
A.M. Abd El- Gawad
Department of Animal Production, Faculty of Agriculture, Cairo University, 12613, Egypt
Y.I. El- Talty
Department of Animal Production, Faculty of Agriculture, Cairo University, 12613, Egypt
K.M. Elsawy
Regional Center for Feed and Food, Agricultural Research Center, 12619, Egypt
A.E.M. Mahmoud
Department of Animal Production, Faculty of Agriculture, Cairo University, 12613, Egypt
M.A. Rawash
Regional Center for Feed and Food, Agricultural Research Center, 12619, Egypt
Asian Journal of Animal Sciences
Year: 2015 | Volume: 9 | Issue: 3 | Page No.: 119-127







ABSTRACT
This study aimed to evaluate the effects of feeding lactating buffaloes on different dietary protein sources on amino acids profile of milk and milk urea nitrogen. Eight lactating Egyptian buffaloes in second lactation season weighed 730±32 kg in average were used after 8 weeks after calving, the animals arranged in three swing over design. Tested sources protein were Soybean Meal (SBM), Cotton Seed Meal (CSM) and Sunflower Meal (SFM). Rations differed in source of protein supplement: Ration one (R1) contained three meals, ration two (R2) contained soybean meal and cottonseed meal; ration three (R3) contained soybean meal and sunflower meal and ration four (R4) contained soybean meal alone. Result indicated that SBM had higher essential, non-essential amino acids values. Also, total Non-Essential Amino Acids (NEAA) were numerically higher for SBM (22.29) vs. CSM (20.46) and SFM (14.78). The ratio EAA/NEAA value in SFM (0.91) was higher than soybean meal (0.89) and cotton seed meal (0.81). Ration three had the highest in most of essential amino acids, non-essential amino acids and total EAA (30.39). Total NEAA value was the highest in R3 (33.52) compared with other rations. Ration one recorded the highest values in most of essential, non-essential amino acids and total EAA and ratio of EAA/NEAA in milk compared with other experimental rations. Whereas, R4 had the lowest ratio of EAA/NEAA in milk. Milk urea nitrogen was significantly lower with R1 (11.67) compared with R4 which recorded highest value (12.97) followed by R2 (12.78) and R3 (11.90). It could be concluded that feeding lactating buffaloes on more than one source of protein in its rations had a positive effect on amino acids profile in milk and decrease milk urea nitrogen (MUN) level.
PDF Abstract XML References Citation
Received: February 16, 2015;
Accepted: April 21, 2015;
Published: May 06, 2015
How to cite this article
A.M. Abd El- Gawad, Y.I. El- Talty, K.M. Elsawy, A.E.M. Mahmoud and M.A. Rawash, 2015. Effect of Different Dietary Protein Sources on Amino Acids and Urea Nitrogen Contents of Dairy Buffaloes Milk. Asian Journal of Animal Sciences, 9: 119-127.
DOI: 10.3923/ajas.2015.119.127
URL: https://scialert.net/abstract/?doi=ajas.2015.119.127
DOI: 10.3923/ajas.2015.119.127
URL: https://scialert.net/abstract/?doi=ajas.2015.119.127
INTRODUCTION
Increasing pressure in the dairy industry to produce milk more efficiently while tailoring its composition to meet the demands of different market segments has given renewed impetus to devising appropriate feed formulations and feeding strategies to meet these challenges (Abu-Ghazaleh et al., 2001). Lactating animals require amino acids (AA) for milk production and these requirements vary with the level of milk production and composition. Required AA is supplied primarily by combinations of microbial protein and rumen degradable protein (RDP) (Faciola and Brodericket, 2014). The RDP portion of crud protein (CP) which consists of both true protein and non-protein nitrogen (NPN), is used to supply nitrogen for microbial protein production in the rumen, whereas RUP passes intact from the rumen. Both microbial protein and RUP contribute to the metabolizable protein pool (Davidson et al., 2003).
