
A. Sudha
Department of Bioinformatics, Alagappa University, Karaikudi -630004, Tamil Nadu, India
K. Sumathi
Department of Biochemistry, STET Women`s College, Mannargudi-614001, Tamil Nadu, India
S. Manikandaselvi
Department of Biochemistry, STET Women`s College, Mannargudi-614001, Tamil Nadu, India
N.M. Prabhu
Department of Animal Health and Management, Alagappa University, Karaikudi -630004, Tamil Nadu, India
P. Srinivasan
Department of Bioinformatics, Alagappa University, Karaikudi -630004, Tamil Nadu, India
Asian Journal of Animal Sciences
Year: 2013 | Volume: 7 | Issue: 1 | Page No.: 1-13







ABSTRACT
The present study was undertaken to evaluate the hepato-protective effect of crude flavonoid fraction of aerial parts of Lippia nodiflora in ethanol induced oxidative stress in liver using experimental animal models. Ethanol-fed (5 g kg-1 day-1) male rats were treated by crude flavonoid fraction (25, 50 mg kg-1) for 21 days. The liver damage was indicated by the significant increase in serum AST, ALT, ALP, LDH, total bilirubin, urea and decrease in total protein and triglyceride (TG). Lipid peroxidation markers like Thiobarbituric Acid Reactive Substances (TBARS) and antioxidant enzymes namely superoxide dismutase (SOD), catalase (CAT), reduced glutathione (GSH), Glutathione Reductase (GR) and Glutathione-S-transferase (GST) activities were also measured. The crude flavonoid fraction showed significant (p<0.05) protective effect by decreasing the elevated liver marker enzymes, total bilirubin, lipid peroxidation marker and ameliorated the diminished serum total protein as well as antioxidant levels in a dose dependent manner. The protective effect of the crude flavonoid fraction was observed at both concentrations and was compared to that of the standard used. Hence, our results highlights the ability of crude flavonoid fraction of Lippia nodiflora to ameliorate oxidative damage in the liver and the observed effects are associated with its antioxidant activities.
PDF Abstract XML References Citation
Received: October 15, 2012;
Accepted: December 12, 2012;
Published: May 03, 2013
How to cite this article
A. Sudha, K. Sumathi, S. Manikandaselvi, N.M. Prabhu and P. Srinivasan, 2013. Anti-hepatotoxic Activity of Crude Flavonoid Fraction of Lippia nodiflora
L. on Ethanol Induced Liver Injury in Rats. Asian Journal of Animal Sciences, 7: 1-13.
DOI: 10.3923/ajas.2013.1.13
URL: https://scialert.net/abstract/?doi=ajas.2013.1.13
DOI: 10.3923/ajas.2013.1.13
URL: https://scialert.net/abstract/?doi=ajas.2013.1.13
INTRODUCTION
Liver is the heaviest organ in the body and plays a crucial role in protecting various biological functions and helps in detoxification and excretion of many endogenous and exogenous compounds including chronic alcoholism (Deshmukh et al., 2008). The mechanism of detoxification results in the progress of many reactive intermediates like alcohol-induced oxidative stress and lipid peroxidation in the liver (Zhou et al., 2002). Acute ethanol administration causes liver damage because 80% of ingested alcohol is metabolized in the liver. The accountability of oxidative stress is believed to be the major concern in alcoholic liver damage (Caro and Cederbaum, 2004). The liver enzyme, alcohol dehydrogenase catalyzes the conversion of ethanol into cytotoxic acetaldehyde which in turn gets oxidized into acetate by aldehyde oxidase or xanthine oxidase by means of cytochrome P450 2E1 (CYP 2E1) and leads to Reactive Oxygen Species (ROS) formation (Faremi et al., 2008). The production of ROS gets prominent in state of excessive ingestion of ethanol. A diverse range of mechanisms viz., improved lipid peroxidation, protein carbonyl formation, formation of 1-hydroxyl ethyl radicals, formation of lipid radicals and diminished hepatic antioxidant defense potentials, particularly glutathione (GSH) were well studied in ethanol induced liver injury (Nwozo and Oyinloye, 2011). During alcohol consumption, various interventions have been promoted to neutralize the liver susceptibility to oxidative challenges by strengthening the endogenous antioxidant defence abilities (Fakurazi et al., 2008). Hence, toxicity of liver is considered as one of the foremost health problem to worldwide extent.
In modern system of medicine, valuable and successful drugs are not available to provoke the liver functions and safeguard the liver against various damages (Pattanayak et al., 2011). Therefore, the potential of antioxidant and hepatoprotective activity of plants were explored using diverse range of toxicants in experimental animals. The hepatoprotective nature of drugs and plant extracts were well documented through alcohol-induced hepatotoxicity model in recent years. Generally, some bioactive compounds found in plants were responsible to safe guard cells from oxidative stress via prevention or detoxification of free radicals and helps to prevent various disordered functions (You et al., 2010).
