
G.S.I. Wogar
Department of Animal Science, University of Calabar, Calabar, Nigeria
Asian Journal of Animal Sciences
Year: 2011 | Volume: 5 | Issue: 6 | Page No.: 373-380







ABSTRACT
This study was conducted to determine the performance of gestating grasscutters on graded crude protein diets formulated from by-products of agricultural production. The aim of the study was to determine the protein requirement and performance of gestating grasscutters on diets formulated from wheat offal and soybean meal. Cassava was used as a source of energy, while its starch served as a binding agent in the pelleted diets. Sixteen 8 months old pregnant grasscutters of equal weight were allotted to four treatment groups with four animals per group. Each group was randomly allotted to one of four treatment diets formulated to supply 10, 14, 18 and 22% Crude Protein (CP), respectively. The gestating grasscutters were assessed for growth and reproductive performance. Data was collected for the twenty (20) weeks of gestation. The performance of gestating grasscutters on the 14% CP diet in respect of weight at end of gestation and daily weight gain was significantly (p<0.05) higher than on other diets. The results also showed that feed intake and average litter size were highest on the 14% diet. Performance in respect of feed conversion ratio, cost to gain ratio and average birth weight of pups was not significantly different between the 14% and 22% CP diets. These findings suggest that the optimum dietary crude protein level for rearing gestating grasscutters is 14%, which level of crude protein could be supplied from the locally available and low-cost agro-industrial by-products used in this study.
PDF Abstract XML References Citation
Received: July 19, 2011;
Accepted: August 29, 2011;
Published: December 01, 2011
How to cite this article
G.S.I. Wogar, 2011. Performance of Gestating Grasscutters (Thryonomys swinderianus) Fed Cassava-based Diets with Graded Protein Levels. Asian Journal of Animal Sciences, 5: 373-380.
DOI: 10.3923/ajas.2011.373.380
URL: https://scialert.net/abstract/?doi=ajas.2011.373.380
DOI: 10.3923/ajas.2011.373.380
URL: https://scialert.net/abstract/?doi=ajas.2011.373.380
INTRODUCTION
The grasscutter is a wild herbivorous rodent, which lives mainly on grass and other succulent forages. It is related to other herbivorous rodents like the African porcupine, the brush-tailed porcupine, the guinea pig and the chinchilla (Baptist and Mensah, 1986; NRC, 1991). Grasscutter meat has been reported to have higher nutritional value (Opara, 2010a) and meat yield (Omole et al., 2005) than meat from traditional livestock species. However, Abulude (2007) found the nutritional qualities of bush meat to be comparable to those of conventional meats. Grasscutter meat tops the bushmeat (generic for all meats from wildlife species) trade and contributes to meet the protein needs of the populations of West African countries. An estimated 40,000 tonnes per year of grasscutter meat is consumed in West Africa, of which only 0.2% is produced in captivity (Mensah and Okeyo, 2005). The balance of the estimated annual consumption of grasscutter meat is supplied from hunting and other environmentally harmful practices like bush burning (Yeboah and Adamu, 2005; Taiwo et al., 2009).
The grasscutter is found where there is dense grass (Adoun, 1993; NRC, 1991) and where there is intensive cultivation of maize, cassava, sugar cane, pineapple and egg plant. They are also found in plantations of cocoa, coconut and oil palm (Asibey, 1974). These farms provide food all year round (Yeboah and Adamu, 1995). Various crops of these farms supply some of the protein and other nutrients for grasscutter in the wild. In its natural environment, foraging is a critical factor in the development of the genetic potential of the wild grasscutter (El-Hassan et al., 2009). In captivity, adequate protein must be supplied in grasscutter diets to meet the requirements for any form of productive activity. In order to satisfy the growing demand for grasscutter meat as well as ensure quality control and sustainable supply, it is desirable that the production of grasscutter is planned (Opara, 2010b).
The gastro-intestinal tract of grasscutter combines the functions of the monogastric and ruminant digestive tracts. It is suggested that the grasscutter would require the same 41 nutrients ordinarily required by other monogastric animals, which include 13 essential amino acids (Olomu et al., 2003). Feeds containing 12-20% CP (Meduna, 2002) have been reported to be suitable for grasscutters. This range of protein requirement is wide and does not specify the needs for various physiological states of the grasscutter. The efficiency of grasscutter production, as is the case with conventional livestock, would depend on the reproductive efficiency of the grasscutter (Chukwuka et al., 2010). Ren et al. (2005) found that the reproductive performance of another pseudo-ruminant, the rabbit, was best on a diet supplying 17.48% CP. Also, the litter size and weaning weight were improved when lactating does were fed a diet supplying 19.54% CP (Ren et al., 2005).