Optimizing the AA balance of rations has been proposed as one approach to enhance milk protein secretion and manipulate milk protein contents. Several strategies have been developed to optimize the AA profile of duodenal digesta to better meet the animal’s requirements for specific AA. These strategies include stimulation of microbial protein synthesis, feeding proteins resistant to ruminal degradation and feeding AA that have been treated to prevent ruminal degradation (Schwab et al., 1992). Milk urea nitrogen (MUN) is another tool to assess the protein and energy balance status of a group of dairy animals and can be used for minimizing feed costs while maximizing production. Milk urea nitrogen indicates the amount of urea found in milk and these values are closely correlated with the concentration found in the blood. Values of MUN can be used in conjunction with evaluating milk production records, feeding management practices, dry matter intake, degradable protein, un-degradable protein, nonstructural carbohydrates and water intakes. If MUN concentrations are outside of the recommended levels, it signifies that there is an imbalance between the rumen soluble carbohydrates and protein needed for microbial synthesis (Nousiainen et al., 2004; Eriksson and Rustas, 2014). Therefore, this study aimed to investigate the effect of different dietary protein sources (soybean meal, cotton seed meal and sunflower meal) on amino acids profile and milk urea nitrogen (MUN) of buffaloes milk.
MATERIALS AND METHODS
Experimental animals and rations: Eight lactating Egyptian buffaloes in second lactation seasons weighed 730±32 kg in average were used after 8 weeks after calving, the animal arranged in three swing over design according to El-Serafy (1968). So, the experiment started and ended with control which represented in ration one (R1) soybean meal (SBM), cottonseed meal (CSM) and sunflower meal (SFM). Ration 2 (SBM and CSM), R3 (SBM and SFM) and R4 (SBM) which represented different experimental sources of protein. The two controls are required to find the normal daily decrease in milk yield and daily increase in fat percentage. The experiment lasted for 140 days in five periods; each period consisted of 21 days for adaptation and 7 days for milk collection. The formulation and chemical composition of the experimental rations are presented in Table 1 and 2.
Feeding procedures: Animals were fed as group on the experimental rations to cover energy and protein requirements according to Paul et al. (2002). Rations offered as TMR to all animals, at 10 am and 6 pm with roughage concentrate ratio 60:40. Water was available at all times.
Milk sampling: During the 7 days collection period two milk samples were individually collected from each buffalo at 7.00 am and 7.00 pm then composite milk samples and stored at -20°C until analyses.
Amino acids analysis: Performic acid oxidation is performed prior to hydrolysis to oxidize cystine and methionine to cystine acid and methionine sulfone, respectively. Sodium metabisulfite is added to decompose performic acid. Amino acids are liberated from protein by hydrolysis with HCl.
| Table 1: | Formulation of the experimental rations (Total mixed ration) percentage on DM basis |
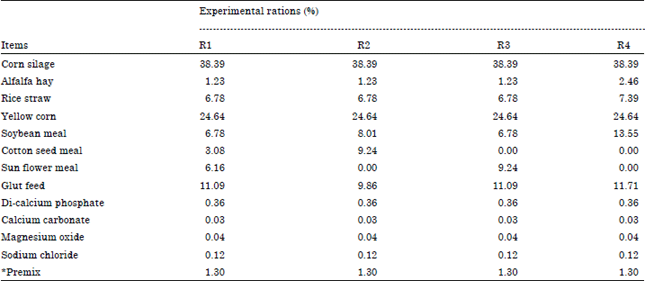 | |
| Premix: Antitoxins: 0.11, Yeast: 0.31, Minerals: 0.44, Vitamins: 0.44 and DM: Dry Matter | |
| Table 2: | Chemical composition and fiber fractions of the experimental feedstuffs and rations (percentage on DM basis) |
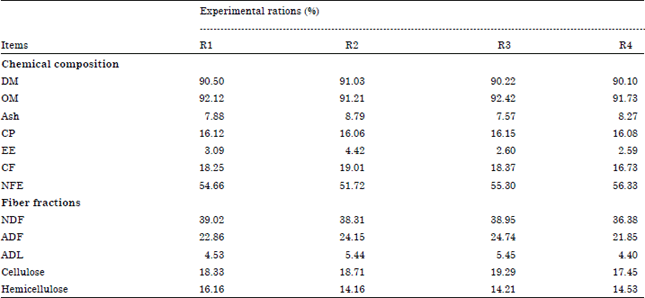 | |
| R1: CFM1+Roughage, R2: CFM2+Roughage, R3: CFM3+Roughage and CFM4+Roughage | |
Hydrolysates are diluted with sodium citrate buffer or neutralized, pH is adjusted to 2.20 and individual amino acid components are separated on ion-exchange chromatograph. Tryptophan is destroyed by hydrolysis, so, this amino acids cannot be determined (AOAC., 2012).