Lippia nodiflora L. (Verbenaceae), a creeping perennial herb, grows in maritime areas near rivers and it is distributed in India (Durairaj et al., 2007), Bangladesh (Ahmed et al., 2004), Baluchistan, South Africa and Central America (Terblanche and Kornelius, 1996). The plant exhibits anodyne, cardiotonic, antibacterial, diuretic, antilithic, parasiticide and refrigerant properties (Dodoala et al., 2010). Previous reports showed its effect in the treatment of ulcers, wounds, asthma, thirst, bronchitis and loss of consciousness (Kirthikar and Basu, 1975). Many chemical constituents like flavone glycosides including lippiflorin A and lippiflorin B, nodiflorin A and B, alkaloids, flavonoids like nepetin, jaceosidin, hispidulin, 6-hydroxyluteolin, flavone monosulfates and flavone disulfates, essential oil, stigmasterol, beta-sitosterol has also been reported (Tomas-Barberan et al., 1987). Many researchers have reported several pharmacological properties of Lippia nodiflora including antispasmodic (Bhakuni et al., 1969), antimalarial (Mukherjee, 1991), anti-inflammatory, analgesic and antipyretic (Forestieri et al., 1996), antinociceptive (Ahmed et al., 2004), anti-Helicobacter pylori activity (Wang and Huang, 2005), gastroprotective (Khalil et al., 2007), antibacterial (Durairaj et al., 2007), antioxidant, free radical scavenging and protective effect (Durairaj et al., 2008), diuretic effect (Shukla et al., 2009) and antidiabetic (Balamurugan and Ignacimuthu, 2011). In view of the above findings, the present study was undertaken to evaluate the protective effect of crude flavonoid fractions of L. nodiflora against ethanol-induced toxicity in rats.
MATERIALS AND METHODS
Chemicals: Reduced glutathione (GSH), oxidized glutathione (GSSG), glutathione reductase, Bovine Serum Albumin (BSA), 1,2-dithio-bisnitrobenzoicacid (DTNB), 1-chloro-2,4-dinitrobenzene (CDNB), thiobarbituricacid (TBA), picric acid, sodium tungstate, sodium hydroxide, trichloroaceticacid (TCA) and silymarin were purchased from Sigma Chemicals Co., Mumbai and all other chemicals used in this study were analytical grade with high purity.
Plant collection and preparation of extract: The whole plants were freshly collected from Karaikudi, Sivagangai District, Tamil Nadu. The plant was taxonomically identified and authenticated by Dr. G.V.S. Murthy, Joint Director, Botanical Survey of India, Tamil Nadu Agricultural University Campus, Coimbatore and the voucher specimen was deposited there with register number BSI/SRC/5/23/2012-13/Tech-19. The collected plant materials were washed, shade dried and one kilogram of coarse powder of dried aerial parts of the plant was extracted with 90% methanol in a Soxhlet extractor for 72 h. The extract was concentrated in a rotary evaporator to yield a syrupy mass (18.38 g) and was carefully scraped into a clean sample bottle and stored at 4°C for further use.
Fractionation of plant extract: The separation of crude flavonoid fraction was carried out in a similar way to the method described by Ibrahim et al. (2008). The methanol extract obtained was dissolved in hot water and filtered. The aqueous filtrate was defatted by using petroleum ether in separating funnel. The ether layer was discarded and the aqueous portion was separated and collected. The collected aqueous portion was fractionated with N-butanol saturated water and this fraction was separated, collected and the aqueous portion was discarded. The N-butanol portion was fractionated with 1% KOH and KOH portion was collected by discarding the N-butanol. The collected KOH portion was fractionated with diluted HCl and N-butanol saturated water. Finally, the diluted HCl portion was discarded to collect the N-butanol portion which serves as crude flavonoid fraction. The crude flavonoid fraction was evaporated to dryness on water bath and the obtained yield was tested for flavonoids according to standard phytochemical method (Kokate et al., 1995).
Animals: Male albino Wistar rats (150-170 g) were used for the experiment. The rats were procured from National Institute of Nutrition, Hyderabad, India. The animals were housed in polypropylene cages with sterile, inert husk materials as bedding. The experimental rats were maintained under controlled conditions (12 h light and dark cycle, temperature 22±10°C and relative humidity 40-70%). The rats were fed with a balanced commercial diet (Hindustan Lever Ltd., Mumbai, India) and water ad libitum. All animal experiments were performed according to the ethical guidelines suggested by the Committee for the purpose of Control and Supervision of experiments on animals, Government of India.
Experimental design: After 2 weeks of acclimatization, the rats were allocated randomly to five experimental groups of six animals each. Group I (C) served as control and received 0.9% saline (5 mL kg-1, b.wt.). Group II (EC) ethanol treated control received ethanol (5 g kg-1 b.wt. 20% v/v). Group III (ECCFF1) received 20% ethanol and crude flavonoid fraction of L .nodiflora (25 mg kg-1), while Group IV (ECCFF2) received 20% ethanol and crude flavonoid fraction of L. nodiflora (50 mg kg-1). Group V (ECPC) served as positive control and received 20% ethanol and silymarin (25 mg kg-1), the known standard drug.
