
D. Paramanatha Swamidoss
School of Biological Sciences, Madurai Kamaraj University, Madurai-6250 21, Tamilnadu, India
P. Parvathiraj
Department of Zoology, Sri Paramakalyani College, Alwarkurichi-627412, Tamilnadu, India
M.R. Sudhakaran
Department of Zoology, Sri Paramakalyani College, Alwarkurichi-627412, Tamilnadu, India
Asian Journal of Animal Sciences
Year: 2014 | Volume: 8 | Issue: 1 | Page No.: 1-14







ABSTRACT
Postnatal growth in H. ater was analysed based on the longitudinal sampling of individuals of known age. Periodic visits were made to the bat roosting sites and the bats were monitored. Young ones of H. ater were observed to have a body mass of 1.5 gm at their birth. Male and female young ones attain 72% (3.8 gm) and 76% (3.6 gm) of the adult body mass in an age of 46 days and 48 days, respectively. Behaviour pattern in parturient bats was observed and the period of labor in H. ater occurs for about 25-32 min.
PDF Abstract XML References Citation
Received: March 18, 2013;
Accepted: June 01, 2013;
Published: February 27, 2014
How to cite this article
D. Paramanatha Swamidoss, P. Parvathiraj and M.R. Sudhakaran, 2014. Parturition and Postnatal Growth of Dusky Leaf-nosed Bat, Hipposideros ater. Asian Journal of Animal Sciences, 8: 1-14.
DOI: 10.3923/ajas.2014.1.14
URL: https://scialert.net/abstract/?doi=ajas.2014.1.14
DOI: 10.3923/ajas.2014.1.14
URL: https://scialert.net/abstract/?doi=ajas.2014.1.14
INTRODUCTION
Postnatal growth period, in bats is defined as the development of neonates to become weaned young ones (Kunz, 1987). The important features of postnatal development in bats are the attainment of flight and independence from parental care.
Postnatal growth studies are important for understanding how life-history traits are influenced by extrinsic and intrinsic factors (Kunz et al., 2009). In mammals, pre-weaning rate of growth may be influenced by the ability of the mother to provision the young. A large litter may decrease rates of growth by reducing the amount of food available to each individual. In cotton rats (Sigmodon hispidus), individuals from small litters grow faster than those from large litters (Rogowitz and McClure, 1995).
The growth rate patterns of bats have been evaluated using different growth models on very few species of bats such as Tadarida brasiliensis (Kunz and Robson, 1995), Pipistrellus mimus (Isaac and Marimuthu, 1996), Plecotus auritus (McLean and Speakman, 2000), Myotis nattereri (Swift, 2001), Hipposideros terasensis (Cheng and Lee, 2002), Myotis blythii (Sharifi, 2004), Hipposideros larvatus (Lin et al., 2010), Hipposideros Pomona (Jin et al., 2011). Postnatal growth period in bats varied from species to species. In mega bats like Pteropus giganteus it may extend to 120 days (Sudhakaran et al., 2013). Variation in morphometric growth can be used as an indicator in analyzing the postnatal growth, it is also used in studies like population biology and sexual dimorphism (Sterbing, 2002).
Bats are seasonal in the timing of their reproductive activities. Microchiroptera can be divided into two groups by reproductive pattern polyoestry and monoestry. Megachiroptera (African fruit bats: Pteropodidae) have more extended parturition periods than the microchiropterans and are more likely to show polyoestry. African bats are monoestrous and a high degree of synchrony in parturition time may exist both within and among species (Bernard and Cumming, 1997). The present study examines the parturition and postnatal development of leaf nosed bat, H. ater.
MATERIALS AND METHODS
Study animal: Hipposideros ater (dusky leaf-nosed bat) is a small species of family Hipposideridae with a forearm length and body mass ranging from 34.1-36.8 mm and 4.5-6 g, respectively. It consists of a horizontal horseshoe often with accessory folioles; an intermediate leaf and posterior leaf. There is a well defined frontal sac situated in the mid-line behind the posterior leaf of males. The pelage is variable in colour ranging from dull yellow, golden orange or pale gray to dark brown on the dorsal aspect. The ventral aspect is also variable in colour but is usually paler than the back. It is a widely distributed species, occurring from Sri Lanka and India to Malaysia, Indonesia, Philippines, New Guinea and north-western Australia (Bates and Harrison, 1997).