Milk urea nitrogen: Milk was conserved by 2-bromo-2-nitropropane-1, 3-diol and cooled to 4-6°C until analysed. At the end of each period were warmed to room temperature (21°C) and mixed thoroughly. Milk was deproteinized with 5 mL of milk from each milking was treated with 5 mL of 25% (w/v) Trichloroacetic acid (TCA). Samples were vortexed and allowed to stand for 30 min at room temperature (22-24°C) before filtering through Whatman No. 1 filter paper. Filtrates were stored at -20°C until MUN analysis by automated commercial Urea/Ammonia Assay Kit Spectrophotometer absorbance was 520 nm. The improved Jung method utilizes a chromogenic reagent that forms a colored complex specifically with urea. The intensity of the color, measured at 520 nm, is directly proportional to the urea concentration in the sample. The optimized formulation substantially reduces interference by substances in the raw samples (Broderick et al., 2013).
Statistical analysis: Data was analyzed using the general liner model procedure of SAS (2009). One way ANOVA procedure used to analyze data following the next model:
where, μ is the overall mean of Yij, Ri is the treatment effect, Eij is the experimental error. The differences among means were separated according to Duncan New Multiple Range Test (Duncan, 1955).
RESULTS AND DISCUSSION
Amino acids profile of the experimental meals: Result in Table 3 showed that soybean meal (SBM) was higher than cotton seed meal (CSM) and sunflower meal (SFM) in essential amino acids. Arginine (4.33) was higher in CSM than other two experimental meals (SBM and SFM), respectively. While, methionine (0.71) had higher in SFM than other experimental meals SBM (0.61) and CSM (0.55).
| Table 3: | Amino acids profile of the experimental meals |
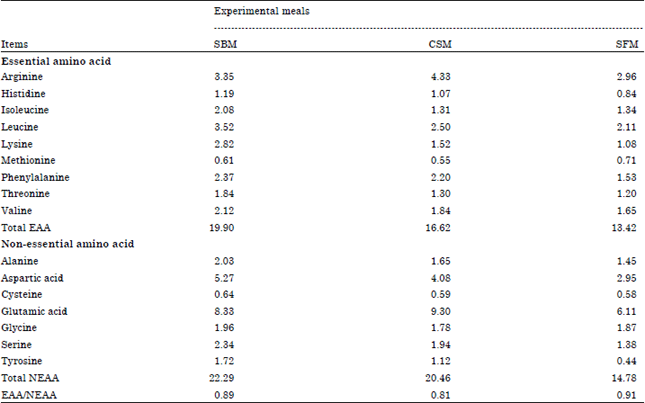 | |
| SBM: Soybean meal, CSM: Cotton seed meal and SFM: Sunflower meal | |
Data of the following amino acids (histidine, leucine, lysine, phenylalanine, threonine and valine) observed that CSM amino acids had higher values than SMF with the same amino acids. Isoleucine in CSM recorded the lowest value (1.31) from SFM (1.34) and SBM (2.08). Above results refers to total EAA in SBM was higher than other meals (CSM and SFM). Generally, SBM had higher values in non-essential amino acids. Glutamic acid which was high in CSM (9.3) then SBM (8.33) and SFM (6.11). Glycine was higher in SFM (1.87) than CSM (1.78). Sunflower meal had the lowest value in non-essential amino acids than others meals. Total NEAA value in SBM (22.29) was higher compared with CSM (20.46) and SFM (14.78). On the contrary, the ratio EAA/NEAA value in SFM (0.91) was higher than soybean meal (0.89) and cotton seed meal (0.81). Feedstuffs contain numerous different proteins and several types of NPN compounds. Proteins are large molecules that differ in size, shape, function, solubility and AA composition (NRC., 2001).