The administration was done repeatedly on a daily basis for three weeks using metal oropharyngeal cannula. At the end of the experimental period of 3 weeks (i.e., on 22nd day), all rats from each group were sacrificed by cervical decapitation and various biochemical parameters were analysed. Fresh blood was immediately collected by cardiac puncture in fresh sterilized tubes, allowed to clot and the serum was removed by centrifugation at 2500 g at 30°C for 10 min. The sterile, haemolysis-free serum samples were kept at 4°C before determination of the biochemical analysis.
Biochemical analysis: The sterile, haemolysis-free serum was used for the assay of marker enzymes. The activities of serum aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were assayed by the method of Reitman and Frankel (1957). Alkaline phosphatase (ALP) was determined by the method of Kind and King (1954). The enzyme lactate dehydrogenase (LDH) was analyzed by the method of King (1965). The biochemical parameter such as total protein was estimated by the method of Reinhold (1953). The total bilirubin was estimated by Malloy and Evelyn (1937) method. Triglyceride was estimated by the method of Fossati and Lorenzo (1983) and urea concentration was determined by the method of Bousquet et al. (1971).
At the end of treatment period, all animals were sacrificed and the liver tissue of each animal was dissected out, washed with ice-cold saline and was homogenized in phosphate buffer (pH 7.4) to give 20% w/v homogenate. This homogenate was centrifuged at 1700 g at 4°C for 10 min and the supernatant was used for the determination of enzyme activities. Lipid peroxidation, as measured by malondialdehyde (MDA) content, was assayed by the TBA method of Buege and Aust (1978). The level of lipid peroxide was expressed as nmol MDA per gram tissue. Superoxide dismutase (SOD) activity was measured by the xanthine oxidase method, which monitor the inhibition of nitro blue tetrazolium reduction by the sample (Sun et al., 1988). Catalase activity was estimated by the method of Aebi (1984) by measuring the rate of decomposition of hydrogen peroxide (H2O2). The level of GSH, a key intra-cellular antioxidant, was measured by the method of Akerboom and Sies (1981). Hepatic GST activity was assayed according to the method of Habig and Jakoby (1981). GR activity was assayed according to Carlberg and Mannervik (1975) as the decrease in absorbance of NADPH at 340 nm.
Statistical analysis: All results were expressed as Mean±SD, (n = 6). Results were analysed statistically by one way analysis of variance (ANOVA) followed by the Dunnett’s test. Statistical significance was considered at p<0.05.
RESULTS
The effect of L. nodiflora on serum markers of ethanol induced rats were summarized in Table 1. Rats intoxicated with ethanol alone (group II) developed hepatocellular damage as evidenced by the significant elevations of serum AST (112.07±5.04), ALT (68.04 ±7.04), ALP (136.6±5.40) and LDH (325.36±1.68) activities, when compared with control. Treatment with crude flavonoid fraction of L. nodiflora (25 and 50 mg kg-1) showed a significant decrease (p<0.05) in all the elevated serum marker levels, AST, ALT, ALP and LDH by affording a significant protection against ethanol-induced liver injury. Moreover, a significant decrease in hepatic LDH activity was evident in 50 mg kg-1 of crude flavonoid fraction treated rats relative to 25 mg kg-1 treated animals. The results obtained were statistically significant and comparable to the silymarin treated group (Table 1). Group I and V shows no significant variation in ALT levels, whereas silymarin produced a significant reduction in AST (84.06±7.16 IU L-1), ALP (74.03±2.01 K.A), LDH (230.17±1.17 IU L-1), when compared with ethanol treated group.
| Table 1: | Effect of crude flavonoid fraction of Lippia nodiflora L. on serum markers in the control and ethanol-induced hepatic damaged rats |
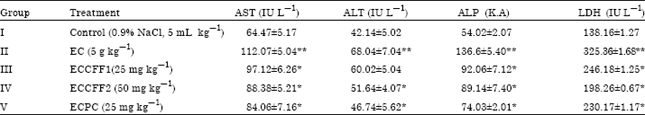 | |
| AST: Aspartate aminotransferase, ALT: Alanine aminotransferase, ALP: Alkaline phosphatase, LDH: Lactate dehydrogenase, Values are Mean±SD, n = 6, *Significant reduction compared to EC group (p<0.05), **Significant increase compared to control group (p<0.05) | |
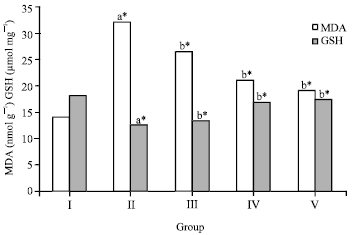 | |
| Fig. 1: | Effect of crude flavonoid fraction of Lippia nodiflora L. on lipid peroxidation (MDA) and reduced glutathione (GSH) in ethanol-induced hepatic damaged rats, Values are Mean±SD, (n = 6), *p<0.05 compared to control group (one way ANOVA followed by Dunnett’s test), where aStatistically significant when compare to control group, bStatistically significant when compare to ethanol treated group |
| Table 2: | Effect of crude flavonoid fraction of Lippia nodiflora L. on biochemical parameters in the control and ethanol-induced hepatic damaged rats |
 | |
| Values are Mean±SD, n = 6, *Significant reduction compared to EC group (p<0.05), **Significant increase compared to EC group (p<0.05), #Significant reduction compared to control group (p<0.05), ##Significant increase compared to control group (p<0.05) | |
The results of changes in serum bilirubin and urea levels were given in Table 2. The bilirubin and urea levels were decreased by 22% and 16% in group III and 31% and 30% in group IV animals; values are greater than that achieved by the standard drug. There was a significant increase in total protein (5.12±0.15 g dL-1), (6.24±0.78 g dL-1) and triglyceride (112.3±1.06 mg dL-1), (128.0±1.48 mg dL-1) levels in the group III and IV treated animals (p<0.05) when compared with the group II ethanol intoxicated rats, which exhibit total protein (4.67±0.28 g dL-1) and triglyceride (98.2±2.01 mg dL-1) levels, respectively (Table 2).
