
F.J. Al-Saffar
Head Section of Histology, Department of Anatomy, Histology and Embryology, Faculty of Veterinary Medicine, Baghdad University, Baghdad, Iraq
Asian Journal of Animal Sciences
Year: 2012 | Volume: 6 | Issue: 3 | Page No.: 110-121







ABSTRACT
Goat’s industry becomes very attractive in various countries and play a vital role in socio-economic organization. A different diet causes different effects in the animal performance. Therefore, this study was intended to detect the effect of different diets on the number and distribution of endocrine Gastrin (G) cells in the goat’s abomasum. Twenty four, 9 months aged local goats were equally divided into control and two treatment groups. The control group was fed a diet consisting of 60% guinea grass and 40% concentrates consist of corn, soya bean meal, calcium, minerals, vitamins and 12.8% crude protein. The first treatment group was fed with 10% guinea grass and 90% palm kernel cake consist of calcium, minerals, vitamins and 15.2% crude protein. The second treatment group was fed with a diet similar to those of the first treatment group but with addition of molybdenum 40 ppm and sulphur 100 ppm. Goats in each group were fed daily with the selected diet for 120 days and then slaughtered and their abdomens were dissected and the abomasums were removed and processed for both histological and immunohistochemistry standard procedures. Microscopic findings revealed that treated groups that fed with high protein diet showed significantly (p<0.05) higher number of G cells than those of the control goats. Fluorescein isothiocyanate technique showed G cells distributed only in the pyloric mucosa of the abomasum with specific pattern of distribution. Current data concluded that G cells density in the pyloric mucosa have different responses to the different supplemented diets.
PDF Abstract XML References Citation
Received: February 28, 2012;
Accepted: April 19, 2012;
Published: May 17, 2012
How to cite this article
F.J. Al-Saffar, 2012. Effect of Different Diets on Number and Distribution of Gastrin Cells in the Mucosa of Goat’s Abomasums. Asian Journal of Animal Sciences, 6: 110-121.
DOI: 10.3923/ajas.2012.110.121
URL: https://scialert.net/abstract/?doi=ajas.2012.110.121
DOI: 10.3923/ajas.2012.110.121
URL: https://scialert.net/abstract/?doi=ajas.2012.110.121
INTRODUCTION
Alimentary tract is considered the first functional part of the digestive system which is capable to digest and transform diets into nutrients and subsequently their absorption through the wall of the intestine. It is frequently stated as the largest endocrine organ in the body (Ahlman and Nilsson, 2001; Chaudhri et al., 2006). However, gastrointestinal hormones are not confined to the gut and many of them are delivered to their target tissue by neural and paracrine routes as well as the blood circulation. These hormones were showed characteristic effects on the digestion and subsequent absorption of food intake beside their different stimulatory effects on the secretion and growth of gastrointestinal mucosa (Guilloteau et al., 2006).
Generally, the mucosa of the stomach is provided with different well defined groups of endocrine cells. According to the different ultrastructural features of these cells, at least seven cell types were recognized, that are Enterochromaffin (EC) cell, Enterochromaffin-like (ECL) cell, somatostatin (D) cell, gastrin (G) cell, A cell, P cell and the X cell (Liu et al., 2003).
Gastrin is one of the important hormones in the gastrointestinal endocrine system. The hormone is synthesized and stored in G cells that are located in the pyloric part of the stomach and lesser amounts of this hormone is frequently produced throughout the small and large intestine (Konda et al., 1999). Gastrin is well known to stimulate the stomach to produce acid for the normal food digestion (Jain and Samuelson, 2006). Beside that gastrin was found very necessary for the normal growth of the gastrointestinal mucosa, so that any disruption in its production may evolve subsequent digestive disturbance (Yu et al., 2011).
In the majority, it was found that gastrin (G) cells are interplay with somatostatin (D) cells to conduct and regulate gastric acid secretion (Liu et al., 2005). The hypothesis is that when food enters the stomach, the protein component stimulates G cells located in the antral area of the stomach to release the hormone gastrin, which subsequently stimulates the ECL cells that are numerous in the gastric mucosa to release histamine and the latter stimulates parietal cells to secrete acid and consequent rise in gastric pH (Vigen et al., 2011). Gastric acid causes feedback inhibition of gastrin release so that inhibit further acid production. The gastrin regulatory effect was found due to its ability to binds the cholecystokinin subtype 2/gastrin receptor located on the ECL cells that secretes histamine which in turn stimulates parietal cells to secrete hydrochloric acid (Fossmark et al., 2011).
