactobacillus acidophilus, Enterococcus faecium and Pediococcus pentosaceus (Fig. 2).
A total of 11 isolates, namely 1C3, 1B1, 2D1, 1D1, 1A1, 1C1, 2C12, 1A5, 1C4, 2B1 and 2B2, classified in one cluster with Lactobacillus plantarum ATCC 14917 and Lactobacillus plantarum ATCC JDM1 and based on NCBI BLAST proximity of 97-98%. The 2A2 and 1A6 isolates were included in a cluster with Pediococcus pentosaceus ATCC 25745 proximity relationship was based on NCBI BLAST of 97%. The 2D2 isolate were included in the cluster Enterococcus faecium 1141733c and had close relations according to the NCBI BLAST of 97%. Six other isolates namely 1C6, 2C2, 1A2, 2B3, 2B4 and 1A32 were included in a cluster with Lactobacillus acidophilus NCFM and have proximity based on NCBI BLAST at 100%.
Characterization of LAB as probiotic by in vitro analysis
Survival rates under simulated gastrointestinal conditions: Gastrointestinal tract is the major location to affect the viability of LAB cells. Lin et al. (2006) reported that the stabilities of LAB cells examined either by in vivo or in vitro study were similar. Acid tolerance is accepted as one of the desirable properties used to select potentially probiotic strains. The survival of LAB strains at pH 2.0, 2.5, 3.2 (described stomach conditions) and 7.2 (described ileum and colon condition) were observed for 3 h (Table 2).
The results showed that viability of LAB were significantly affected by the acidity (Table 2). It was described that 10 isolates of LAB could survive on the stomach conditions but their ability were different and only five strains had good survival rates on pH 2.0. Strains of LAB have different capabilities on acidity survival (Table 2). It was affected by specific homeostatis. The three main system involved in LAB pH homeostatis i.e., Arginine deiminase (ADI) system, the H+-ATPase proton pump and the Glutamate decarboxylase (GAD) system (Cotter and Hill, 2003).
| Table 2: | Survival rates of LAB at different pH value |
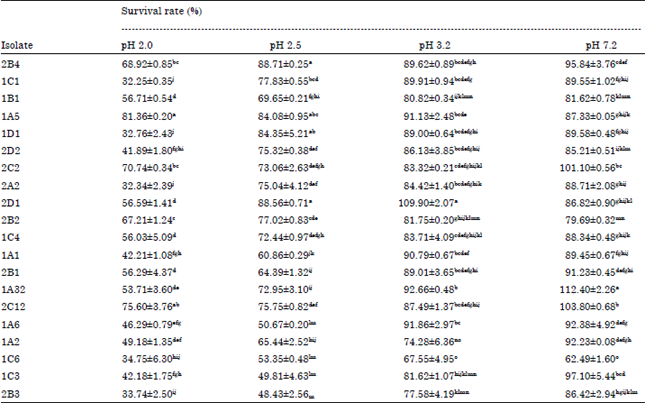 | |
| Treatments with different letters are different at p<0.05 | |
Initial population of all LAB isolates used ranged from 109-1011 CFU mL-1. On the condition of pH 2.0 for 3 h, survival of indigenous LAB isolates ranged from 32.34-75.60%. Ten isolates (2B4, 1B1, 1A5, 2C2, 2D1, 2B2, 1C4, 2B1, 1A32 and 2C12) had survival rates in conditions of pH 2.0 greater than 50%.
Survival percentage of all LAB isolates at pH 2.5 were better than pH 2 which was above 40%, with a range of 47.13-88.71%. Survival rates of LAB isolates at pH 3.2 were even better than pH 2.5. All isolates were able to survive with the percentage survival rates above 60%, with a range of 67.55-109.90%. The ten isolates that were able to survive at pH 2.0 and 2.5, showed better survival at pH 3.2, above 80%, some of them were able to survive over than 100% at pH 7.2. This suggests that the ten isolates survived at pH 2.0 is also able to survive well in higher pH until pH 7.2. The ten isolates were selected and carried out for further characterization as probiotics on subsequent testing (Table 2). In similar studies, Lin et al. (2006) reported that Lactobacillus bulgaricus and Streptococcus thermophilus in yogurt product could survive at pH 2.5 but not at pH 2.0. Mishra and Prasad (2005) reported that survival of Lactobacillus casei pH 3 was promising for all strains but not at pH 2.
