
Claire d`Andre Hirwa
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Paul Wallace
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Xu Shen
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Qinghua Nie
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Guanfu Yang
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Xiquan Zhang
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Asian Journal of Animal Sciences
Year: 2011 | Volume: 5 | Issue: 1 | Page No.: 34-45







ABSTRACT
The candidate genes with important functions in animal metabolism can help to identify potential molecular markers for cattle production traits. The study of the genes related to economically traits is one important aspect of the breeding. This study reviews the genes related to economically important traits in beef cattle such as Peroxisome proliferatior-activated receptor-γ (PPAR-γ), Sterol regulatory element-binding protein 1(SREBP-1), leptin, stearoyl-CoA desaturase (SCD1), CCAAT-enhancer-binding proteins (or C/EBBS), calpastatin, thyroid hormone-responsive protein (THRSP), corticotrophin releasing hormone (CRH), fatty acid binding protein 4 (FABP4), DGAT1, FASN,… genes. In conclusion, this study will help in genetic improvement program in cattle.
PDF Abstract XML References Citation
Received: February 17, 2010;
Accepted: April 09, 2010;
Published: July 27, 2010
How to cite this article
Claire d`Andre Hirwa, Paul Wallace, Xu Shen, Qinghua Nie, Guanfu Yang and Xiquan Zhang, 2011. Genes Related to Economically Important Traits in Beef Cattle. Asian Journal of Animal Sciences, 5: 34-45.
DOI: 10.3923/ajas.2011.34.45
URL: https://scialert.net/abstract/?doi=ajas.2011.34.45
DOI: 10.3923/ajas.2011.34.45
URL: https://scialert.net/abstract/?doi=ajas.2011.34.45
INTRODUCTION
In breeding for beef, adipose tissue types and amounts are reported to be directly associated with quality and value of the meat. The use of molecular techniques has provided substantial knowledge regarding molecular markers paralleling the processes of adipogenesis and lipid metabolism, both of which coordinate the overall fatness of an animal. Adipogenesis is defined as the proliferation, differentiation and initial incorporation of lipids in cells of adipose lineage to form lipid-assimilating adipocytes. Adipogenesis within specific adipose depots in ruminants may be under different regulation as against that experienced by 3T3-L1 cells. Marbling (intramuscular fat) is the main factor in determining important parameters that contribute to meat quality such as juiciness and flavor. It is also an essential factor in determining the economic value of beef. Marbling of cattle meat is dependent on the coordinated expression of multiple genes. Cattle dramatically increase their intramuscular fat content in the longissimus dorsi muscle between 12 and 27 months of age according to Lee et al. (2005).
However, it can still increase as a result of other fat depots such as subcutaneous fat and visceral fat, which lead to inefficient meat production. As such genomics studies have been performed to identify marbling specific genes for application in animal breeding programs. Fat development in muscles begin at the early stage of animal development and the intramuscular fat content at these early stage is an essential factor in determining the final level of marbling in the adult animal. Marbling is the fat within muscle and it is a valued trait which improves the eating quality of beef. It is also highly sought after by many exporters of meat.
Though beef is the number one protein source and the demand continues to grow, it also contains high amounts of saturated fat as a result of bio-hydrogenation in the rumen (De Smet et al., 2004). Marbling depots in cattle is important because industrially it is the primary determinant of price or value and scientifically easy to access. As a result of the fact that the growth of intramuscular fat appears to be driven by excess glucose in the diet (Pethick et al., 2004), it may be of relevance in human studies. For many years, fats in meat have received considerable attention because they do not only lead to obesity, but also increase the risk of a number of health problems such as heart disease, stroke, diabetes, obesity and in some cases cancers. Molecular genetics have led to the identification of DNA polymorphisms that affect traits of interest in animal breeding (Abe et al., 2008; Nie et al., 2004, 2005a, b; Bionaz and Loor Jaun, 2008), the identification of genes that contribute to variation in meat quality, including intramuscular fat level, remains a challenge.