Amino acids profile of the experimental rations: Data in Table 4 illustrated that the R3 had the highest values in most of essential amino acids compared with other experimental rations. Whereas, Methionine (0.12) had the same value in (R1, R2 and R3) but R4 had the lowest in Methionine value (0.11). However, R1 had the lowest values in all essential amino acids compared to other experimental rations. On the other hand, Arginine in R4 (2.21) had the lowest value compared to R1 (2.32), R2 (2.64) and R3 (2.96), respectively. Not only, R3 was the highest in most of essential amino acids but also had the highest values in total EAA (30.39) then R2 (28.28), R4 (27.26) and R1 (23.14). Generally, R3 had higher values in non-essential amino acids.
| Table 4: | Amino acids profile of the experimental rations |
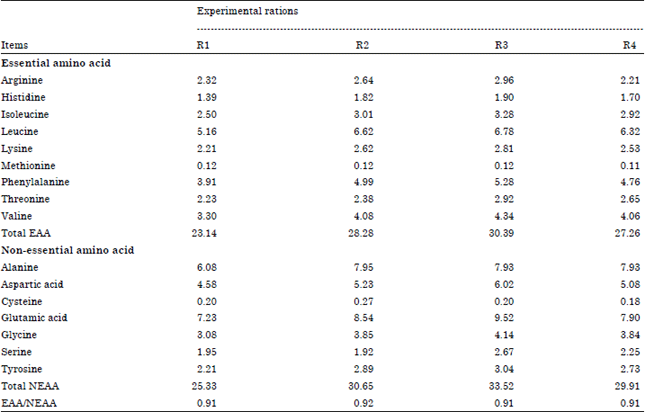 | |
| R1: CFM1+Roughage, R2: CFM2+Roughage, R3: CFM3+Roughage and R4: CFM4+Roughage | |
In the same time, Cysteine recorded the lowest value in R4 (0.18) compared with R1, R3 (0.20) and R2 (0.27). Results observed that R1 had lowest values in most of non-essential amino acids, excepted serine was higher value in R3 (2.67) followed by R4 (2.25), R1 (1.95) and R2 (1.92). Total NEAA value was the highest in R3 (33.52) compared with other rations. The ratio of EAA/NEAA value had the same value in R1, R3 and R4 (0.91), but had the highest value in R2 (0.92). Amino acid requirements may be supplied by formulating diets to maximize microbial protein synthesis while supplying additional RUP of the amount and quality that will complement microbial protein. Protein supplements are more expensive and increase the feed cost. By optimizing the use of protein supplement within the ruminant system, can reduce the quantity of protein in the diet or can enhance the production of the animals (Broderick, 2003). Any deficiency in quality of the AA profile provided solely by microbial protein can be corrected by feeding supplemental sources of RUP. These types of protein sources resist degradation by the ruminal microbes and reach the abomasum basically unaltered (Merchen and Titgemeyer, 1992).
Effect of the experimental rations on milk amino acids profile: Results in Table 5 showed that arginine in milk was significantly (p<0.05) higher for R1 (0.14) compared to other experimental rations R3, R4 (0.12) and R2 (0.09) respectively. Data of the following of essential amino acids (histidine, lysine, methionine and threonine) indicated that these were significantly (p<0.05) lower with R2 than other experimental rations whereas, there were no significant differences among R1, R3 and R4. Isoleucine had significantly higher for R4 (0.25) than R3 (0.21) and R2 (0.17).