The bile in human intestine is also an important factor which affected the LAB viability. Although, the composition of human bile juice is not exactly the same as 0.5% bile salt solution, most studies used bile salt as one substituted for human bile because of their similarity (Mishra and Prasad, 2005; Marteau et al., 1997). In this study, all strains that could survive well on pH 2, also could survive at 0.5% bile salt (Fig. 3).
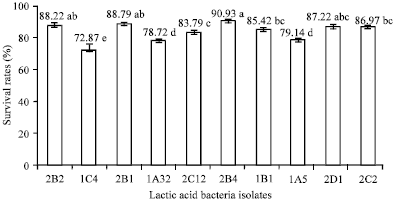 | |
| Fig. 3: | Survival rates of LAB at 0.5% bile salt treatments with different letters are different at p<0.05 |
The survival rates at 0.5% bile salt of all isolates were greater than 70%. In this study, survival rates counted by comparing initial population and residual population after incubating in the bile-salt solution. The initial populations of 10 LAB isolates were 108 CFU mL-1. The variation in bile sensitivity observed in this study was consistent with many previous reports, including Mishra and Prasad (2005) and Lin et al. (2006). Emerges that wide variation existed in the susceptibility of probiotic cultures to bile and this property was species, as well as strain specific (Pan et al., 2009). LAB could hydrolyse bile salt with Bile Salt Hydrolase enzyme (BSH), decreasing their solubility and thus weakening their detergent effect (Moser and Savage, 2001; Begley et al., 2006; Erkkila and Petaja, 2000).
Antibacterial activity assay: All LAB strains had antibacterial ability, showed by their capability to produce clear zone against pathogenic bacteria by diffusion agar methods (Fig. 4). Their antibacterial activities were significantly different which affected by the strain of pathogenic bacteria.
Escherichia coli ATCC 25922, enteropathogenic E. coli (EPEC), S. typhimurium ATCC 14028 and S. aureus ATCC 25923 were able to be inhibited significantly (p<0.05) by LAB isolates (Fig. 4a-d). Inhibitory zones of LAB isolates against pathogenic bacteria were in the range of 5.5-14 mm which showed inhibitory activity of moderate (inhibition zones of 5-10 mm) to high for inhibition zones >10 mm (Savadogo et al., 2004). The cell free supernatant of LAB that used in this study had low pH value (3.7-4.1) and titratable acid were 0.28-0.57%. Based on the results, it was suggested that the antagonistic activity of these LAB strains relies on acidity, lactic acid or other organic acid produced. Other possible factors may be some bacteriocins which played roles at low pH value (Savadogo et al., 2004). If LAB survived, passed the stomach and colonized in the intestine, the acid environment they produced could inhibit the growth of harmful bacteria.
Antimicrobial activity of each LAB isolate affected differently to different species of pathogenic bacteria may be caused by antimicrobial components produced by each isolate were also different. Antimicrobial activity of LAB caused mainly by organic acids produced as a result of glucose metabolism. In this study, the total value of lactic acid in the 0.28-0.57 range with a pH range of 3.90-4.70%. Lactic acid is a weak acid having pKa value 3.86 at 25°C (Bogaert and Naidu, 2000).
The mechanism of inhibition of lactic acid to bacterial cell, was hydrophobic properties, thus facilitating diffusion in the form of protons into the cell through the cell membrane. As a result, intracellular pH is higher than the extracellular pH.