There are three active forms of gastrin hormone that are G14, G17 and G34, out of them; G17 is the most common and predominant in the gastric secretion (Guilloteau et al., 2006). This endocrine hormone is released via exocytosis from the vesicles into the blood by stimulating the synthesis and release of histamine through their binding to specific receptors on ECL cells in the oxyntic mucosa (Lindstrom et al., 2001). Beside the actions of this hormone on the cellular growth and maintenance of the gastric mucosa, it showed cellular effect on the proliferation of both gastric stem cell and ECL cells (Koh and Chen, 2000).
During the last two decades, many investigations were carried out to find out the potential relevance of the diet on the gastrin secretion in both animals and human. Dietary calcium and fat effect on the level of serum gastrin was investigated in rats (Perry et al., 1988), response of gastrin secretion to protein intake in ruminants (Floor et al., 1991) the changes of pyloric and plasma gastrin level in response to different dietary fatty acids in rats (Frank, 2000), the effect of protein supplementation on goat and sheep performance (Kabir et al., 2002) and the increased dietary protein in lambs leads to an improvement in the energy store and subsequent raise in their body weights (Kioumarsi et al., 2008).
Nowadays, it has become a practice to use Palm Kernel Cake (PKC) as feed in the goat production. It was found one of the most successful by-products to be used as feed for ruminants and non ruminants due to its high nutritive value, good percentage of crude protein and good metabolizing energy (Alimon, 2008). Palm kernel cake (PKC) is one of the important agro-industrial by-products in different countries which are obtained from the palm oil industry (Ratnasingam et al., 2008; Abdeshahian et al., 2010). Date palm is grown mainly in tropical, subtropical and arid regions of the Middle East and North Africa and that the date represents an important income for local farmers and the date industry in these countries (Ahmed and Al-Qaradawi, 2009; Al-Khayri, 2010).
Palm kernel cake is readily available in tropical countries at competitive prices. It has been widely used in ruminant diets because of its moderate digestible energy, crude protein and high fibrous contents (Chanjula et al., 2011). However, the use of PKC in the diets for small ruminants especially sheep is usually restricted due to the high contents of copper which may lead to chronic copper poisoning (Akpan et al., 2005).
Abomasum is the final gastric compartment of the stomach in ruminants. It receives gradually the broken-down food particles which then subjected to the usual digestive juices. The main function of the abomasum is the acid hydrolysis of the microbial and dietary protein and preparing them for further digestion and absorption in the small intestine. The produced gastric acid secretion is closely correlated with the amount of food that inters the abomasums (Masot et al., 2007).
Goats comprise one of the most important domestic livestock species, since they provides a good source of meat, milk, fibers and skin (Mahmoudi, 2010). Accordingly, many procedures and technologies were developed to improve their population, production (Bughio et al., 2001; Pal et al., 2011; Kioumarsi et al., 2011) and reproduction (Rahman et al., 2008; Kor and Ziaei, 2012). On the same time, many recent reports indicated that the nature of nutrition given to the goats have prominent effect on their meat quality and certainly on their general performance (Jin et al., 2012).
Most of the previous investigations paid a particular attention concerning the activity, histochemical properties and localization of G cells and gastrin hormone in the rats, cat, dogs and human, whereas, there is paucity of work that explored these aspects in the goats. The current research analyzed the effect of three different diets mainly the level of crude protein on the frequency of immunoreactive G cells and their pattern of distribution in the mucosa of goat’s abomasum.
MATERIALS AND METHODS
Animals and experimental design: Twenty-four, 9 month aged local goats were used in this study. This research project was conducted from March 2011 to August 2011 and approved by the local ethics committee. Goats were equally divided into control group, first treated group and second treated group. Goats in the control group (n = 8) were fed a diet consisting of 60% guinea grass and 40% concentrates which included corn, soya bean meal, calcium, minerals, vitamins and 12.8% crude protein. Goats in the first treated group were fed with 10% guinea grass and 90% palm kernel cake which include calcium, minerals, vitamins and 15.2% crude protein. Finally goats in the second treated group were fed with a diet similar to those of the first treated group but with addition of molybdenum 40 ppm and sulphur 100 ppm to the concentrates. Feeding of goats with the prepared diets was continuous for 120 days. On the 121th day, animals were slaughtered and their abdomens were dissected and then stomachs were removed from the carcasses of the goats. The abomasum of each animal was opened along its greater curvature and samples from cardiac, fundic, corpus and pyloric regions were collected and labelled. For the microscopic study, tissue samples were immediately immersed in 4% paraformaldehyde in Phosphate Buffer Saline (PBS) for 16 h and cryoprotected overnight in 30% sucrose before being sectioned at 10 μm on a cryostat (Ericson et al., 2009). For general morphology some samples were processed and stained with H and E stain, whereas, the most of the other were processed for immunohistochemistry study in which Fluorescein Isothiocyanate (FITC) technique was used.