Field procedures: Postnatal growth studies in H. ater colony roosting in an abandoned building roost at Cheranmahadevi; Tamilnadu, India was conducted from April 2011 to June 2012. The observation of postnatal growth was based on the longitudinal (mark-recapture) sampling of individuals of known age (Burnett and Kunz, 1982). Day-old young ones were collected from the natural roosts, sexed, marked and the morphometric measurements were recorded. For marking, the non-toxic paints of two different colours (red colour for male pup and white colour for female pup) were applied on the claw of first toe and thumb. The major criterion for assessing day-old status of young was based on the presence of fresh umbilical cord (Kunz, 1973). Morphometric growth parameters such as length of forearm, fingers, tail, tibia and condylobasal were measured to the nearest 0.1 mm using dial caliper. Body mass was recorded to the nearest 0.1 g using 50 g (Avinet) spring balance. At an interval of approximately 10 days, young bats that had been previously marked were reexamined, once again marked in the claw of second toe and the morphometric characters were measured. In addition, behaviour of the pups was also monitored. This process continued till the separation of young from adult.
Attempts were made to observe the parturition behaviour under natural conditions. Periodic visits were made at least twice a week, by day and night and more frequently during the parturition period. The behaviour of parturient females, labour and delivery were observed in close quarters. To minimize disturbances, parturient females were observed by using red-filtered torchlight (>610 nm).
Statistical analysis: Postnatal growth curves of both species were constructed based on linear growth of forearm and body mass compared with relative age in days. The linear increase in length of forearm, body mass, length of III finger and length of V finger from neonate to volant and from volant to weaned were used to derive regression equation. A ninety-five percent confidence interval was used for regression equations.
RESULTS
Parturition: H. ater was observed to be a monotocous species and have a seasonal bimodal breeding pattern with definite breeding periods during the month of mid April and last week of September. It was possible to observe the complete process of parturition in 3 parturient females of H. ater in close quarters. 2 bats took 25 and 32 min to complete the process of labour during delivery. The sequence of behaviour exhibited during delivery/labour of the third bat started at 11.30 h with dilation of the vaginal opening and ended at 11.57 h with the mother keeping the young one in one of her pubic teats. Pregnant bats were found restless throughout the process of parturition. After delivery, the mother bat immediately licked its pup. In all the cases, parturient females chewed the placenta. The complete process of parturition behaviour is tabulated in Table 1.
| Table 1: | Labour sequence during parturition of H. ater |
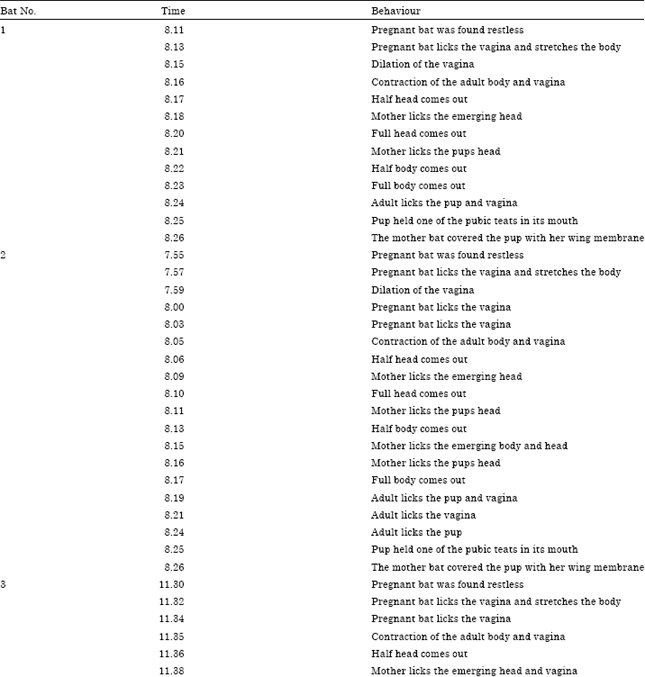 | |
 | |
Postnatal growth and development: During our study period a total of 64 (30 male and 34 female) neonates were marked for growth analysis. Out of which, a minimum of 4 bats in each sex and a maximum of 5 bats were recaptured on subsequent weeks till the date of weaning. At birth, the young ones of both the sexes were altrical. They were naked and pink in colour, with eyes closed and folded ear pinnae. The wing membranes were translucent and umbilical cord was found attached. The mean length of umbilical cord in male and female pup was 4.00± 0.75 and 4.21±0.49 mm, respectively. The morphometric parameters of neonates of both sexes were given in Table 2. In male neonates, the mean forearm length was 15.88±0.46 mm (n = 10) and the mean body mass was 1.55±0.16 (n = 10). In female neonates the mean forearm length was 15.95±0.37 mm and the mean body mass was 1.55±0.16 g (n = 10).