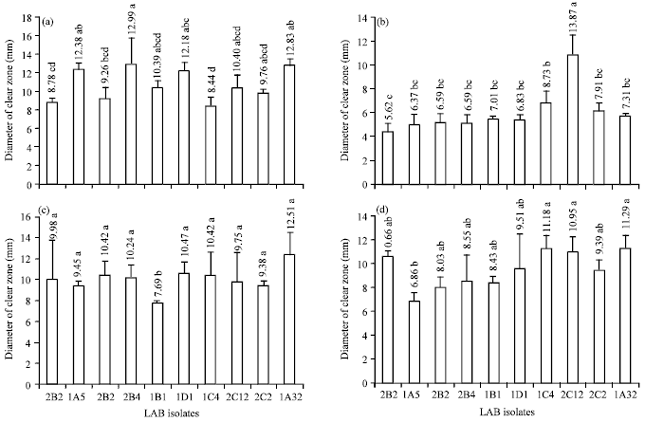 | |
| Fig. 4(a-d): | Diameter of clear zone of LAB isolates expressed antibacterial activity againts (a) E. coli ATCC 25922, (b) EPEC, (c) S. typhimurium ATCC 14028 and (d) S. aureus ATCC 25923 treatments with different letters are different at p<0.05 |
Furthermore, in the cell, lactic acid dissociates and lowers the intracellular pH with proton release (Bogaert and Naidu, 2000). The release of protons/hydrogen ions can disrupt metabolic functions such as the translocation of the substrate and oxidative phosphorylation, resulting in bacterial cell growth is inhibited.
Other antimicrobial compounds can be produced by LAB are bacteriocin. Bacteriocin is an extracellular protein or peptide molecules that has a bactericidal or bacteriostatic action against bacteria that have a close kinship (Savadogo et al., 2006; Albano et al., 2007). Bacteriocins can be degraded by protease enzymes in the digestive tract. Bacteriocins are irreversible, easy to digest, positive effect on health and active at low concentrations (Pal et al., 2005; Savadogo et al., 2006). Antimicrobial mechanisms of bacteriocin begins with entry into target cells by forming pores in the cell membrane that are sensitive and lower potential or pH gradient that causes damage to cellular material that could inhibit the growth of target cells (Ogunbanwo et al., 2003; Moll et al., 1999).
Coaggregation capacity: Probiotic and pathogenic bacteria can form joint aggregate called coaggregation (Surono, 2004). Probiotics are able to do coaggregation with pathogenics will effectively inhibit and kill pathogenic bacteria as antimicrobial compounds produced can move directly on pathogenics (Bao et al., 2010). Coaggregation capacity of LAB isolates against pathogenic bacteria are shown in Fig. 5a-d.
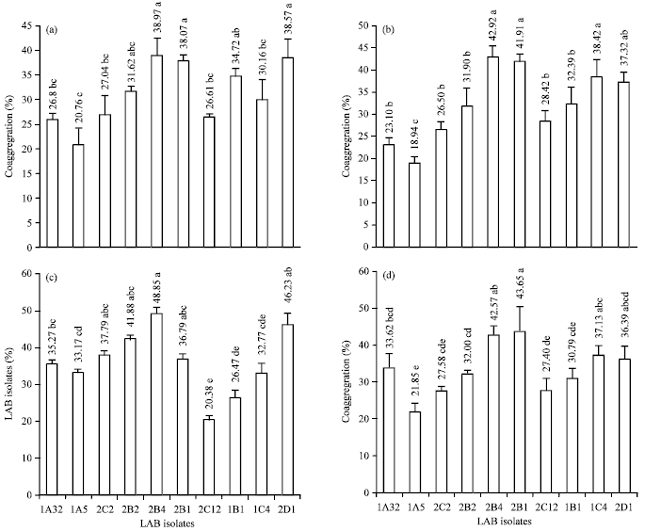 | |
| Fig. 5(a-d): | Coaggregation capacity of LAB isolates against pathogenic bacteria (a) E. coli ATCC 25922, (b) EPEC, (c) S. typhimurium ATCC 14028, (d) S. aureus ATCC 25923 treatments with different letters are different at p<0.05 |
LAB isolates had coaggregation capacity significantly to E. coli ATCC 25922 (p<0.05) of 20.76-38.97%. Coaggregation capacity of LAB with EPEC was statistically significantly different (p<0.05) with a range of value 18.94-49.92%. If LAB isolates formed coaggregate with pathogenic bacteria, the antimicrobial compounds produced by LAB will be more effective to work in inhibitory processes. Coaggregation capacity of LAB with S. typhimurium ATCC 14028 was significantly different (p<0.05) among the 10 isolates, at 21.85-43.65%. The LAB isolates were also able to perform a significantly different coaggregation capacity with S. aureus ATCC 25923 (p<0.05), at 20.38-48.85%. Comparing the strains of pathogenic bacteria, the highest coaggregation capacity of LAB isolates were against S. aureus ATCC 25923, followed by S. typhimurium ATCC 14028, EPEC and the lowest was E. coli ATCC 25922. Bao