| Table 5: | Effect of the experimental rations on milk amino acids profile |
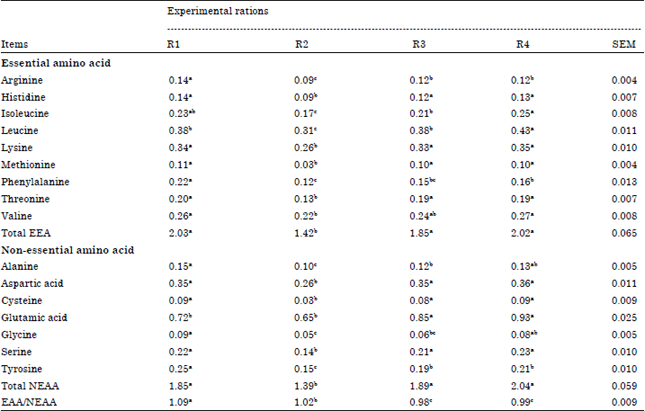 | |
| a,bMeans in the same row with different superscripts are significantly different (p<0.05). R1: CFM1+Roughage, R2: CFM2+Roughage, R3: CFM3+Roughage and R4: CFM4+Roughage | |
While, leucine had significantly (p<0.05) higher for R4 (0.43) followed by R1 (0.38) then R2 (0.31), whereas, R1 and R3 didn’t show any difference. On the other hand, R1 (0.22) was significantly (p<0.05) higher in phenylalanine followed by R4 (0.16) and R2 (0.12). Although, no significant difference between R2 (0.22) and R3 (0.24) in valine. But the same amino acid were significantly (p<0.05) lower for R2 followed by R4 (0.27) and R1 (0.26), respectively. However, R2 was significantly (p<0.05) lower in milk total EAA (1.42) compared with others. Result observed that each of following non-essential amino acids (aspartic, cysteine and serine) had significantly lower for R2 than other experimental rations. Ration two showed significant decrease in alanine (0.10) followed by R3 (0.12), R4 (0.13) and R1 (0.15). Glutamic acid in milk recorded higher value with R4 (0.93). Whereas, Glycine was significantly lower with R2 (0.05) than R4 and R1, respectively. Tyrosine was highly significant (p<0.05) in R1 (0.25) compared with R3 (0.19) and R2 (0.15). Total NEAA in R2 was significantly lower than other rations. The ratio EAA/NEAA in milk had highly significantly with R1 (1.09) compared with R2 (1.02), R3 (0.99) and R4 (0.98). Generally, data in Table 5 showed that R1 not only had the highest values in most of essential and non-essential amino acids but also had highest value in total EAA and ratio of EAA/NEAA compared with other experimental rations. In this pattern, Drackley and Schingoethe (1986) observed that animals fed on sunflower and soybean meal had a more desirable amino acid balance than animals fed soybean meal alone for high-producing animals. Due to the use of different sources of protein in quality, degradation (NRC., 1989) and profile of amino acids were different among feed stuffs and output (NRC., 2001). Factors effecting on utilization of dietary protein are complex and related to supplying sufficient rumen degradable protein to cover needs of rumen flora (Broderick, 2003).
Effect of the experimental rations on Milk Urea Nitrogen (MUN): Results in Fig. 1 indicated that milk urea nitrogen was significantly (p<0.05) higher with R4 (12.97) compared to R3 (11.90), R2 (12.78) and R1 (11.67). Milk urea nitrogen in R1 was the lowest value and in R4 was the highest value. This results refers to using one source of protein in ration had a negative effect compared to using two or three sources of dietary protein. This results matched with fact explained that dietary protein classified into two components, Rumen Degradable Protein (RDP) and Rumen Un-degradable Protein (RUP).
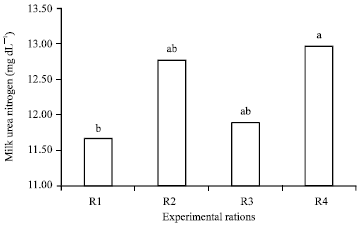 | |
| Fig. 1: | Effect of the experimental rations on milk urea nitrogen, a,bMeans in the same row with different superscripts are significantly different (p<0.05), R1: CFM1+Roughage, R2: CFM2+Roughage, R3: CFM3+Roughage and R4: CFM4+Roughage |
Rumen un-degradable protein is sometimes referred to as rumen by-pass protein because it resists breakdown in the rumen and broken down in the small intestine. Protein degraded in the rumen is used by the rumen microbes to make microbial protein which used by the animal to make milk (Nousiainen et al., 2004; Spek et al., 2013; Eriksson and Rustas, 2014).