It has been reported that balanced feeds for grasscutters may be formulated from maize, rice bran or wheat bran, palm kernel cake meal or groundnut cake, oyster or periwinkle shells, bone meal, common salt and vitamin/mineral premix (Yi and Tewe, 1980).
The objective of this study was to determine the performance and protein requirement of pregnant grasscutters fed diets formulated from low-cost agro-industrial by-products, involving soybean meal and wheat offal as the main sources of dietary crude protein.
MATERIALS AND METHODS
The study was carried out at the Grasscutter Research Farm at Calabar, under the supervision of the Department of Animal Science, University of Calabar, Calabar, Nigeria. Calabar is at latitude 3° North and longitude 7° East. It has an annual rainfall of 2,650 to 3000 mm and relative humidity of 57 to 93%, with annual temperatures between 25 and 32°C.
Experimental diets: Four different dietary protein levels of 10 (CP 10), 14 (CP 14), 18 (CP18) and 22% (CP 22) were formulated using soybean as the main source of protein and cassava as the main source of energy. Dietary energy was equalized to 2400 kilocalories (kcal), Metabolizable Energy (ME) kg-1. All the ingredients used were purchased from the local market in Calabar. A thick paste of the ingredients was made using cassava starch as the binding agent. The paste was pelleted in a pelleting machine and dried in a kerosene-fired oven at 75°C. The composition of the test diets is shown in Table 1, while the proximate composition of the diets is shown in Table 2.
Research animals: The sixteen (16) gestating grasscutters were 7 to 8 months old and weighed between 2.3 and 2.4 kg. All the growing grasscutters had shown signs of sexual maturity by opening of the vagina.
| Table 1: | Composition of experimental diets for determination of protein requirements of gestating grasscutters |
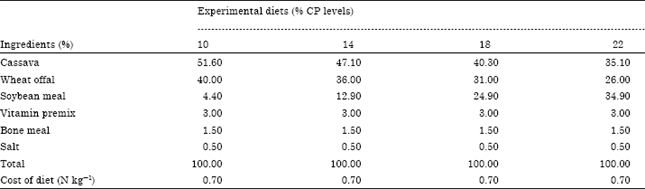 | |
| N kg-1 = Naira (Nigerian currency) per kilogram. $1.00 = N152.00 (as at 10/08/2011) | |
Management of research animals: The grasscutters were randomly allotted, in groups of four, to the four treatment diets. Each group was randomly fed one of the four dietary protein levels. There were four replicates per treatment with one (1) grasscutter per replicate in a completely randomized experimental design. The grasscutters were individually housed in clearly and properly-labeled concrete cells measuring 150x75x35 cm (lengthxwidthxheight). The housing provided for only one opening (35 highx45 cm wide) into the cell in order to eliminate cross-ventilation and prevent the adverse effect of cold on grasscutters, which are very susceptible to pneumonia. Temperatures in the cells were in the range of 25-31°C during the experimental period. On introduction into the cells, the animals were dewormed and provided with anti stress agents in drinking water. Elephant grass (Pennisetum purpureum), which has been cut and allowed to wilt for two days, was weighed and fed daily as basal diet. Water, diet and elephant grass were supplied ad libitum. The animals were weighed, at the beginning and every two weeks thereafter, during the 20 weeks of the study. All cells were cleaned daily in order to ensure an acceptable level of sanitation.
Data collection and analysis: Records of daily feed (elephant grass and diet) intake, daily weight gain of does, weight of does at end (terminal weight) of gestation, average litter size and average birth weight of pups and cost of diet, were taken. Data collected were subjected to analysis of variance using the software of Genstat (2007).
RESULTS AND DISCUSSION
The proximate composition of the experimental diets is presented in Table 2. The results show that there were no differences between diets in respect of percentage composition of dry matter and ash. There were, however, significant differences in respect of composition of crude protein, crude fibre, ether extract and nitrogen free extract. It was observed that the percentage crude fibre, ether extract and nitrogen free extract of the diets decreased with increase in the protein level of the diet. This was not unexpected as diets were formulated to satisfy the objective of the study, which was to determine the crude protein requirement of gestating grasscutters.
Growth and reproductive performance of gestating grasscutters fed graded protein levels is presented in Table 3.