The levels of Thiobarbituric Acid Reactive Substances (TBARS) in liver tissues of ethanol intoxicated rats were significantly elevated when compared to the level of TBARS in control rats. Treatment with two different doses of crude flavonoid fraction (25 and 50 mg kg-1) showed significant (p<0.05) decrease in concentrations of TBARS compared to the ethanol treated group. However, maximum protection was offered by 50 mg kg-1 of crude flavonoid fraction. On the other hand, the standard drug silymarin also maintained the lipid peroxidation level to the normal limits than the ethanol treated group (Fig. 1). The concentration of hepatic GSH in the ethanol treated group was decreased from 18.12±0.24 to 12.46±0.17. However, treatment with crude flavonoid fraction of L. nodiflora (group III and IV) prevented the ethanol induced decline in GSH level and group IV caused 34% increase in GSH content (p<0.05) compared to ethanol treated group and restored near to its normal level. A significant increase in GSH level was evident in the silymarin treated group relative to group IV and the results were presented in the Fig. 1.
Compared to control group, hepatic GST and GR activities were also significantly decreased in the ethanol treated groups (Fig. 2), whereas no statistical alteration in GST activity was found in the silymarin treated rats. The decreased activities of GR and GST were significantly enhanced (p<0.05) by administration of 50 mg kg-1 (group IV) of crude flavonoid fraction while both the activity was only slightly changed with 25 mg kg-1 (group III) treatment in the liver tissue. These antioxidant enzymes were under normal limits in the group V animals which were intoxicated by ethanol and treated by silymarin.
The effect o f crude flavonoid fraction of L. nodiflora on SOD and CAT activities in the liver of ethanol treated animals were presented in Fig. 3.
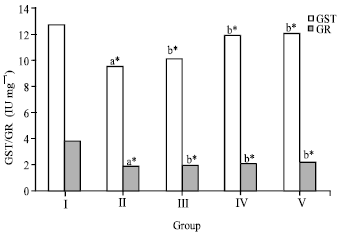 | |
| Fig. 2: | Effect of crude flavonoid fraction of Lippia nodiflora L. on glutathione-S-transferase (GST) and glutathione reductase (GR) in ethanol-induced hepatic damage rats, Values are Mean±SD, n = 6, *p<0.05 compared to control group (one way ANOVA followed by Dunnett’s test), where aStatistically significant compare to control group, bStatistically significant compare to ethanol treated group |
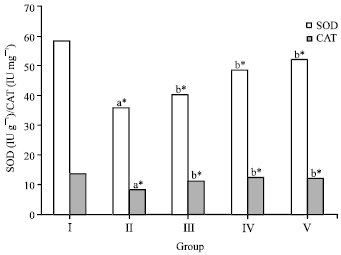 | |
| Fig. 3: | Effect of crude flavonoid fraction of Lippia nodiflora L. on superoxide dismutase (SOD) and catalase (CAT) in ethanol-induced hepatic damaged rats, Value are Mean±SD, n = 6, *p<0.05 compared to control group (one way ANOVA followed by Dunnett’s test), Where, aStatistically significant compare to control group, bStatistically significant compare to ethanol treated group |
SOD and CAT activities were decreased from 58.09±0.18 to 36.16±0.36 IU g-1 (for SOD) and from 13.45±1.09 to 8.07±0.92 IU mg-1(for CAT) in liver tissues of ethanol intoxicated rats. The reduced SOD and CAT activities were increased by 33 and 54%, respectively after administration of 50 mg kg-1 of crude flavonoid fraction. Whereas treatment with 25 mg kg-1 of crude flavonoid fraction (group III) show less significance (p<0.05) in the SOD and CAT levels as compared to the group IV animals.