Fluorescein isothiocyanate technique: Cryosections were hydrated in distilled water for 15 min and then immersed in 0.5% hydrogen peroxide for 30 min to inhibit endogenous peroxidase activity and to reduce lipid content to allow subsequently better penetration of the antisera (Masot et al., 2007). Sections were washed again in the distilled water and rinsed in 0.1 PBS (pH 7.4) for 3 changes each of 10 min. Sections were then incubated in the normal goat serum (diluted 1 to 20 with PBS) for 20 min at room temperature to block non-specific binding of antibodies (Ericson et al., 2009). Then sections were rinsed in 0.1 PBS for 3 changes each of 10 min. After that sections incubated for 24 h in the primary antisera at 4°C. The current primary antisera used were antisera raised in rabbits against gastrin cells (G17) which were diluted (1:1000) in antisera diluents (100 mL of PBS, 2.5 g of bovine serum albumin and 0.25 g sodium azide). Once more, sections were rinsed in 0.1 PBS for 3 changes each of 10 min then incubated in the secondary antisera (Goat anti-rabbit IgG Cy3 conjugate) diluted in antisera diluents (1:400) for 1 h at room temperature (Al-Saffar et al., 2011). Finally, sections were rinsed in 0.1 PBS for 3 changes each of 10 min and then mounted with Dako fluorescence mounting medium and visualized with fluorescence microscopy (Fang et al., 2012) using image analyzer microscope (Olympus, BX 51, Japan).
Statistical analysis: Statistical calculations were carried out with the SPSS 15.0 for windows software package. Data was expressed as Mean±SEM and analyzed with Analysis of Variance (ANOVA) for all groups and subsequently confirmed by using student t-test between each two groups. The numbers of immunoreactive gastrin cells were counted in 5 randomly selected fields in each slide of the 3 selected slides belong to each animal sample. Gastrin cells were counted in three different locations that are superficial (surface), middle and the basal (deep) parts of the gastric glands. Means of the counted G cells were compared between these three locations and in the other hand the average means of the G cells of each group was compared with those of the other groups (Table 1). The value of p<0.05 considered significant difference.
RESULTS
General morphology: The microscopic examination revealed that the wall of the abomasum consists of four layers that are inner mucosa, submucosa, tunica muscularis and the outer most serosa. In the pyloric part, the epithelium of the mucosa invaginated deeply into the lamina propria, forming the gastric pit. Pyloric glands are exclusively mucous type and empty into the bottom of gastric pit. The surface epithelial cells and gastric pits were also found to be mucous cells. The gastric pits were lined with a high columnar epithelium (Fig. 1a-1, 1b-1). The surface epithelium of cardiac and pylorus was made up of surface mucous cells and branched tubular mucous glands (Fig. 1d-2), whereas fundic and corpus were formed of branched tubular glands that contain several cell types. The epithelium rest over muscularis mucosa consists of few layers of smooth muscle fibers (Fig. 1c-4). The lamina properia and submucosa contains loose connective tissue, lymphoid cells and blood vessels (Fig. 1c-3). The outer wall of the abomasum contains layers of smooth muscles bundles (Fig. 1c-5). The outermost layer of the abomasum is the tunica serosa representing the visceral peritoneum which contains loose connective tissue, blood vessels and a covering of mesothelial cells.