Growth performance of gestating grasscutters: The daily weight gain of gestating grasscutters was significantly (p<0.05) higher (14.83 g) on the CP14 (14% CP) than on other diets.
| Table 2: | Proximate composition of experimental diets for determination of protein requirements of gestating grasscutters |
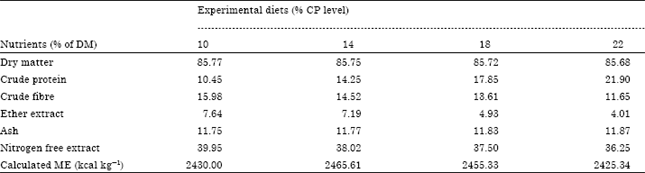 | |
| Table 3: | Performance of gestating grasscutters fed varying dietary protein levels |
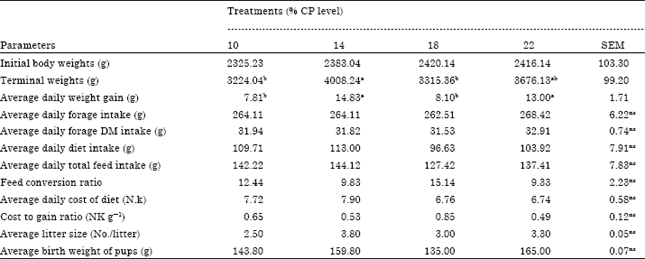 | |
| Means along the same row having no common superscript differ significantly at p<0.01; ns refers to non-significant differences between means | |
This observation indicates that the agro-industrial by-products (soybean and wheat offal) as sources of protein in the diets significantly affected weight gain in gestating grasscutters. This finding differs from the findings of Adeniji (2008) that diets supplemented with groundnut cake, soybean meal and blood meal had no significant effect on growing grasscutters. Results indicated that daily weight gain declined with increase in dietary protein levels above 14% CP. This finding indicates that higher than required dietary protein levels did not increase weight gain. The higher weight gain of gestating grasscutters on the CP 14 diet could be due to the higher levels of total feed (144.12 g) and diet (113.00 g) intake on that diet. The high feed intake could be due to the relatively high levels of crude fibre in the CP 14 diet. This is in consonance with earlier reports that high fibre diets tend to increase feed intake in rabbits (Jokthan et al., 2006). The high fibre content of the CP14 diet could have caused a decrease in the caloric density of that diet. Hence, the need for increased feed intake to meet the energy demands of the grasscutters on CP14 diet. This agrees with the report of NRC (1977) that rabbits voluntarily adjust their feed intake to satisfy energy demand. The reduced weight gain on the higher dietary protein levels could be due to the significantly lower fibre levels in those diets. This is in agreement with Taiwo et al. (2005) who observed reduced growth rates in weaned rabbits fed low fibre diets. It is suggested that the low fibre content of these diets may have prolonged retention times of digesta in the caecum resulting in reduced feed intake and weight gain (Bawa et al., 2008). The average daily weight gain which ranged from 7.81 to 14.83 g, however, agrees with the 8 to 13 g reported by Jori and Chardonnet (2001) and the 7 to 12 g reported by Mensah (1995).
Reproductive performance: Litter size and birth weight
Litter size: Reproductive performance in respect of average litter size at kindling was higher (3.80 pups) on the 14% CP diet than on other diets. The smallest litter size (2.5 pups) was observed on the CP 10 diet. The range of 2.50 to 3.80 pups per litter was within the range of 2.60 to 5.00 pups per litter reported by other studies (Addo et al., 2007; Henry, 2011). However, the average litter size (2.50 to 3.80 pups litter) at kindling was below the range of 3.8 to 5.7 pups per litter reported by Mensah (2000). Non-significant differences have been reported by Lameed and Ogundijo (2006), with litter size increasing as dietary CP levels increased from 20, 22, to 24%. The findings of this experiment suggest that the 14% CP diet was more consistent with larger litter sizes in gestating grasscutters than were other diets.
Birth Weight of Pups (BWP): The average birth weight (159.80 g) on the 14% CP diet was second highest but not significantly lowers than the highest (165.00 g) on the 22% CP diet. The lowest birth weight (135 g) was observed on the CP 18 diet. The average birth weight of 135 to 165 g obtained in this study is beyond the range of 81 to 157 g reported in other studies (Schrage and Yewadan, 1995; Addo, 2002; Henry, 2011). Lameed and Ogundijo (2006) reported significant differences between the BWP from grasscutters fed diets containing 20 to 22% CP, with mean BWP of 133.33±2.36 and 132.31±3.12 g, respectively. The significantly higher Birth Weight of Pups (BWP) observed for grasscutters fed the CP22 and CP 14 diets were consistent with the significantly higher Daily Weight Gain (DWG) and Terminal Weight (TWG) of primiparous grasscutter does on these diets. The findings of this experiment indicate that pregnant grasscutters, like pregnant rabbit does (Patridge et al., 1986; Parigi-Bini et al., 1990), moderate their maternal environment to cater for the nourishment of foetuses and maintain body condition. The findings further indicate that, as in the rabbit (Lebas et al., 1996), grasscutters are able to adjust intake according to the nutrient content of their diet in order to cater for developing foetuses. These results indicate that the 14% diet was more consistent with higher birth weights than the 10 and 18% diets but comparable with the 22% diet.