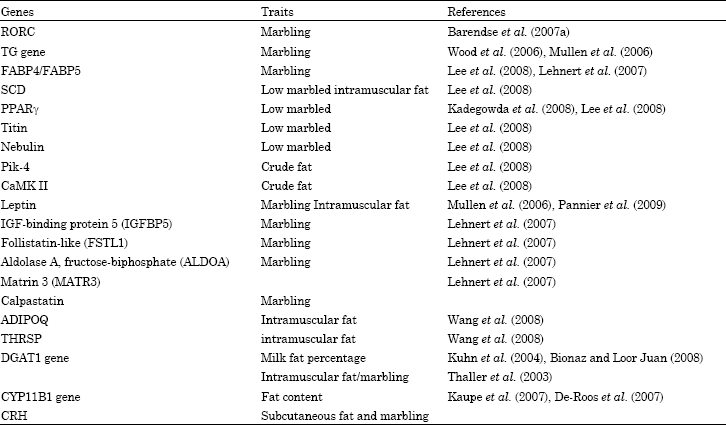
In our recent review, we presented different genes related to fat deposition and mammary lipid metabolism in cattle such as PPAR-γ, SREBP-1, leptin, stearoyl-CoA desaturase (SCD1), CCAAT-enhancer-binding proteins (or C/EBBS), calpastatin, thyroid hormone-responsive protein (THRSP), corticotrophin releasing hormone (CRH), fatty acid binding protein 4 (FABP4), DGAT1 and FASN genes (Taniguchi et al., 2007). The following highlights the influence of some of the genes that contributes to economically important traits in beef cattle (Table 1).
Peroxisome Proliferatior-activated Receptor-γ PPARγ): Peroxisome proliferatior-activated receptor-γ (PPARγ) is a transcription factor that modulates lipid and glucose metabolism in mammals. This is a member of the nuclear receptor super family. PPARγ is expressed in many cell types including adipocytes, epithelial cells, endothelial cells smooth muscle cells as well as many tissues (Spiegelman, 1997; Clark et al., 2000; Law et al., 2000).
| Table 1: | Overview of genes related to fat deposition in cattle |
 | |
The fat content in cattle may therefore be regulated by the PPARy transcription factor in a depot-specific manner.
Considerable progress has been made during the past years in understanding the molecular mechanisms of adipogenesis. Three classes of transcription factors that directly influence adipogenesis have so far been identified. These are the peroxisome proliferator activated receptor (PPAR), CCAAT/enhancer binding proteins (C/EBBS) and sterol regulatory element binding protein 1c (SREBP-1c, also called ADD1, adipocyte determination and differentiation factor). PPARγ is therefore an important transcription factor during adipogenesis. In addition, Rosen et al. (1999) reported from their studies of mice that were chimeric for wild-type and PPARγ null cells, that PPARγ null cells showed little or no contribution to adipose tissue, whereas most of the other organs examined did not require PPARγ for the proper development.
The role of Peroxisome Proliferator-activated Receptors (PPARs) in fat accumulation: On the whole, research into adipogenesis has focused on a few important regulators involved in the control of lipid metabolic genes namely PPARy, SREBP1/ADD1 and C/EBBS (Rosen et al., 1999). Nonetheless, researchers continue to characterize new regulators of adipogenesis on a regular basis. For example, recent studies with 01 f-1/early B-cell factor (O/E-l) has indicated the expression in pre-adipocyte cell lines that adipogenesis and expression in non-committed fibroblasts lead to the initiation of adipocyte differentiation (Akerblad et al., 2002). Peroxisome proliferators regulate gene transcription by activating a class of nuclear receptors, the peroxisome proliferator-activated receptors (PPAR), which are related to receptors for thyroid and steroid hormones (Rosen et al., 1999) and are members of the nuclear hormone receptor super family (Escher and Wahli, 2000). Synthetic PPAR ligands include peroxisome proliferators, hypolipidemic, anti-inflammatory and insulin sensitizing compounds whereas natural PPAR ligands include medium- and long-chain fatty acids and eicosanoids (Escher and Wahli, 2000). Three PPAR isoforms designated a, p and y has been characterized in a wide variety of species, with each isoform encoded by a separate gene and demonstrating ligand-selectivity (Grimaldi, 2001). For example, Belury et al. (2003) recently demonstrated that Conjugated Linoleic Acid (CLA) is an activator and ligand for both PPARcc and PPARγ. PPARγ is highly expressed in adipocytes and plays a crucial role in adipocyte differentiation (Tontonoz et al., 1994). PPARγ directly controls the expression of many genes that define the adipocyte phenotype and its expression is essential for the development of adipose tissue in vivo.