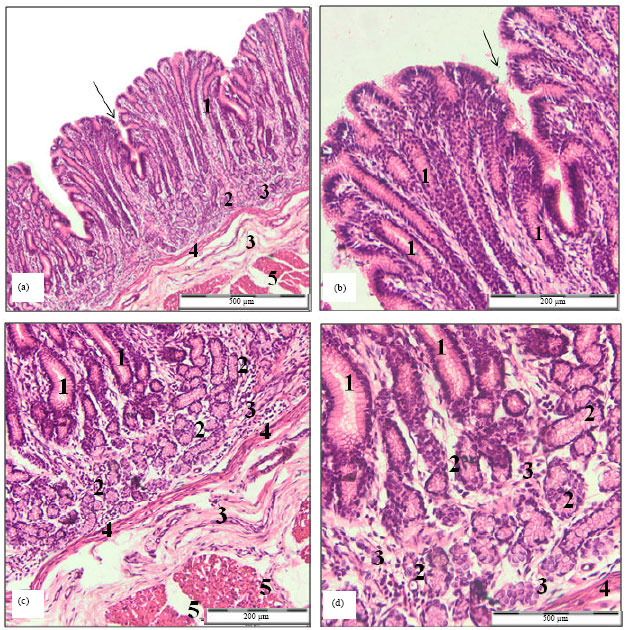 | |
| Fig. 1(a-d): | Pyloric mucosa of the goat’s abomasum showed “gastric pits” (arrow), mucus-secretory simple columnar lining epithelium (1), simple tubular mucus-secretory glands (2), loose connective tissue with blood vessels in the underlining propria-submucosa (3), smooth muscle layers of muscularis mucosa (4) tunica muscularis (5), H and E, (a) X40, (b) X100, (c) X100 and (d) X200 |
Immunohistochemistry: Microscopic examination with the fluorescence microscope showed numerous number of positive G cells distributed throughout the length of the pyloric gastric glands in the abomasums of the control and both treated groups. Whereas, the mucosa of cardiac, fundic and the corpus were characteristically lack this type of cells. The immunoreactive G cells appeared red-golden fluorescent-stained cells which were distributed in a single pattern all over the gastric glands of the pyloric mucosa and they were concentrated significantly (p<0.05) in the basal (deeper) part compared to those counted at both middle and surface (superficial part) of these glands.
| Table 1: | Number of gastrin cells in the pyloric mucosa of the control and the two treated groups of goats |
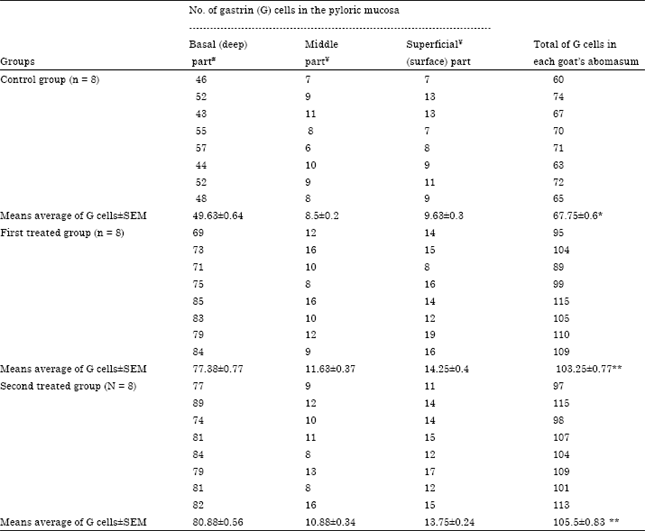 | |
| *Significantly (p<0.05) lower number of gastrin cell present in the pyloric mucosa compared to those of the first and second treated groups, **Non significant (p<0.05) difference in the number of gastrin cells present in the pyloric mucosa of the first and second treated groups, #Significantly (p<0.05) higher number of gastrin cell present in the basal (deeper) part of the pyloric mucosa compared to those of the middle and surface (superficial) epithelium, -Non significant (p<0.05) difference in the number of gastrin cells present in the middle and surface parts of the pyloric mucosa in the control, first and second treated groups | |
The lining epithelium of the gastric glands rests on an underlining propria-submucosa layer of loose connective tissue which is interrupted by smooth muscle layers of muscularis mucosa (Fig. 2-4).
The mean number of G cells observed in the control and treated groups is well publicized in the Table 1. In each group, the average number of G cells which was counted at the basal (deeper) part of the gastric glands of the pyloric mucosa was significantly (p<0.05) higher than those observed at the middle and surface (superficial) parts of the gastric glands. The number of G cells at the basal, middle and surface part of the gastric glands of the control group was 49.63±0.64, 8.5±0.2 and 9.63±0.3, respectively. Whereas, the G cells number observed in the basal, middle and surface part of the gastric glands of the first treated group was 77.38±0.77, 11.63±0.37 and 14.25±0.4, respectively. Second treated group showed no significant changes in the number of G cell compared to those of the first group and their G cells number was 80.88±0.56, 10.88±0.34 and 13.75±0.24, respectively.