Terminal weight (weight at end of pregnancy): The terminal weight of gestating grasscutters, which is an index of maternal well-being, differed significantly (p<0.05) with dietary protein levels. Average weight at the end of gestation was higher (4008.28 g) on the 14% CP diet, than on the 10% (3224.03 g) and 22% (3676.13 g) CP diets. This observation is in agreement with the findings of Azahan and Noraziah (2011) that a medium protein diet (17.5%) was more efficient than the 12.5 and 22.5% protein diets for growing ostriches. The differences in the terminal weights of pregnant grasscutter does could be due to differences in the efficiency of adjustments required to balance the nutrients and body reserves of fat, energy and protein between the maternal body and the foetuses (De Wilde, 1980; Patridge et al., 1986; Parigi-Bini et al., 1990; Xiccato et al., 1999). The higher average weight of does at kindling, in this study, reflects more effective maternal moderations by pregnant grasscutters on the CP 14 diet to cater for the developing foetuses. Similar observations in rabbit do have been reported in earlier studies (Patridge et al., 1986; Parigi-Bini et al., 1990). The results of this experiment suggest that pregnant grasscutters were more efficient in adjusting the balance of nutrients and body reserves on the 14% dietary protein level. Maternal malnutrition has been reported by Yahaya (1993) and Smith and Somade (1994) to compromise neonatal survival through its adverse effects on birth weight of rabbits.
The higher reproductive performance of gestating grasscutters on the CP 14 diet could be due to the higher level of total feed intake (including especially the higher intake of the 14% dietary protein). Results showed that the relationship between feed intake and reproductive performance followed the same trend as that observed between feed intake and daily weight gain or average terminal weights of gestating grasscutters. The role of fibre in determining the level of feed intake could also be important in the reproductive performance of gestating grasscutters. The results also indicate that higher than required (optimum) dietary protein levels did not increase reproductive performance. These findings suggest that gestating grasscutters adjust intake to meet dietary protein requirements for growth and maintenance of pregnancy. According to Lebas et al. (1996) rabbits adjust their feed intake according to the concentration of energy and other nutrients in their feed in order to satisfy demands. The average daily total feed intake (127.42 to 144.12 g) obtained in this study is lower than the range of 150 to 250 g reported by Mensah (1995) for all physiological classes of grasscutter.
The amount of feed consumed by gestating grasscutters per unit weight gain (or feed conversion ratio) was least (9.33) on the CP 22 diet, followed by that on the CP 14 diet. Feed conversion ratio tended to be better with increase in the dietary protein levels above 14% CP. This could be due to increased caecal microbial fermentation in the presence of higher dietary protein and lower dietary fibre. The cost of diet per unit weight gain was lowest on the 22% CP diet, followed by that on the 14% CP diet. The cost to gain ratio tended to decrease with increase in the dietary protein levels. The higher dietary protein levels were, therefore, more consistent with better feed utilization and cost effectiveness of diets.
CONCLUSION
The growth and reproductive performance of gestating grasscutters was best on the 14% CP diet. These findings suggest that the optimum dietary crude protein level for raising gestating grasscutters is 14%, which level of crude protein could be supplied from the locally available and low-cost agro-industrial by-products used in this study.
REFERENCES
- Abulude, F.O., 2007. Determination of the chemical composition of bush meats found in Nigeria. Am. J. Food Technol., 2: 153-160.
CrossRefDirect Link - Adeniji, A.A., 2008. Effects of feeding three protein sources with or without fishmeal supplementation on the performance of growing grasscutters. Asian J. Anim. Vet. Adv., 3: 98-103.
CrossRefDirect Link - Addo, P., 2002. Detection of mating, pregnancy and imminent Parturition in the grasscutter (Thryonomys swinderianus) Livest. Res. Rural Dev., 14: 8-13.
Direct Link - Asibey, E.O.A., 1974. Wildlife as a source of protein in Africa South of the Sahara. Biol. Conservation, 6: 32-39.