DISCUSSION
The metabolic activity of the liver and other organs gets altered by alcohol consumption and therefore paves way to the development of alcoholic liver disease (Nordmann et al., 1992). Acetaldehyde and Reactive Oxygen Species (ROS) were accountable for ethanol-induced hepatic tissue damage (Zima et al., 2001). The vital steps in the prevention of alcohol-related liver diseases include the removal and neutralization of deleterious metabolites like ROS (Faremi et al., 2008). The present study concentrated on determining changes in the hepatic antioxidation defense system in association with oxidative stress, using animal models claimed with ethanol. The seepage of enzymes signifies the excellent relationship with the cellular damage (Sehrawat and Sultana, 2006). The most perceptive markers engaged in the finding of hepatic injury caused by ethanol includes serum AST, ALT, ALP and LDH (Saravanan and Nalini, 2008; Pradeep et al., 2010). These enzymes were cytosolic in nature, released into the circulatory system as a consequence of hepatocellular damage during ethanol administration and thus lead to the increase in the activities of these enzymes in serum. The enhanced activities of these serum marker enzymes observed in ethanol-treated rats in our study correspond to the extensive liver damage induced by the toxin. The rise in ALT activity is usually accompanied by rise in AST with hepatocellular, damage/necrosis, hepatocyte proliferation, or hepatocellular degeneration (Ravikumar et al., 2005). The ALP increases in serum reveal the pathological variation in biliary flow and also disclose the liver cell membrane damage (Plaa and Hewitt, 1989). LDH is a cytosolic enzyme mainly present in periportal hepatocytes and released when the cells are lysed by hepatotoxin. Cell injury was found to be proportional to the extent of the amount of enzyme released (Naik et al., 2004). Treatment with two different doses of crude flavonoid fraction of L. nodiflora significantly reversed the alterations in the status of these markers to normal levels, possibly by maintaining the hepatocellular membrane integrity. Recovery towards normalization suggests that crude flavonoid fraction of L. nodiflora causes parenchymal cell regeneration in liver, thus protecting membrane fragility, thereby, decreasing enzyme leakage.
In this present study, it was noted that the administration of ethanol decreased the levels of total protein and triglycerides. The most important cause of metabolic dysfunction that occurs during pathogenesis relates to the site-specific oxidative damage in some responsible amino acids of proteins (Bandyopadhyay et al., 1999). Hence the decline in total protein content can be considered as an useful indicator of the severity of cellular dysfunction in chronic liver diseases.
These parameters were brought back to the normal levels in the group III and IV animals, indicating the hepatoprotective nature of the drug. The rise in protein levels attributed to the stabilization of the endoplasmic reticulum which in turn leads to protein synthesis (Tanna et al., 2009). Stabilization of serum protein and triglyceride levels in the treated groups administered with crude flavonoid fraction is further a clear indication of the improvement of the functional status of the liver cells.
Serum bilirubin levels are also well associated with the status and the function of hepatic cells. The serum bilirubin level is elevated normally in hepatotoxicity (Mondal et al., 2005) and it is the probable marker of liver diseases (Achliya et al., 2004). In the present study, the marked elevation of serum bilirubin levels in group II ethanol intoxicated rats were decreased significantly by treatment with both (25 and 50 mg kg-1) doses of crude flavonoid fraction. Similarly the serum urea levels were also decreased in group III and IV treated rats. The reason for the restoration of these biochemical markers may be due to the inhibitory effects on cytochrome P450 or promotion of its glucuronidation (Cavin et al., 2001).
The liver marker enzymes increasing activity is the indicative of hepatic injury or hepatic damage (Weber et al., 2003). The excessive ROS generated during ethanol metabolism rapidly react with lipid membranes and thus initiates the lipid peroxidation chain reaction, resulting in lipid peroxyl radicals’ formation (Nordmann et al., 1992). The elevation of lipid peroxidation caused by ethanol has been previously reported in a mouse model (Balasubramaniyan et al., 2003), which is in concurrence with the results obtained in this study. Since membrane lipids are more prone to ROS, the status of Lipid Per Oxidation (LPO) has been widely used as a sign of oxidative stress (Sayeed et al., 2003). In the present investigation, the MDA production was significantly lower on crude flavonoid fraction treated groups, apparently indicating the protective role against ethanol induced damage on cell membranes (Fig. 1). Moreover, 50 mg kg-1 dose of crude flavonoid fraction revealed a strong inhibitory ability towards lipid peroxidation with an activity of 34.44% than 25 mg kg-1 dose (17.70%).
The antioxidant defense system in the body plays an important role in protection against oxidative stress. The formation of lipid peroxides and ROS gets elevated due to administration of ethanol, which primarily causes the inactivation of enzymatic and non-enzymatic antioxidants in the liver. GSH plays an important role in the antioxidant effects, nutrient metabolism and regulation of various cellular events including cell differentiation, proliferation and apoptosis (Wu and Cederbaum, 2003). GSH depletion is one of the principal factors that pave way to lipid peroxidation in cell membranes and thus disturbs the functional reliability of the cellular structure (Konukoglu et al., 1998). In our study, the GSH levels were decreased in the ethanol intoxicated rats as compared to control (Fig. 1). The decreased GSH level may be due to increased level of lipid oxidation products and it is well associated with decreased GR activity in our study. Consequently, the decrease in the GSH level could be due to its utilization to counteract the effects of ROS. The detoxification mechanisms promoted by enhanced glutathione production signifies the protective effects of L. nodiflora. This also might be the reason for the restoration of other antioxidant enzymes such as SOD and CAT.