CLAs are a mixture of positional and geometric isomers of octadecadienoic acid (CI 8:2). Of the nine known isomers, the cis-9, trans-l 1 form, also known as rumenic acid, is the most common natural form with biological activity (Moloney et al., 2001). CLA has been shown to have anti-carcinogenic, anti-atherogenic, lean-body mass enhancing and anti-diabetic properties (McGuire and McGuire, 2000). The highest concentrations of CLA are found in the fat from ruminants where it is produced in the rumen from linoleic acid (Bauman et al., 1998). Ruminants synthesize rumenic acid from trans-l 1 octadecenoic acid via the enzymatic action of A9-desaturase which is normally present in mammary tissue and adipose tissue (Bauman et al., 1998).
Sterol Regulatory Element-Binding Protein 1(SREBP-1): Sterol regulatory element-binding protein 1 is a transcriptional factor that plays a pivotal role in energy homeostasis by promoting glycolysis, lipogenesis and adipogenesis. SREBP1 is pivotal in the regulation of milk fat synthesis in mouse and cow. SREBP-1c/ADD1 displays a dual DNA specificity, binding to both an E-box motif and non-E-box motif (SRE-1). A role in adipocyte differentiation is suggested by the early increase of its mRNA levels during adipocyte differentiation and by its transactivation of several key lipogenic genes (Bennett et al., 2008). SREBP-1c/ADD1 has been shown to increase fatty acid and fat synthesis and this has been attributed in part to its proposed influence on PPAR-γ activity; it has been hypothesized to be involved in the generation of endogenous ligands for PPAR-γ SREBP1 and 2 reside as inactive precursors in the ER membrane and are transported to the Golgi for proteolytic cleavage (i.e., activation) prior to entering the nucleus and activation of sterol responsive element-containing genes (e.g., ACACA, FASN). The transport step to the Golgi is blocked by sterols via the sterol-sensing protein SCAP (SREBP cleavage activating protein). Among the three major isoforms, SREBP-1a is a potent transcriptional activator of the LDLR gene and certain genes that encode enzymes in the biosynthesis of cholesterol and unsaturated fatty acids. SREBP-1c selectively stimulates fatty acid synthase while SREBP-2 drives cholesterol biosynthetic genes. Recent studies have demonstrated that the interaction of SREBP with SRE in human promoter enhances Sp1 binding to R3 and also promote transcription synergistically.
Insulin induced gene (INSIG) 1 and 2 are proteins that interact with SCAP in an oxysterol-dependent and independent fashion and regulate the responsiveness of SREBP1 and 2 processing via SCAP, thus altering rates of lipogenesis.
Leptin gene: It is a hormone that is mainly synthesized in the adipose tissue at a rate strongly related to adiposity in ruminants. Leptin is the hormonal product of the obese gene synthesized and secreted predominantly by white adipocytes (Zhang et al., 1994; Ji et al., 1998). Leptin (from the Greek leptos, meaning thin) is a protein hormone with important effects in regulating body weight, metabolism and reproductive function. The protein is approximately 16 kDa in mass and encoded by the obese (ob) gene. Leptin is expressed predominantly by adipocytes, which fits into the idea that body weight is sensed as the total mass of fat in the body. Smaller amounts of leptin are also secreted by cells in the epithelium of the stomach and placenta. Its receptors are highly expressed in areas of the hypothalamus known to be important in regulating body weight, as well as in T lymphocytes and vascular endothelial cells (Schenkel et al., 2006a).