CrossRef - Bawa, G.S., S.O. Ajide, I.A. Adeyinka and M.K. Ajala, 2008. Effect of varying levels of groundnut haulms and cowpea shells on the performance weaned rabbits. Asian J. Anim. Vet. Adv., 3: 54-61.
CrossRefDirect Link - Chukwuka, O.K., I.C. Okoli, N.J. Okeudo, M.N. Opara, U. Herbert, I.P. Ogbuewu and B.U. Ekenyem, 2010. Reproductive potentials of west african dwarf sheep and goat: A review. Res. J. Vet. Sci., 3: 86-100.
CrossRefDirect Link - De Wilde, R.O., 1980. Protein and energy retentions in pregnant and non-pregnant gilts. 2. Energy retention. Livestock Prod. Sci., 7: 505-510.
CrossRef - El-Hassan, K., El-Abid and A.M.A. Abu Nikhaila, 2009. A study on some non-genetic factors and their impact on some reproductive traits of sudanese nubian goats. Int. J. Dairy Sci., 4: 152-158.
CrossRefDirect Link - Azahan, E.A.E. And M. Noraziah, 2011. Evaluation of dietary protein intake by growing ostriches. Asian J. Poult. Sci., 5: 102-106.
CrossRefDirect Link - Lameed, G.A. and O.O. Ogundijo, 2006. Effect of varied dietary protein levels on the reproductive performance of grasscutter (Thryonomys swinderianus) in captivity. J. Anim. Vet. Adv., 5: 361-363.
Direct Link - Meduna, A.J., 2002. Preliminary observations on cane rat (Thryonomys swinderianus, Termminck) feeding and breeding of the Federal College of forestry, Ibadan. Proceeding of the 27th Annual Conference on Nigerian Society for Animal Production, March 17-21, 2002, Federal University of Technology, Akure, pp: 304-305.
- Mensah, G.A., 2000. General presentation on the breeding of grasscutters, history and distribution in Africa. Proceedings of the International Seminar Proceedings on Intensive Breeding of Wildlife Animals for the Purpose of Food in Liberville, May 23-24, 2000, Gabon Project, DGEG/VSF/ADIE/CARPE/UE., pp: 45-59.
- Mensah, G.A. and A.M. Okeyo, 2005. Continued harvest of the diverse African animal genetic resources from the wild through domestication as a strategy for sustainable use: A case of the larger grasscutter (Thryonomis swinderianus). International Livestock Research Institute, http://agtr.ilri.cgiar.org/agtrweb/index.php?option=com_content&task=view&id=177&Itemid=199.
- NRC, 1991. Microlivestock: Little-Known Small Animals with a Promising Economic Future. 1st Ed. National Academy of Sciences, Washington, DC., pp: 241-249.
Direct Link - Omole, A.J., O. Ayodeji, O.A. Ashaye and A.K. Tiamiyu, 2005. Effect of scalding and flaming methods of processing on physiochemical and organoleptic properties of grasscutter meat. J. Applied Sci. Res., 1: 249-252.
Direct Link - Opara, M.N., 2010. The grasscutter I: A livestock of tomorrow. Res. J. For., 4: 119-135.
CrossRefDirect Link - Opara, M.N., 2010. Grasscutter: The haematology and major parasites. Res. J. Parasitol., 5: 214-223.
CrossRefDirect Link - Parigi-Bini, R., G. Xiccato and M. Cinetto, 1990. Energy and protein retention and partition in pregnant and non-pregnant rabbit does during the first pregnancy. Cuni-Science, 6: 19-29.
Direct Link - Patridge, G.G., Y. Daniels and R.A. Fordyce, 1986. The effects of energy intake during pregnancy in doe rabbits on pup birth weight, milk output and maternal body composition change in the ensuing lactation. J. Agric. Sci., 107: 697-708.
CrossRef - Ren, K., Y. Li, Q. Liang, J. Ren, H. Luo, Y. Tang and S. Huang, 2005. Digestible energy and crude protein level for growing rex rabbits. Proceedings of the 8th World Rabbit Congress, September 7-10, 2005, Pueblo, Mexico, pp: 968-972.
Direct Link - Xiccato, G., M. Bernardini, C. Castellini, A. Dalle Zotte, P.I. Queaque and A. Trocino, 1999. Effect of post-weaning feeding on the performance and energy balance of female rabbits of different physiology states. J. Anim. Sci., 77: 416-426.
Direct Link - Yi, S.S.A. and O.O. Tewe, 1980. Food preference and carcass composition of the grasscutter (Thryonomys swinderianus) in captivity. Afr. J. Ecol., 18: 133-140.
CrossRef