Male and female neonates that were attached to the pubic teats of mother bats became volant at an age of 28 days. However, neonate visited their mothers periodically and suckled upto the period of weaning. The morphometric growth parameters of volant pups of both sexes were given in Table 2. The average forearm length of male volant pups was 30.15±1.19 mm (n = 4) and its mean body mass was 2.75±0.29 g (n = 4). In the case of female volant pups, mean forearm length and body mass were 32.33±0.49 mm (n = 4) and 2.85±0.29 g (n = 4), respectively. During volant stage, male pups reached an average body mass of 55.0% and a forearm length of 83.3% of the adult bats. Volant female pups reached an average body mass of 57.0% and forearm length of 89.31% of the postpartum adult female bats.
The growth parameters were found to increase in a linear manner in both the sexes. In male volant bats, regression analysis made from neonate to volant stage between the growth of the body weight and the forearm length (Y = 0.0859X+0.2912; R2 = 0.7852) and between the growth of III finger and forearm length (Y = 1.8295X-10.679; R2 = 0.9176) and between the growth of V finger and forearm length (Y = 1.6438X-8.4731; R2 = 0.9397) shows a positive linear relation and the correlation coefficient is nearer to 1 (Fig. 1: Graph 1.1).
In female volant bats, the regression analysis made from neonate to volant stage between the growth of the body weight and the forearm length (Y = 0.078 X+0.3354; R2 = 0.8339) and between the growth of III finger and forearm length (Y = 1.3742X-3.336; R2 = 0.9751) and between the growth of V finger and forearm length (Y = 1.1911X-1.0109; R2 = 0.9664) shows a positive linear relation and the correlation coefficient is nearer to 1 (Fig. 2: Graph 2.1). When the pups of both sexes reached the volant stage, they began to flap their wings while hanging from their mothers’ neck. Flapping of wings by volant’s was observed frequently in day roost. The rate of flapping increased in a linear manner upto the stage of weaning. Rate of wing flapping of young ones was found to be higher during the onset of foraging of adult bats and in the morning hours.
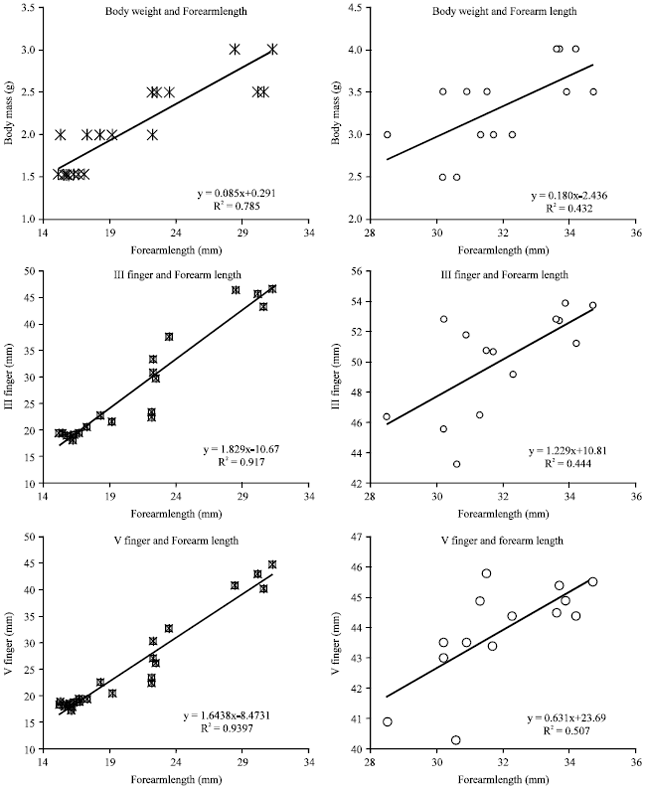 | |
| Fig. 1: | Regression of Growth Parameters of Male H. ater |
The male pups of H. ater weaned at an age of around 46 days and the female pups weaned at an age of 48 days.
The male pups of H. ater did not begin to forage till they reached 72% of the adult post-partum body weight and the female pups did not begin to forage till they reached 76% of the adult post-partum body weight (5.0 g). The average forearm length of weaned male bats was 34.02±0.44 mm (n = 5) and mean body mass was 3.60±0.22 g (n = 5).