CONCLUSION
Summing up, results indicated that using three different dietary protein sources with the same percentage of crud protein in the rations and different degradability improved the efficiency of obtained high ratio of amino acids which have high biological value in animal body and milk composition that effect on human health. The overall results indicated that feeding lactating buffaloes on different sources of dietary protein not only decrease the degradation rate of protein but also decrease Milk Urea Nitrogen (MUN) level.
REFERENCES
- Abu-Ghazaleh, A.A., D.J. Schingoethe and A.R. Hippen, 2001. Blood amino acids and milk composition from cows fed soybean meal, fish meal, or both. J. Dairy Sci., 84: 1174-1181.
CrossRefPubMedDirect Link - Broderick, G.A., 2003. Effects of varying dietary protein and energy levels on the production of lactating dairy cows. J. Dairy Sci., 86: 1370-1381.
CrossRefPubMedDirect Link - Broderick, G.A., T.M. Kerkman, H.M. Sullivan, M.K. Dowd and P.A. Funk, 2013. Effect of replacing soybean meal protein with protein from upland cottonseed, Pima cottonseed, or extruded Pima cottonseed on production of lactating dairy cows. J. Dairy Sci., 96: 2374-2386.
CrossRef - Davidson, S., B.A. Hopkins, D.E. Diaz, S.M. Bolt, C. Brownie, V. Fellner and L.W. Whitlow, 2003. Effects of amounts and degradability of dietary protein on lactation, nitrogen utilization and excretion in early lactation holstein cows. J. Dairy Sci., 86: 1681-1689.
CrossRefDirect Link - Drackley, J.K. and D.J. Schingoethe, 1986. Extruded blend of soybean meal and sunflower seeds for dairy cattle in early lactation. J. Dairy Sci., 69: 371-384.
CrossRef - Eriksson, T. and B.O. Rustas, 2014. Effects on milk urea concentration, urine output and drinking water intake from incremental doses of potassium bicarbonate fed to mid-lactation dairy cows. J. Dairy Sci., 97: 4471-4484.
CrossRef - Faciola, A.P. and G.A. Broderick, 2014. Effects of feeding lauric acid or coconut oil on ruminal protozoa numbers, fermentation pattern, digestion, omasal nutrient flow and milk production in dairy cows. J. Dairy Sci., 97: 5088-5100.
CrossRef - Merchen, N.R. and E.C. Titgemeyer, 1992. Manipulation of amino acid supply to the growing ruminant. J. Anim. Sci., 70: 3238-3247.
Direct Link - NRC, 2001. Nutrient Requirements of Dairy Cattle. 7th Edn., National Academy Press, Washington, DC, ISBN: 978-0-309-06997-7, Pages: 405.
CrossRefDirect Link - Nousiainen, J., K.J. Shingfield and P. Huhtanen, 2004. Evaluation of milk urea nitrogen as a diagnostic of protein feeding. J. Dairy Sci., 87: 386-398.
CrossRefPubMedDirect Link - Paul, S.S., A.B. Mandal and N.N. Pathak, 2002. Feeding standards for lactating riverine buffaloes in tropical conditions. J. Dairy Res., 69: 173-180.
CrossRefDirect Link - Schwab, C.G., C.K. Bozak, N.L. Whitehouse and M.M.A. Mesbah, 1992. Amino acid limitation and flow to duodenum at four stages of lactation. 1. Sequence of lysine and methionine limitation. J. Dairy Sci., 75: 3486-3502.
PubMed - Spek, J.W., A. Bannink, G. Gort, W.H. Hendriks and J. Dijkstra, 2013. Interaction between dietary content of protein and sodium chloride on milk urea concentration, urinary urea excretion, renal recycling of urea and urea transfer to the gastrointestinal tract in dairy cows. J. Dairy Sci., 96: 5734-5745.
CrossRef