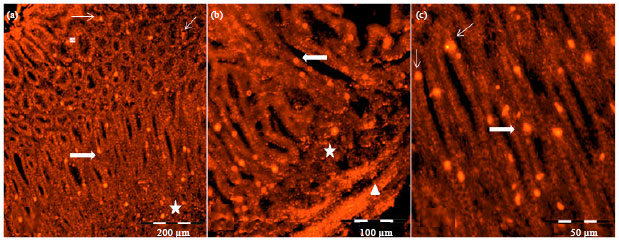 | |
| Fig. 2(a-c): | Pyloric mucosa of goat’s abomasum in the control group showed gastrin cells distributed in the surface part (thin arrow), middle part (thick arrow) and deeper part of the gastric glands (star), Loose connective tissue located in the underlining propria-submucosa which is interrupted by smooth muscle layers of muscularis mucosa (triangle), fluorescein isothiocyanate (FITC), (a) X100, (b) X200, (c) X200 |
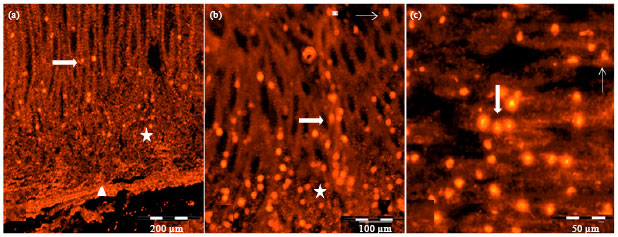 | |
| Fig. 3(a-c): | Pyloric mucosa of goat’s abomasum in the first treated group showed characteristically higher number of gastrin cells compared to the control group; distributed in the surface part (thin arrow), middle part (thick arrow) and deeper part of the gastric glands (star). Loose connective tissue located in the underlining propria-submucosa which is interrupted by smooth muscle layers of muscularis mucosa (triangle), FITC, (a) X100, (b) X200, (c) X400 |
Statistical analysis revealed that goats of both treated groups were showed significantly (p<0.05) higher number of G cells in their pyloric mucosa compared to those observed in the control animals. The average number of G cells counted in the pyloric mucosa of the control group was 67.75±0.6, whereas, the average number was significantly higher up to 103.25±0.77 and 105.5±0.83 in the first and second treated groups, respectively.
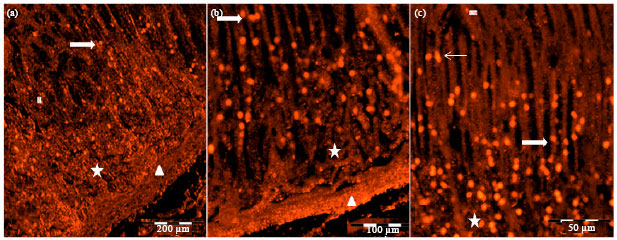 | |
| Fig. 4(a-c): | Pyloric mucosa of goat’s abomasum in the second treated group showed obviously higher number of gastrin cells compared to the control group; distributed in the surface part (thin arrow), middle part (thick arrow) and deeper part of the gastric glands (star), Loose connective tissue located in the underlining propria-submucosa which is interrupted by smooth muscle layers of muscularis mucosa (triangle), FITC, (a) X100, (b) X200, (c) X200 |
Analysis with Student t-test revealed absence of mean difference (p<0.05) between both treated groups. Finally, all groups showed no significant (p<0.05) differences in their average number of G cells distributed at both middle and surface parts of the gastric glands.
DISCUSSION
The immunoreactive G and somatostatin (D) cells were considered the major endocrine cells in the stomach that are play an important role in acid secretion (Liu et al., 2005). Previous thinking indicated that the gastrin hormone and gastrin cells play a role in gastric normal diet digestion. The present study was performed to assess the effect of different diets on the numbers and distribution of gastrin cells in the abomasum of goats. The crude protein which was 12.8% in the control groups as standards feed for goat production, raised up to 15.2% in the treated groups fed with combination of Palm Kernel Cake (PKC) and grass. The selection of palm kernel cake (PKC) based on its high nutritive value and its extensive use as feed ingredients in ruminants (Chanjula et al., 2011). Current findings revealed significant effect of the diet on the mean numbers of gastrin cells in the treated goats compared to those of the control group. Results indicated that different diets particularly the protein level does affect the numbers of gastrin cells abomasum of goats. The findings were consistence with previous observations that are considered proteins and amino acids as the most powerful gastrin secretagogues (Calbet and Holst, 2004) and are the most potent stimulator of gastrin hormone (Feng et al., 2010).