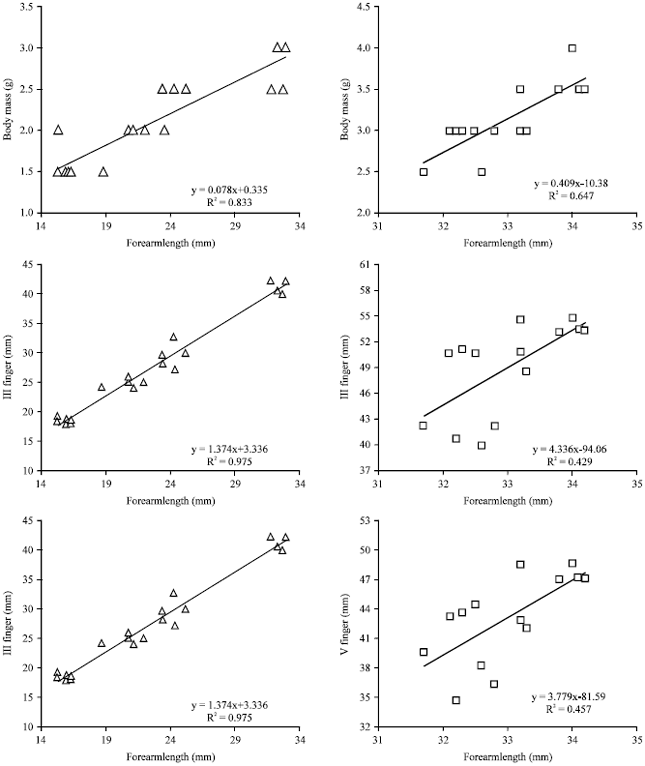 | |
| Fig. 2: | Regression of Growth Parameters of Female H. ater |
In the case of female weaned bats, mean forearm length and body mass was 33.86±0.39 mm (n = 5) and 3.80±0.22 g (n = 5), respectively (Table 2).
There is a linear increase in the morphometric growth parameters of young ones from volant stage to weaned stage.
| Table 2: | Morphological parameters of male and female pups of H. ater |
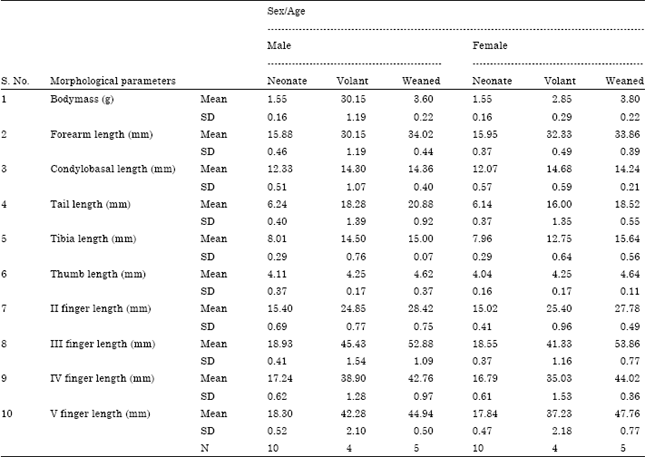 | |
In male weaned bats, regression analysis made from volant to weaned stage, between the growth of the body weight and the forearm length (Y = 0.1802X-2.4365; R2 = 0.4323), between the growth of III finger and forearm length (Y = 1.2296X +10.815; R2 = 0.444) and between the growth of V finger and forearm length (Y = 0.6319X + 23.695; R2 = 0.5073) shows a positive linear relation and the correlation coefficient is nearer to 1 (Fig. 1: Graph 1.2).
In female weaned bats, regression analysis made from volant to weaned stage, between the growth of the body weight and the forearm length (Y =1.4098X-10.382; R2 = 0.6478, between the growth of III finger and forearm length (Y = 4.3361X-94.06; R2 = 0.4299) and between the growth of V finger and forearm length (Y = 3.7799X-81.593; R2 = 0.4573) shows a positive linear relation and the correlation coefficient is nearer to 1 (Figure 2: Graph 2.2). The study on growth patterns of the forearm length and body mass of male and female pups shows that there is an increasing rate of growth in linear dimensions. The rate of growth of female pups was higher than that of male pups (Table 3). The growth pattern of forearm length and body mass of male and female pups of H. ater shows that there is a statistically insignificant difference during various growth periods.