The role of leptin as a lipostatic signal regulating whole body energy metabolism makes it one of the best physiological markers of body weight, food intake and energy expenditure (Houseknecht et al., 1998; Woods et al., 1998; Schenkel et al., 2005), reproduction (Cunningham et al., 1999) and certain immune functions (Lord et al., 1998). Circulating leptin and adipose tissue leptin mRNA levels are correlated with body weight, food intake, nutritional status as well as adipose tissue mass in humans and animals (Larsson et al., 1998; Delavaud et al., 2002).
Control of leptin synthesis and secretion: The amount of leptin expressed by adipocytes correlate well with the lipid content of cells. The mechanisms responsible for regulating leptin expression in adipocytes are unknown. It is likely that a number of hormones modulate ob gene expression including glucocorticoids and insulin (http://www.en.wikipedia.org/wiki/ Leptin#Discovery). Therefore, once leptin has bound to the Ob-Rb receptor, it activates the stat 3, which is phosphorylated and travels to the nucleus to, presumably, effect changes in gene expression. One of the main effects on gene expression is the down-regulation of the expression of endocannabinoids which is responsible for increasing appetite.
In cattle, the leptin gene has been mapped to bovine chromosome 4 according to Stone et al. (1996). Polymorphisms in the coding regions of the leptin gene in cattle have been associated with body fatness (Buchanan et al., 2002; Nkrumah et al., 2004, 2006; Pannier et al., 2009).
Stearoyl-CoA Desaturase (SCD1): Stearoyl-CoA desaturase (SCD1) activity is associated with increased fat accumulation and mono-unsaturation of saturated fatty acids in skeletal muscle. It is a rate-limiting enzyme that catalyzes the synthesis of mono-unsaturated fatty acids, which are components of triglycerides, wax esters, cholesteryl esters and membrane phospholipids (Miyazaki and Ntambi, 2003). The gene is highly expressed in white adipose tissue as well as brown adipose tissue under normal dietary conditions (Jiang et al., 2008).
The recent study provided a relationship between SCD gene polymorphism and SCD activity and fatty acid composition in Italian Holstein cattle breed (Conte et al., 2006). The same SCD gene polymorphism has been related to fatty acid composition of beef in the Japanese Black cattle breed, highly appreciated for its marbled beef and low fat melting point (Taniguchi et al., 2004). SCD gene could particularly interesting for the improvement of fat quality of ruminant-derived food products and for its implications on human health. The mammalian SCD gene, isolated in several species including mouse, cattle and human, shows a conserved genomic structure, spanning approximately 15-24 kb and consisting of six exons and five introns (Milanesi et al., 2007).
CCAAT-enhancer-binding proteins (or C/EBBS): C/EBBS are a family of transcription factors that are composed of six members i.e., C/EBP α to C/EBP. They promote the expression of certain genes through interaction with their promoters. Once bound to DNA, C/EBBS can recruit so-called co-activators (such as CBP) which in turn, can open up chromatin structure, or recruit basal transcription factors. C/EBP proteins interact with the CCAAT (cytidine-cytidine-adenosine-adenosine-thymidine) box motif which is present in several gene promoters. They are characterized by a highly conserved basic-leucine zipper (bZIP) domain at the C-terminus. This domain is involved in dimerization and DNA binding like other transcription factors of the leucine zipper family (e.g., c-Fos and Jun). C/EBBS bZIP domain structure is composed of α-helix that forms a coiled structure when it dimerizes. Studies of adipocyte differentiation, primarily in 3T3-L1 cells, provide extensive evidence for C/EPB-α function in adipocyte differentiation. Although not strictly adipocyte specific, C/EPB-α is expressed just before the transcription of most adipocyte-specific genes is initiated. CCAAT/enhancer binding protein-α binds and Trans-activate the promoters of several adipocyte genes including a P2, SCD1, GLUT-4, PEPCK, leptin and insulin receptor. Mutation of the C/EPB-α site in these genes abolishes transactivation. These proteins are found in hepatocytes, adipocytes, hematopoietic cells, spleen, kidney, brain and many others organs. C/EBBS proteins are involved in different cellular responses like in the control of cellular proliferation, growth and differentiation, metabolism, immunology and many others. All the members of the C/EBP family, except C/EBPγ, can induce transcription, through their activation domains and this is accoplished by interacting with components of the basal transcription apparatus. Their expression is regulated at multiple levels through hormones, mitogens, cytokines, nutrients, etc.