The microscopic observations of the immunofluorescence technique showed that in the goat’s abomasum, immunoreactive G cells were located and distributed in the pyloric region only and they were totally absent in the cardiac, fundic and corpus regions of the abomasum. These findings were similar to those previously observed in the sheep’s abomasum in which gastrin cells were very numerous in the abomasum and duodenum but are deficient in the cardiac or fundic regions of the abomasum (Franco et al., 1993). Similarly, previous findings confirmed that the immunoreactive gastrin cells located mainly in the middle third of the pyloric mucosa in the stomach of dogs, cats and humans (Takahashi et al., 1981; Marino et al., 1994; Galan et al., 1996).
Gastrin cells are considered essential regulators of gastric acid secretion and the alterations in its relative numbers may play a key role in gastroduodenal disease (Liu et al., 2005). The importance of G cells arises from the fact that the dietary protein intake triggers the production of gastrin hormone from these cells, which subsequently stimulates the secretion of hydrochloric acid that aids the process of digestion (Mullin et al., 2009).
Current findings showed higher number of G cells located at the basal part of abomasal pyloric glands of treated goats. These findings appeared similar to those previously observed in the rat antral mucosa, which revealed the existences of numerous G cells in the basal part of the glandular area of the stomach (Varagic et al., 1996). The present data revealed significantly lesser density of the G cells at the surface and middle parts compared to those of the basal deeper part of the pyloric glands. The difference of G cell density reflects the function of the surface mucosal protection against gastric acid secretion produced during the process of digestion. Morphological examination showed a picture which was quite clear to explain this consideration as it showed highly mucous cell secretory surface epithelium in the pyloric mucosa of all groups which were in consistency with previous findings (Suprasert et al., 1999).
CONCLUSION
The current data concluded that the protein content of the diet have certain effect on the G cells number which certainly affect the gastric acid secretion and subsequently gastric function of the goat’s abomasum. The results of fluorescein technique showed prominent effect of higher protein diet indicated by the numerous G cells appearance in the pyloric mucosa.
ACKNOWLEDGMENTS
Great respect and thanks to the staff of Anatomy, Histology and Embryology Department for their great support to achieve this research.
REFERENCES
- Abdeshahian, P., N. Samat and W.M. Wan Yusoff, 2010. Utilization of palm kernel cake for production of β-glucosidase by Aspergillus niger FTCC 5003 in solid substrate fermentation using an aerated column bioreactor. Biotechnology, 9: 17-24.
CrossRefDirect Link - Ahlman, H. and O. Nilsson, 2001. The gut as the largest endocrine organ in the body. Ann. Oncol., 12: S63-S68.
CrossRefPubMedDirect Link - Ahmed, T.A. and A.Y. Al-Qaradawi, 2009. Molecular phylogeny of Qatari date palm genotypes using simple sequence repeats markers. Biotechnology, 8: 126-131.
CrossRefDirect Link - Akpan, H.D., E.O. Udosen, A.A. Udofia, E.J. Akpan and A.A. Joshua, 2005. The effect of phytase and zinc supplementation on palm kernel cake toxicity in sheep. Pak. J. Nutr., 4: 148-153.
CrossRefDirect Link - Al-Khayri, J.M., 2010. Somatic embryogenesis of date palm (Phoenix dactylifera L.) improved by coconut water. Biotechnology, 9: 477-484.
CrossRef - Al-Saffar, F.J., S. Ganabadi, S. Fakurazi and H. Yaakub, 2011. Zerumbone significantly improved immunoreactivity in the synovium compared to Channa striatus extract in monosodium iodoacetate (MIA)-induced knee osteoarthritis in rat. J. Med. Plants Res., 5: 1701-1710.
Direct Link - Bughio, S., M.H. Baloch, F.C. Oad, A.A. Moryani, A.H. Mirani and M.A. Brohi, 2001. Productive performance of kamori goat flock under semi-intensive management. J. Applied Sci., 1: 347-348.