DISCUSSION
During the early stages of labour, pregnant bats of H. ater remain in their natural freely hang head-down posture and hangs with legs hooked wide apart. A similar pattern of such behaviour was observed in Cynopterus sphinx (Ramakrishna, 1950), Megaderma lyra (Gopalakrishna et al., 1975; Balasingh, 1990).
| Table 3: | Comparison of morphological variables between male and female pups of H. ater |
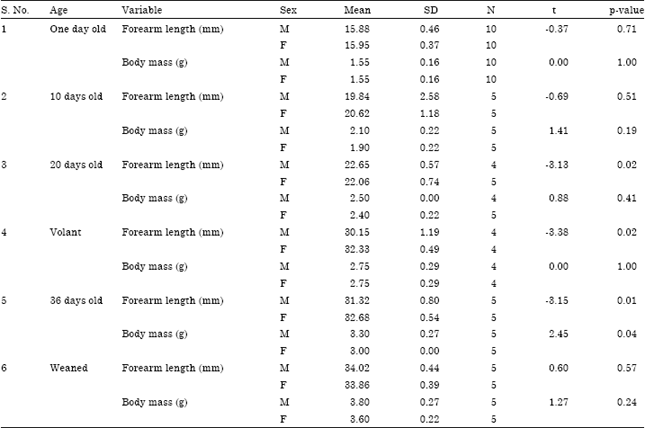 | |
In most bat species so far studied, parturition is a seasonal phenomenon that the young are born at a particular time of a year, which is presumably most favourable for their nurture and survival. In temperate bats, the time of parturition invariably falls in the spring or early summer. In many tropical forms, which live in relatively constant environment, the time of parturition falls during the period of vernal equinox (Baker and Baker, 1936) whereas in some polyestrous species like Desmodus rotundus (Mills, 1980) and Pipistrellus mimus (Gopalakrishna et al., 1975) continuous breeding is noticed. H. ater shows a seasonal bimodal breeding pattern, like most tropical bats like C. sphinx (Ramakrishna, 1950) and Rousettus leschenaulti (Gopalakrishna and Choudhari, 1977) and also like that of some temperate species Artibeus literatus (Wilson, 1979) and Epomops buettikoferi (Thomas and Marshall, 1984). Wimsatt (1979) reported that the spread in parturition dates might be expected to be greater in tropical forms, especially those that are polyestrous.
Parturition has been observed to occur in the day roost, which feature resembles that of most of the other species of bats like C. sphinx (Ramakrishna, 1950), M. lyra (Gopalakrishna et al., 1976; Balasingh, 1990) and Rhinophoma kinneari (Anand Kumar, 1965); hence, it can be assumed that parturition in general occurs some time during the periods of rest (Wimsatt, 1979).
Literature on the breeding behaviour of bats reveals that polytocous species, which bring forth more than one foetus (usually two foetus e.g. Myotis austroriparius) have longer duration of labour, while comparing with the monotocous species with exceptions. A similar pattern was observed in H. ater (25-32 min). In Phyllostomidae, Brown flower bats, Erophylla sezekorni the complete process of parturition occurs in between ca. 30-35 min (Soto Centeno and Kurta, 2003). Vespertilionids seem to have the shortest duration as reported by Sherman (1937) in Tadarida brasiliensis cynocephala, where the whole process of parturition completed in one and a half min. Like most other bat species, in H. ater, the movement of foetus involved a shift from a transverse position to longitudinal position.
Breech presentation of the foetus is common among several species of bats and most of the vespertilionids (Sherman, 1937), whereas in some species of bats like C. sphinx (Ramakrishna, 1950) of Pteropodidae and in E. sezekorni of Phyllostomidae (Soto Centeno and Kurta, 2003) and H. ater, the foetus is delivered headfirst.
The sequence of behaviour during delivery involving the movement of foetus, participation of foetus and mother in the presentation of the foetus, delivery of the placenta and placentophagy closely resembles that of M. lyra (Balasingh, 1990). The biting and pulling of the placenta by the mother may be interpreted as the deliberate effort to dislodge the placenta to hasten its delivery. The consumption of placenta by the mother suggests that placentophagy is relatively constant in this species.