GST represents one of the major cellular defence mechanisms against electrophilic xenobiotics and their metabolites. The conversion of oxidized glutathione (GSSG) to reduced glutathione (GSH) by the oxidation of NADPH to NADP+ takes place in the presence of Glutathione Reductase (GR), a glutathione regenerating enzyme. In the present study GST, GR activities were significantly decreased in the liver of rats exposed to ethanol as compared to control rats, which confirms previous studies (Balasubramaniyan et al., 2003; Sultana et al., 2005). The activities of GST and GR were declined on ethanol exposure and it may be due to the probable contribution of these enzymes in the detoxification and repair mechanism in liver. Treatment with crude flavonoid fraction of L. nodiflora maintains the activities of GR and GST in the liver (Fig. 2) and the stimulation of these enzymes has been assessed in a way to determine the potency of many antioxidant substances (Singh et al., 2006). With this framework, L. nodiflora is known to suppress the formation of reactive oxygen species and enhance the status of these antioxidant enzymes.
SOD prevents the formation of H2O2 by catalyzing the removal of superoxide anion radicals. The reduction in SOD activity may leads to increased H2O2 production which in turn prevents the formation of hydroxyl radicals and thus protects the visceral organs from oxidative damage (Albano, 2006). The marked reduction in the activity of SOD in rats administered ethanol and the enhanced SOD activities when crude flavonoid fraction was administered were also in agreement with other studies (Das et al., 2008; Adaramoye et al., 2009). Both doses of crude flavonoid fraction of L. nodiflora considerably increased the ethanol-induced decrease in the activity of SOD by 11.11 and 33%, respectively (Fig. 3). Catalase (CAT) is an important factor of the antioxidant defense system, widely distributed in all animal tissues and its activity is found higher in the red cells and in liver (Singanan et al., 2007). The activity of catalase was found to be decreased in ethanol intoxicated rats. The inhibition of catalase activity during ethanol induced toxicity may be due to the increased generation of reactive free radicals, which can create an oxidative stress in the cells. The administration of both doses of crude flavonoid fraction of L. nodiflora inversed the catalase activity in the liver tissues and protected from the free radical induced oxidative stress. Durairaj et al. (2008) reported that methanol extract of L. nodiflora prevent lipid peroxidation and replenish hepatic antioxidant enzymes against paracetamol induced liver damage.
It has been recognized that the use of natural products is an important preventive approach to minimize the pathological consequences of oxidative stress. Our interpretation revealed that administration of crude flavonoid fraction of L. nodiflora improved the impaired antioxidative defense system in rat livers claimed with ethanol, as indicated by the restoration of enzyme activities. Many flavonoids act as antioxidants. L. nodiflora contains some flavonoids like nepetin, jaceosidin, hispidulin, 6-hydroxyluteolin, flavone monosulfates and flavone disulfates (Tomas-Barberan et al., 1987). The protective effects of some flavonoids against oxidative stress were reported; luteolin and luteolin-7-O-glucoside as ROS scavenger (Lourdes, et al., 2008). Studies are ongoing to identify the particular bioactive molecule in L. nodiflora against ethanol-induced toxicity.
CONCLUSION
The results of our findings clearly demonstrate that there were significant variations in the observed biochemical parameters. The level of TBARS in ethanol intoxicated rats increased two fold when compared with the control. The supplementation of crude flavonoid fraction of L. nodiflora antagonizes the ethanol-induced hepatic injury as evidenced by a reversal of hepatic antioxidant status and lipid peroxidation. It is supposed that the antioxidant-sparing action of the crude flavonoid fraction of L. nodiflora could be responsible for the amelioration of oxidative stress during ethanol toxicity. Therefore, the results of the present study suggest that L. nodiflora can function as an effective free-radical scavenger, thereby preventing hepatic injury and exhibit comparable therapeutic value with the standard drug silymarin. The crude flavonoid fraction merits further investigation to identify the active principles responsible for its protective properties.
REFERENCES
- Achliya, G.S., S.G. Wadodkar and A.K. Dorle, 2004. Evaluation of hepatoprotective effect of Amalkadi Ghrita against carbon tetrachloride-induced hepatic damage in rats. J. Ethnopharmacol., 90: 229-232.
CrossRef - Adaramoye, O.A., I. Awogbindin and J.O. Okusaga, 2009. Effect of kolaviron, a biflavonoid complex from Garcinia kola seeds, on ethanol-induced oxidative stress in liver of adult Wistar rats. J. Med. Food, 12: 584-590.
CrossRefDirect Link - Ahmed, F., M.S.T. Selim, A.K. Das and M.S.K. Choudhuri, 2004. Anti-inflammatory and antinociceptive activities of Lippia nodiflora Linn. Pharmazie, 59: 329-330.
Direct Link - Akerboom, T.P.M. and H. Sies, 1981. Assay of glutathione, glutathione disulfide and glutathione mixed disulfides in biological samples. Methods Enzymol., 77: 373-382.
CrossRef - Albano, E., 2006. Alcohol, oxidative stress and free radical damage. Proc. Nutr. Soc., 65: 278-290.
CrossRefDirect Link - Balamurugan, R. and S. Ignacimuthu, 2011. Antidiabetic and hypolipidemic effect of methanol extract of Lippia nodiflora L. in streptozotocin induced diabetic rats. Asian Pac. J. Trop. Biomed., 1: S30-S36.
CrossRef - Balasubramaniyan, V., J.K. Sailaja and N. Nalini, 2003. Role of leptin on alcohol-induced oxidative stress in Swiss mice. Pharmacol. Res., 47: 211-216.