CrossRefDirect Link - Calbet, J.A.L. and J.J. Holst, 2004. Emptying, gastric secretion and enterogastrone response after administration of milk proteins or their peptide hydrolysates in humans. Eur. J. Nutr., 43: 127-139.
CrossRef - Chanjula, P., Y. Siriwathananukul and A. Lawpetchara, 2011. Effect of feeding rubber seed kernel and palm kernel cake in combination on nutrient utilization, rumen fermentation characteristics and microbial populations in goats fed on Briachiaria humidicola hay-based diets. Asian Aust. J. Anim. Sci., 24: 73-81.
Direct Link - Chaudhri, O., C. Small and S. Bloom, 2006. Gastrointestinal hormones regulating appetite. Phil. Trans. R. Soc. B, 361: 1187-1209.
CrossRefDirect Link - Ericson, A., E.M. Nur, F. Petersson and S. Kechagias, 2009. The Effects of capsaicin on gastrin secretion in isolated human antral glands: before and after ingestion of red chilli. Dig. Dis. Sci., 54: 491-498.
CrossRef - Fang, J., M. Ma, Y. Jin, N. Qiu, C. Wang, G. Ren and X. Huang, 2012. Assessment of salmonella enteritidis viability in egg white during early incubation stages by fluorescent staining method. Asian J. Animal Vet. Adv., 7: 556-567.
CrossRef - Feng, J., C.D. Petersena, D.H. Coyb, J.K. Jiangc, C.J. Thomasc and S.A. Wanka, 2010. Calcium-sensing receptor is a physiologic multimodal chemosensor regulating gastric G-cell growth and gastrin secretion. Proc. Natl. Acad. Sci., 107: 17791-17796.
CrossRef - Floor, M.K., S. Jahangeer, C. D'Ambrosio and O. Alabaster, 1991. Serum gastrin increases with increasing dietary calcium but not with increasing dietary fat or fiber in fischer-344 rats. J. Nutr., 121: 863-868.
PubMed - Fossmark, R., G. Qvigstad, T. Martinsen, Y. Hauso and H.L.Waldum, 2011. Animal models to study the role of long-term hypergastrinemia in gastric carcinogenesis. J. Biomed. Biotechnol., Vol. 2011.
CrossRef - Frank, P.N., 2000. The effect of different dietary fats on gastrin levels in the pyloric antrum and plasma of weaner and adult wistar rats. Afr. J. Med. Sci., 29: 135-139.
PubMed - Franco, A, S. Regodon, A. Gazquez, A.J. Masot and E. Redondo, 1993. Ontogeny and distribution of gastrin cells in the gastrointestinal tract of the sheep. Eur. J. Histochem., 37: 83-90.
PubMed - Galan, J.A., F.J. Alonso, P. Moratinos, J.L. Gonzalez, B. Fraile and M.V. Lobo, 1996. The G-cells in the dog: A light and electron microscope immunocytochemical study. Histochem. J., 28: 883-893.
PubMed - Guilloteau, P., V.L.M. Metzinger, J. Morisset and R. Zabielski, 2006. Gastrin, cholecystokinin and gastrointestinal tract functions in mammals. Nutr. Res. Rev., 19: 254-283.
PubMed - Kabir, F., M. Shahjalal, S.A. Chowdhury, J. Alam and M.R. Islam, 2002. Effect of protein supplementation to grazing on growth and reproductive performance in female goats and sheep. Pak. J. Biol. Sci., 5: 719-721.
CrossRefDirect Link - Kioumarsi, H., K.J. Khorshidi, M. Zahedifar, A.R. Seidavi, S.Z. Mirhosseini and M.R. Taherzadeh, 2008. The effect of dietary energy and protein level on performance, efficiency and carcass characteristics of Taleshi lambs. Asian J. Anim. Vet. Adv., 3: 307-313.
CrossRefDirect Link - Kioumarsi, H., Z.S. Yahaya, W.A. Rahman and P. Chandrawathani, 2011. A new strategy that can improve commercial productivity of raising boer goats in Malaysia. Asian J. Anim. Vet. Adv., 6: 476-481.
CrossRefDirect Link - Konda, Y., H. Kamimura, H. Yokota, N. Hayashi, K. Sugano and T. Takeuchi, 1999. Gastrin stimulates the growth of gastric pit with less-differentiated features. Am. J. Physiol., 277: G773-G784.