At birth, the neonates of both sexes were altrical, naked and pink in colour with closed eyes and folded ear pinnae. The wings were translucent and the umbilical cord was found attached. The morphometric measurements recorded in day-old male and female pups did not differ significantly. In R. leschenaulti and C. sphinx (Elangovan et al., 2002, 2003) day-old pups had the same characters as said above and the above findings were also observed in a few microchiropterans like M. lyra (Subbaraj et al., 1997), Myotis nattereri (Swift, 2001), E. sezekorni (Soto Centeno and Kurta, 2003), P. pipistrellus (Hughes et al., 1995) and in a few vespertilionid bats (Gould, 1971, 1975). But in the case of some phyllostomids (Kleiman and Davis, 1979) which bear all the characters as observed above have got fur around the body at birth.
In the present study the average length of forearm and the average body mass of the male and the female pups of H. ater at birth were 15.88±0.46 mm; 1.55±0.16 g (n = 10) and 15.95±0.37 mm; 1.55±0.16 g (n = 10), respectively. In Rhinolophus ferrumequinum (a large member of Rhinolophidae in the temperate region, with an adult body mass of 23.6 g), the body mass at birth is 5.8 g (Kunz and Hood, 2000). In P. pipistrellus at birth it is 1.19±0.17 g (Hughes et al., 1995). The young of most microchiropterans weigh around 20 to 30% and mostly about 25% of their mother weight at birth (Altringham, 1996). From the perspective of relative body mass at birth, bats are clearly precocial because they are generally heavier than the young of other mammals having comparable litter size (Hayssen and Kunz, 1996; Kurta and Kunz, 1987), which average only 5-10% of the adult body weight (Altringham, 1996). However, bats are highly altrical with respect to development of forelimb and locomotive function (Powers et al., 1991).
Altringham (1996) reported that megachiropterans could fly at an age of 9-12 weeks, with weaning at 15-20 weeks, whereas in microchiropterans these traits occur at 2-6 and 5-10 weeks, respectively. Young vespertilionid bats become volant at an age between 2 weeks and 2 months, mostly at an age of 3-4 weeks and weaning occurs between 5 and 8 weeks (Tuttle and Stevenson, 1982). Young H. ater born in mid-April and in the last week of September attained the volant stage at an age of 28 days. The pattern of postnatal growth and development showed a basic trend of linear growth of forearm and body mass during volant period. Juveniles of several species of microchiropteran bats typically began to fly when they attained 70% adult body mass (Barclay, 1994). However in H. ater the male and female pups attained volant stage when they reached 55.0% and 57.05% of the adult body mass. (Elangovan et al., 2002, 2003) observed that the young R. leschenaulti and C. sphinx were found to attain volant stage when they reached 35.0% and 40.0% of adult body mass, respectively. It also substantiates the statement of Orr (1970), which stated that microchiropterans resemble the megachiropterans in the course of their development. Among vespertilionids, the growth rate of forearm ranges between 0.4 and 1.6 mm day-1 and body weight between 0.1 and 0.47 g day-1 (Tuttle and Stevenson, 1982). The growth rate of forearm, body mass and finger length of H. ater is relatively linear upto the volant stage and it is observed in both male and female bats. Kunz (1973) observed that Myotis velifer spread its wings and forced them in a downward motion against the roost substrate one week before the onset of flight, young bats usually engaged in so-called push-ups. A similar pre-flight activity was observed in Rhinolophus cornutus (Yokoyama et al., 1975), E. serotinus, Nyctalus noctula, P. pipistrellus (Kleiman, 1969). H. ater also exhibit a similar behaviour.
Male and female pups of H. ater, weaned at an average age of 46 and 48 days when they attained 72% and 76% of the adult body mass, respectively. At this stage, the pups of both sexes attained true flight and forage independently. Other species like Hipposideros commersoni weaned at an age of 20 weeks (Brosset, 1969) and H. terasensis weaned at an age of 2.5 months (Cheng and Lee, 2002). As observed in other microchiropterans like M. lyra (Rajan and Marimuthu, 1999), Myotis lucifugus (Kunz and Anthony, 1982), P. mimus (Isaac and Marimuthu, 1996), H. ater showed a linear growth in their morphometric measurements. The time at which the pups of H. ater attain volant stage does not seem to be much different from that of other tropical hipposiderid and vespertilionid species but their weaning time is earlier than that of some tropical species of hipposiderid. The phenomenon of early weaning time in the pups of H. ater is similar to that of some temperate species, such as Lasiurus cinereus and R. ferrumequinum nippon. Young of this species become weaned when they are 4 to 7 weeks old (Koehler and Barclay, 2000; Sano, 2000). One of the reasons for H. ater having an earlier weaning time may be related to the survival strategy.