CrossRefDirect Link - Bandyopadhyay, U., D. Das and R.K. Banerjee, 1999. Reactive oxygen species: Oxidative damage and pathogenesis. Curr. Sci., 77: 658-666.
Direct Link - Buege, J.A. and S.D. Aust, 1978. Microsomal Lipid, Peroxidation. In: Methods in Enzymology, Vol. 52, Flesicher, S. and L. Packer (Eds.)., Academic Press, New York, pp: 302-310.
CrossRefDirect Link - Bhakuni, D.S., M.L. Dhar, M.M. Dhar, B.N. Dhawan and B.N. Mehrotra, 1969. Screening of Indian plants for biological activity: Part II. Indian J. Exp. Biol., 7: 250-262.
Direct Link - Carlberg, I. and B. Mannervik, 1975. Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J. Biol. Chem., 250: 5475-5480.
PubMed - Caro, A.A. and A.I. Cederbaum, 2004. Oxidative stress, toxicology and pharmacology of CYP2E1. Annu. Rev. Pharmacol. Toxicol., 44: 27-42.
CrossRef - Cavin, C., K. Mace, E.A. Offord and B. Schilter, 2001. Protective effects of Coffee diterpenes against aflatoxin B1-induced genotoxicity: Mechanisms in rat and human cells. Food Chem. Toxicol., 39: 549-556.
CrossRefPubMedDirect Link - Das, S.K., S. Varadhan, L. Dhanya, S. Mukherjee and D.M. Vasudevan, 2008. Effects of chronic ethanol exposure on renal function tests and oxidative stress in kidney. Indian J. Clin. Biochem., 23: 341-344.
CrossRef - Deshmukh, P., T. Nandgude, M.S. Rathode, A. Midha and N. Jaiswal, 2008. Hepatoprotective activity of Calotropis gigantea root bark experimental liver damage induced by D-galactosamine in rats. Int. J. Pharm. Sci. Nanotechnol., 13: 281-286.
Direct Link - Dodoala, R., B. Divit, K. Koganti and V.S.R.G. Prasad, 2010. Effect of ethanolic extract of Phyla nodiflora (Linn.) Greene against calculi producing diet induced urolithiasis. Indian J. Nat. Prod. Resour., 1: 314-321.
Direct Link - Durairaj, A.K., T.S. Vaiyapuri, U.K. Mazumder and M. Gupta, 2007. Antimicrobial and lipid peroxide scavenging activity of Lippia nodiflora (Verbenaceae). Pharmacologyonline, 3: 177-189.
Direct Link - Durairaj, A.K., T.S. Vaiyapuri, M.U.K. Kanti and G. Malaya, 2008. Protective activity and antioxidant potential of Lippia nodiflora extract in paracetamol induced hepatotoxicity in rats. Iran. J. Pharmcol. Ther., 7: 83-89.
Direct Link - Fakurazi, S., I. Hairuszah, J.M. Lip and G. Shanthi, 2008. The effect of pretreatment of zerumbone on fatty liver following ethanol induced hepatotoxicity. J. Biol. Sci., 8: 1348-1351.
CrossRefDirect Link - Faremi, T.Y., S.M. Suru, M.A. Fafunso and U.E. Obioha, 2008. Hepatoprotective potentials of Phyllanthus amarus against ethanol-induced oxidative stress in rats. Food Chem. Toxicol., 46: 2658-2664.
CrossRefDirect Link - Forestieri, A.M., M.T. Monforte, S. Ragusa, A. Trovato and L. Iauk, 1996. Antiinflammatory, analgesic and antipyretic activity in rodents of plant extracts used in African medicine. Phytother. Res., 10: 100-106.
CrossRef - Habig, W.H. and W.B. Jakoby, 1981. Assays for differentiation of Glutathione S-transferases. Methods Enzymol., 77: 398-405.
CrossRefPubMedDirect Link - Ibrahim, G., S. Abdulmumin, K.Y. Musa and A.H. Yaro, 2008. Anticonvulsant activities of crude flavonoid fraction of the stem bark of Ficus sycomorus (Moraceae). J. Pharmacol. Toxicol., 3: 351-356.
CrossRefDirect Link - Khalil, H., H. Ismail, A. Taye and M. Kamel, 2007. Gastroprotective effect of Lippia nodiflora L. extracts in ethanol-induced gastric lesions. Pharmacogn. Mag., 3: 258-261.
Direct Link - Konukoglu, D., O. Serin, G.D. Kemerli, E. Serin, A. Hayırlioglu and B. Oner, 1998. A study on the carotid artery intima-media thickness and its association with lipid peroxidation. Clin. Chim. Acta, 277: 91-98.
CrossRef - Malloy, H.T. and K.A. Evelyn, 1937. The determination of bilirubin with the photometric colorimeter. J. Biol. Chem., 119: 481-490.
CrossRefDirect Link - Naik, R.S., A.M. Mujumdar and S. Ghaskadbi, 2004. Protection of liver cells from ethanol cytotoxicity by curcumin in liver slice culture in vitro. J. Ethnopharmacol., 95: 31-37.