PubMed - Kor, N.M. and N. Ziaei, 2012. Effect of PGF2α administration and subsequent eCG treatments on the reproductive performance in mature raieni goats during the breeding season. Asian J. Anim. Vet. Adv., 7: 94-99.
CrossRefDirect Link - Jain, R.N. and L.C. Samuelson, 2006. Differentiation of the Gastric Mucosa II. Role of gastrin in gastric epithelial cell proliferation and maturation. Am. J. Physiol., 291: G762-G765.
CrossRef - Jin, H., H.Q. Chen, J. Qin, Y.J. Zhu and H. Chen et al., 2012. The polymorphism in 5 regulatory region and exon 13 of PRKAG3 gene and its distribution pattern in different goat breeds. Asian J. Animal Vet. Adv., 7: 568-577.
CrossRefDirect Link - Lindstrom, E, D. Chen, P. Norlen, K. Andersson and R. Hakanson, 2001. Control of gastric acid secretion: The gastrin-ECL cell-parietal cell axis. Comp. Biochem. Physiol. A Mol. Integr. Physiol., 128: 503-511.
CrossRef - Liu, Y., G.D.C. Vosmaer, G.N.J. Tytgat, S.D. Xiao and F.J.W. Kate, 2005. Gastrin (G) cells and somatostatin (D) cells in patients with dyspeptic symptoms: Helicobacter pylori associated and non-associated gastritis. J. Clin. Pathol., 58: 927-931.
CrossRef - Mahmoudi, B., 2010. Genetic diversity of lori goat population based on microsatellite marker. Asian J. Anim. Sci., 4: 13-19.
CrossRefDirect Link - Marino, L., B. Muglia and C. Dickinson, 1994. Glycine-extended post-translational processing intermediates of gastrin and cholecystokinin in the gut. Regul. Pept., 50: 73-85.
CrossRef - Masot, A.J., A.J. Franco and E. Redondo, 2007. Morphometric and immunohistochemical study of the abomasum of red deer during prenatal development. J. Anat., 211: 376-386.
CrossRef - Pal, U.K., P.K. Mandal, V.K. Rao and C.D. Das, 2011. Quality and utility of goat milk with special reference to India: An overview. Asian J. Anim. Sci., 5: 56-63.
CrossRefDirect Link - Perry, K.W., T.E.C. Weekes, J.A. Rooke, D.S. Parker and D.G. Armstrong, 1988. Effect of protein intake on gastrin secretion in ruminants. Q. J. Exp. Physiol., 73: 985-993.
PubMed - Rahman, A.N.M.A., R.B. Abdullah and W.E.W. Khadijah, 2008. A review of reproductive biotechnologies and their application in goat. Biotechnology, 7: 371-384.
CrossRefDirect Link - Ratnasingam, J., T. McNulty and M. Manikam, 2008. The machining characteristics of oil palm empty fruit bunch particleboard and its suitability for furniture. Asian J. Applied Sci., 1: 253-258.
CrossRefDirect Link - Suprasert, A., U. Pongchairerk, P. Pongket and T. Nishida, 1999. Lectin histochemical characterization of glycoconjugates present in abomasal epithelium of the goat. Kasetsart J., 33: 234-242.
Direct Link - Takahashi, T., M. Tani and H. Shimazu, 1981. Effect of truncal vagotomy on tissue gastrin content and G cell density in rats. Gastroenterol. Jpn., 16: 71-75.
PubMed - Vigen, R.A., D. Chen, U. Syversen, K. Stunes, R. Hakanson and C.M. Zhao, 2011. Serum gastrin and enterochromaffin-like cells during estrous cycle, pregnancy and lactation, and in response to estrogen-like agents in rats. J. Physiol. Pharmacol., 63: 335-340.
PubMed - Varagic, J., V. Koko, V. Todorovic, Z. Bozovic, V. Lackovic and V. Vuzevsk, 1996. The effect of long-term low-protein intake on gastrin cells of the rat antral mucosa during adulthood. Histol. Histopathol., 11: 837-843.
PubMed - Yu, H.G., J.O. Nam, N.L.G. Miller, I. Tanjoni and C. Walsh et al., 2011. P190RhoGEF (Rgnef) promotes colon carcinoma tumor progression via interaction with focal adhesion kinase. Cancer Res., 71: 360-370.
CrossRef