et al. (2010) reported that coaggregation L. fermentum against S. aureus was higher than that of E. coli and S. typhimurium. This is caused by the same cell wall morphology between LAB and S. aureus that is included in the Gram-positive which has a thick layer of peptidoglycan and hydrophobic making it easier to bond together. The results of this study, compares favorably with the results of Kos et al. (2003) reported that strains of L. acidophilus M92 had coaggregation capacity with E. coli at 15.11% and S. typhimurium by 15.70%. El-Naggar (2004) reported that L. acidophilus and L. plantarum possesses good coaggregation capacity with E. coli and S. typhimurium.
CONCLUSION
Identification by 16S rRNA gene sequencing of 20 isolates showed that 11 isolates were identified as Lactobacillus plantarum, 6 isolates as Lactobacillus acidophilus, 2 isolates as Pediococcus pentosaceus and 1 isolate as Enterococcus faecium. The twenty LAB isolates showed different ability to survive under simulated gastrointestinal conditions. Only 10 strains could survive well at pH 2.0 (survival rate over than 50%) and the population increased at pH 2.5, 3.2 and 7.2. The ten LAB strains (strain 2B4, 1B1, 1A5, 2C2, 2D1, 2B2, 1C4, 2B1, 1A32 and 2C12) could survive well at 0.5% bile salt, showed antibacterial activities and good coaggregation capacity against the tested pathogenic bacteria (Escherichia coli ATCC 25922, Salmonella typhimurium ATCC 14028, enteropathogenic Escherichia coli/EPEC and Staphylococcus aureus ATCC 25923). The ten LAB strains isolated from Indonesian local beef had characteristics as probiotic by in vitro analysis.
ACKNOWLEDGMENTS
This study was funded by Directorate General of Higher Education, Ministry of National Education Republic of Indonesia, through the 2011 Research Competitive Grant Program (I3.24.4/SPP/PHB/2011) and Sandwich-Like program at International Center for Biotechnology (IC-Biotech) Osaka University, Japan.
REFERENCES
- Albano, H., M. Oliveira, R. Aroso, N. Cubero, T. Hogg and P. Teixeira, 2007. Antilisterial activity of lactic acid bacteria isolated from Alheiras (traditional Portuguese fermented sausages): In situ assays. Meat Sci., 76: 796-800.
CrossRef - Bao, Y., Y. Zhang, Y. Zhang, Y. Liu and S. Wang et al., 2010. Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control, 21: 695-701.
CrossRefDirect Link - Begley, M., C. Hill and C.G.M. Gahan, 2006. Bile salt hydrolase activity in probiotics. Applied Environ. Microbiol., 72: 1729-1738.
CrossRefDirect Link - Brady, L.J., D.D. Gallaher and F.F. Busta, 2000. The role of probiotic cultures in the prevention of colon cancer. J. Nutr., 130: 410S-414S.
PubMedDirect Link - Cotter, P.D. and C. Hill, 2003. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev., 67: 429-453.
CrossRefDirect Link - El-Naggar, M.Y.M., 2004. Comparative study of probiotic cultures to control the growth of Escherichia coli O157: H7 and Salmonella typhimurium. Biotechnology, 3: 173-180.