CrossRefPubMedDirect Link - Nordmann, R., C. Ribiere and H. Rouach, 1992. Implication of free radical mechanisms in ethanol-induced cellular injury. Free Radic. Biol. Med., 12: 219-240.
PubMed - Nwozo, S.O. and B.E. Oyinloye, 2011. Hepatoprotective effect of aqueous extract of Aframomum melegueta on ethanol-induced toxicity in rats. Acta. Biochim. Pol., 58: 355-358.
PubMedDirect Link - Pradeep, K., C.V.R. Mohan, K. Gobianand and S. Karthikeyan, 2010. Protective effect of Cassia fistula Linn. on diethylnitrosamine induced hepatocellular damage and oxidative stress in ethanol pretreated rats. Biol. Res., 43: 113-125.
CrossRef - Ravikumar, V., K.S. Shivashangari and T. Devaki, 2005. Hepatoprotective activity of Tridax procumbens against D-galactosamine/lipopolysaccharide-induced hepatitis in rats. J. Ethanopharmacol., 101: 55-60.
CrossRefPubMedDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Lourdes, R.F., R.E. Jorge, B. Scott, H.R. Dea and T. Eliseo, 2008. Risks and benefits of commonly used herbal medicines in Mexico. Toxicol. Applied Pharmacol., 227: 125-135.
PubMed - Saravanan, N. and N. Nalini, 2008. Hemidesmus indicus protects against ethanol-induced liver toxicity. Cell. Mol. Biol. Lett., 13: 20-37.
CrossRef - Sayeed, I., S. Parvez, S. Pandey, B. Bin-Hafeez, R. Haque and S. Raisuddin, 2003. Oxidative stress biomarkers of exposure to deltamethrin in freshwater fish, Channa punctatus Bloch. Ecotoxicol. Environ. Saf., 56: 295-301.
CrossRefDirect Link - Sehrawat, A. and S. Sultana, 2006. Evaluation of possible mechanisms of protective role of Tamarix gallica against DEN initiated and 2-AAF promoted hepatocarcinogenesis in male Wistar rats. Life Sci., 79: 1456-1465.
CrossRef - Shukla, S., R. Patel and R. Kukkar, 2009. Study of phytochemical and diuretic potential of methanol and aqueous extracts of aerial parts of Phyla Nodiflora linn. Int. J. Pharmacy Pharm. Sci., 1: 85-91.
Direct Link - Singh, R.P., S. Banerjee, P.V.S. Kumar, K.A. Raveesha and A.R. Rao, 2006. Tinospora cordifolia induces enzymes of carcinogen/drug metabolism and antioxidant system and inhibits lipid peroxidation in mic. Phytomedicine, 13: 74-84.
CrossRefPubMedDirect Link - Sultana, R., B.S.S. Rajua, V. Sharma, P. Reddanna and P.P. Babu, 2005. Formation of acetaldehyde adducts of glutathione S-transferase A3 in the liver of animals administered alcohol chronically. Alcohol, 35: 57-66.
CrossRef - Sun, Y., L.W. Oberley and Y. Li, 1988. A simple method for clinical assay of superoxide dismutase. Clin. Chem., 34: 497-500.
CrossRefDirect Link - Tanna, A., R. Nair and S. Chanda, 2009. Assessment of anti-inflammatory and hepatoprotective potency of Polyalthia longifolia var. pendula leaf in Wistar albino rats. J. Nat. Med., 63: 80-85.
CrossRefPubMedDirect Link - Terblanche, F.C. and G. Kornelius, 1996. Essential oil constituents of the genus Lippia (Verbenaceae)-A literature review. J. Essent. Oil Res., 8: 471-485.
Direct Link - Tomas-Barberan, F.A., J.B. Harborne and R. Self, 1987. Twelve 6- oxygenated flavone sulphates from Lippia nodiflora and L. canescens. Phytochemistry, 26: 2281-2284.
Direct Link - Wang, Y.C. and T.L. Huang, 2005. Screening of anti-Helicobacter pylori herbs deriving from Taiwanese folk medicinal plants. FEMS Immunol. Med. Microbiol., 43: 295-300.
Direct Link - Weber, L.W.D., M. Boll and A. Stampfl, 2003. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol., 33: 105-136.
CrossRefPubMedDirect Link - Wu, D. and A.I. Cederbaum, 2003. Alcohol, oxidative stress and free radical damage. Alcohol Res. Health, 27: 277-284.
Direct Link - You, Y., S. Yoo, H.G. Yoon, J. Park and Y.H. Lee et al., 2010. In vitro and in vivo hepatoprotective effects of the aqueous extract from Taraxacum officinale (dandelion) root against alcohol-induced oxidative stress. Food Chem. Toxicol., 48: 1632-1637.
CrossRefDirect Link - Zhou, Z., X. Sun and Y.J. Kang, 2002. Metallothionein protection against alochol liver injury through inhibition of oxidative stress. Exp. Biol. Med., 227: 214-222.
Direct Link - Zima, T., L. Fialova, O. Mestek, M. Janebova and J. Crkovska et al., 2001. Oxidative stress, metabolism of ethanol and alcohol-related diseases. J. Biomed. Sci., 8: 59-70.
CrossRefPubMedDirect Link