Sexual dimorphism with regard to body mass and the length of forearm in various species commonly occur before weaning, suggesting differential maternal investment (Stern and Kunz, 1998; Holroyd, 1993). In the case of Phyllostomus hastatus, a pattern of faster mass gain in males during suckling period has been noted (Stern and Kunz, 1998) and also in other sexually dimorphic species (Lee and Moss, 1986; Soderquist, 1995). But in the case of H. ater, a faster mass gain was noted in females than in males during suckling period. A similar pattern of faster mass gain was also noted in Antrozous pallidus (Davis, 1969) and E. fuscus (Kunz, 1974). The fact that females weighed slightly more than males after this period may be due to different rates of fat deposition.
Kunz and Hood (2000) reviewed and compared the postnatal growth rate of body mass of 41 species of bats including 31 microchiropterans from temperate and tropical regions, by means of the logistic growth equation. They found a significant negative correlation between postnatal growth rate and asymptotic body mass. The postnatal growth of the H. ater (in the case of H. ater body mass at birth is 1.5 g, length of forearm averages 15.8 mm, which increases rapidly to reach 94.4% of adult size in just 46-48 days) is faster than that of the other tropical microchiropteran bats such as M. lyra-body mass at birth is 8.6 g, length of forearm averages 30.6 mm, which increases rapidly to reach 98.5% of adult size in 73 days (Subbaraj et al., 1997) but slower than that of temperate bats such as R. ferrumequinum-body mass at birth is 5.8 g, length of forearm of neonates averages 25.2 mm, which increases rapidly to reach 90.4% of adult size in just 16-18 days (Kunz and Hood, 2000).
CONCLUSION
The study on parturition and postnatal development in H. ater revealed that, it had a bimodal breeding pattern. Parturition occurs during the month of mid April and last week of September. Postnatal study revealed that the young ones become weaned at an age of ca.46 days. Raining season and food availability plays a vital role in determining the breeding season in bats. Young bats exhibit a linear growth in morphometric growth parameters from their neonate to weaned stage.
ACKNOWLEDGMENT
We followed the Guidelines for the use of animals in research (ABS, 1991). This study was supported by Department of Science and Technology ‘Fast Track Scheme for Young Scientists to DPSD (File No. SR/FT/LS-73/2010) and a UGC Major Research Project to MRS (File No. 39-665/ 2010 SR).
REFERENCES
- Lin, A.Q., L.R. Jin, Y. Liu, K.P. Sun and J. Feng, 2010. Postnatal growth and age estimation in Horsfield’s leaf-nosed bat Hipposideros larvatus. Zool. Stud., 49: 789-796.
Direct Link - Anand Kumar, T.C., 1965. Reproduction in the rat-tailed bat Rhinopoma kinneari. Proc. Zool. Soc. London, 147: 147-155.
CrossRef - Soto Centeno, J.A. and A. Kurta, 2003. Description of fetal and newborn brown flower bats, Erophylla sezekorni (Phyllostomidae). Caribbean J. Sci., 39: 233-234.
Direct Link - Barclay, R.M.R., 1994. Constraints in reproduction by flying vertebrates: Energy and calcium. Am. Nat., 144: 1021-1031.
Direct Link - Bernard, R.T.F. and G.S. Cumming, 1997. African bats: Evolution of reproductive patterns and delays. Q. Rev. Biol., 72: 253-274.
PubMedDirect Link - Burnett, C.D. and T.H. Kunz, 1982. Growth rates and age estimation in Eptesicus fuscus and comparison with Myotis lucifugus. J. Mammal., 63: 33-41.
Direct Link - Cheng, H.C. and L.L. Lee, 2002. Postnatal growth age estimation and sexual maturity in the formosan leaf-nosed bat (Hipposideros terasensis). J. Mammal., 83: 785-793.
Direct Link - Davis, R., 1969. Growth and development of young pallid bats, Antrozous pallidus. J. Mammal., 50: 729-736.
Direct Link - Elangovan, V., H. Raghuram, E. Yuvana Satya Priya and G. Marimuthu, 2002. Postnatal growth, age estimation and development of foraging behaviour in the fulvous fruit bat Rousettus leschenaulti. J. Biosci., 27: 695-702.
PubMedDirect Link - Elangovan, V., E. Yuvana Satya Priya, H. Raghuram and G. Marimuthu, 2003. Postnatal development in the Indian short-nosed fruit bat Cynopterus sphinx: Growth rate and age estimation. Acta Chiropterol., 5: 107-116.