Direct Link - Erkkila, S. and E. Petaja, 2000. Screening of commercial meat starter cultures at low pH and in the presence of bile salts for potential probiotic use. Meat Sci., 55: 297-300.
CrossRefDirect Link - Herich, R. and M. Levkut, 2002. Lactic acid bacteria, probiotics and immune system. Vet. Med. Czech, 47: 169-180.
Direct Link - Kimoto-Nira, H., K. Mizumachi, M. Nomura, M. Kobayashi and Y. Fujita et al., 2007. Lactococcus sp. as potential probiotic lactic acid bacteria. Jpn. Agric. Res. Q., 41: 181-189.
Direct Link - Kos, B., J. Suskovic, S. Vukovic, M. Simpraga, J. Frece and S.J. Matosic, 2003. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol., 94: 981-987.
CrossRefPubMedDirect Link - Lin, W.H., C.F. Hwang, L.W. Chen and H.Y. Tsen, 2006. Viable counts, characteristic evaluation for commercial lactic acid bacteria products. Food Microbiol., 23: 74-81.
CrossRefPubMedDirect Link - Marteau, P., M. Minekus, R. Havenaar and J.H.J. Huis In't Veld, 1997. Survival of lactic acid bacteria in a dynamic model of the stomach and small intestine: Validation and the effects of bile. J. Dairy Sci., 80: 1031-1037.
CrossRefDirect Link - Mishra, V. and D.N. Prasad, 2005. Application of in vitro methods for selection of Lactobacillus casei strains as potential probiotics. Int. J. Food Microbiol., 103: 109-115.
CrossRefDirect Link - Moll, G.N., W.N. Konings and A.J.M. Driessen, 1999. Bacteriocins: Mechanism of membrane insertion and pore formation. Antonie Van Leeuwenhoek, 76: 185-198.
PubMedDirect Link - Moser, S.A. and D.C. Savage, 2001. Bile salt hydrolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in lactobacilli. Applied Environ. Microbiol., 67: 3476-3480.
CrossRefDirect Link - Ogunbanwo, S.T., A.I. Sanni and A.A. Onilude, 2003. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr. J. Biotechnol., 2: 219-227.
Direct Link - Pal, V., M. Jamuna and K. Jeevaratnam, 2005. Isolation and characterization of bacteriocin producing lactic acid bacteria from a South Indian special Dosa (APPAM) batter. J. Cult. Collect., 4: 53-60.
Direct Link - Pan, X., F. Chen, T. Wu, H. Tang and Z. Zhao, 2009. The acid, bile tolerance and antimicrobial property of Lactobacillus acidophilus NIT. Food Control, 20: 598-602.
CrossRefDirect Link - Pennacchia, C., E.E. Vaughan and F. Villani, 2006. Potential probiotic Lactobacillus strains from fermented sausages: Further investigations on their probiotic properties. Meat Sci., 73: 90-101.
CrossRef - Savadogo, A., C.A.T. Ouattara, I.H.N. Bassole and S.A. Traore, 2006. Bacteriocins and lactic acid bacteria-a minireview. Afr. J. Biotechnol., 5: 678-683.
Direct Link - Savadogo, A., C.A.T. Ouattara, I.H.N. Bassole and A.S. Traore, 2004. Antimicrobial activities of lactic acid bacteria strains isolated from Burkina Faso fermented milk. Pak. J. Nutr., 3: 174-179.
CrossRefDirect Link - Zoumpopoulou, G., B. Foligne, K. Christodoulou, C. Grangette, B. Pot and E. Tsakalidou, 2008. Lactobacillus fermentum ACA-DC 179 displays probiotic potential in vitro and protects against trinitrobenzene sulfonic acid (TNBS)-induced colitis and Salmonella infection in murine models. Int. J. Food Microbiol., 121: 18-26.
CrossRefDirect Link - Weisburg, W.G., S.M. Barns, D.A. Pelletier and D.J. Lane, 1991. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol., 173: 697-703.
CrossRefPubMedDirect Link