Direct Link - Gould, E., 1975. Neonatal vocalizations in bats of eight genera. J. Mammal., 56: 15-29.
PubMedDirect Link - Hayssen, V. and T.H. Kunz, 1996. Allometry of litter mass in bats: Maternal size, wing morphology and phylogeny. J. Mammal., 77: 476-490.
Direct Link - Hughes, P., J.M.V. Rayner and G. Jones, 1995. Ontogeny of true flight and other aspects of growth in the bat Pipistrellus pipistrellus. J. Zool., 236: 291-318.
CrossRef - Isaac, S. and G. Marimuthu, 1996. Postnatal growth and age estimation in the Indian pygmy bat Pipistrellus mimus. J. Mammal., 77: 199-204.
Direct Link - Koehler, C.E. and R.M.R. Barclay, 2000. Post-natal growth and breeding biology of the hoary bat (Lasiurus cinereus). J. Mammal., 81: 234-244.
Direct Link - Kleiman, D.G., 1969. Maternal care, growth rate and development in the noctule (Nyctalus noctula), pipistrelle (Pipistrellus pipistrellus) and serotine (Eptesicus serotinus) bats. J. Zool., 157: 187-211.
CrossRef - Kunz, T.H., 1974. Reproduction, growth and mortality of the vespertilionid bat, Eptesicus fuscus, in Kansas. J. Mammal., 55: 1-13.
PubMedDirect Link - Kunz, T.H. and E.L.P. Anthony, 1982. Age estimation and postnatal growth in the bat Myotis lucifugus. J. Mammal., 63: 23-32.
Direct Link - Kurta, A. and T.H. Kunz, 1987. Size of bats at birth and maternal investment during pregnancy. Symp. Zool. Soc. Lond., 57: 79-106.
Direct Link - Kunz, T.H. and S.K. Robson, 1995. Postnatal growth and development in the Mexican free-tailed bat (Tadarida brasiliensis mexicana): Birth size, growth rates and age estimation. J. Mammal., 76: 769-783.
Direct Link - Lee, P.C. and C.J. Moss, 1986. Early maternal investment in male and female African elephant calves. Behav. Ecol. Sociobiol., 18: 353-361.
CrossRefDirect Link - Powers, L.V., S.C. Kandarian and T.H. Kunz, 1991. Ontogeny of flight in the little brown bat, Myotis lucifugus: Behavior, morphology and muscle histochemistry. J. Comput. Phys., 168: 675-685.
CrossRef - Rajan, K.E. and G. Marimuthu, 1999. Postnatal growth and age estimation in the Indian false vampire bat (Megaderma lyra). J. Zool., 248: 529-534.
CrossRef - Rogowitz, G.L. and P.A. McClure, 1995. Energy export and offspring growth during lactation in cotton rats (Sigmodon hispidus). Funct. Ecol., 9: 143-150.
Direct Link - Sharifi, M., 2004. Postnatal growth in Myotis blythii (Chiroptera, Vespertilionidae). Mammalia, 68: 283-290.
Direct Link - Sterbing, S.J., 2002. Postnatal development of vocalizations and hearing in the phyllostomid bat, Carollia perspicillata. J. Mammal., 83: 516-525.
CrossRefDirect Link - Stern, A.A. and T.H. Kunz, 1998. Intraspecific variation in postnatal growth in the greater spear-nosed bat. J. Mammal., 79: 755-763.
Direct Link - Sudhakaran, M.R., D. Paramanatha swamidoss and P. Parvathiraj, 2013. Postnatal growth in pteropus giganteus. Asian J. Exp. Biol. Sci., 4: 123-128.
Direct Link - Swift, S.M., 2001. Growth rate and development in infant Natterer's bats (Myotis nattereri) reared in a flight room. Acta Chiropterol., 3: 217-223.
Direct Link - Thomas, D.W. and A.G. Marshall, 1984. Reproduction and growth in three species of West African fruit bats. J. Zool., 202: 265-281.
CrossRef - Wimsatt, W.A., 1979. Reproductive asymmetry and unilateral pregnancy in Chiroptera. J. Reprod. Fertil., 56: 345-357.
CrossRefDirect Link - Yokoyama, K., T. Uchida and S. Shiraishi, 1975. Functional morphology of wings from the standpoint of adaptation for flight in Chiroptera, 1: Relative growth and ossification in forelimb, wing loading and aspect ratio. Zool. Mag., 84: 233-247.
Direct Link