Intensive efforts dedicated to the isolation of regulatory genes for adipocyte differentiation have resulted in the identification of C/EBP and PPAR family of transcription factors, which play a central role in the terminal differentiation of adipocytes. In addition, the expression pattern of the above-mentioned helix-loop-helix transcription factors in the adipocyte lineage is consistent with the concept that other transcription factors may function in, or modulate, different stages in adipocyte development. The use of new cell culture models such as ES cells has been shown to differentiate at high efficiency into adipocytes in certain culture conditions. This may provide a system for the identification of new adipogenic regulatory genes or those that function at earlier stages.
Calpastatin: The protein encoded by this gene is an endogenous calpain (calcium-dependent cysteine protease) inhibitor. It consists of an N-terminal domain L and four repetitive calpain-inhibition domains (domains1-4) and it is involved in the proteolysis of amyloid precursor protein. The calpain/calpastatin system is involved in numerous membrane fusion events, such as neural vesicle exocytosis, platelet and red-cell aggregation. The encoded protein is also thought to affect the expression level of genes encoding structural or regulatory proteins. Calpastatin is a specific inhibitor of calpains (Goll et al., 2005) and is believed to be involved in the degradation of myofibrillar proteins in post-mortem proteolysis through regulating calpain activity (Koohmaraie, 1992) and it is implicated in meat tenderization. In beef cattle, two single nucleotide polymorphisms (SNPs) have been identified in the CAST gene, a G/C SNP in intron 5 (Schenkel et al., 2006b) and an A/G SNP in the 3' UTR region (Barendse, 1999; Barendse et al., 2007b). The association with meat tenderness (Casas et al., 2006; Morris et al., 2006; Schenkel et al., 2006b).
Thyroid Hormone-responsive Protein (THRSP): Thyroid hormone-responsive protein (THRSP) is a nuclear protein expressed in liver, fat and lactating mammary gland and functions in the regulation of fatty acid synthesis. The THRSP gene referred to as thyroid hormone responsive Spot 14 protein (THRSP), encodes a small acidic protein that was discovered in earlier studies of thyroid hormone action on hepatocytes (Joel et al., 1998; D’Andre Hirwa et al., 2009). The expression of this gene was determined in bovine mammary gland and mapped the THRSP gene to bovine chromosome 29 nearest microsatellite marker RM179 on the USDA linkage map. The full sequence of the bovine THRSP mRNA was determined by sequencing a clone from a normalized bovine mammary gland cDNA library (Accession No. AY656814). THRSP was highly correlated with intramuscular fat content (Wang et al., 2008). THRSP plays a role in intramuscular adipose tissue differentiation during the early portion of the growing phase (Graugnard et al., 2009).
Corticotrophin Releasing Hormone (CRH) gene: CRH is a stress hormone, released to the anterior pituitary to stimulate the secretion of adrenocorticotrophic hormone (ACTH), which upregulates cortisol. Cortisol has many significant metabolic effects that include stimulating gluconeogenesis in the liver, inhibiting glucose uptake in muscle and adipose tissue as well as stimulating fat breakdown in adipose tissue. In addition, transgenic mice with overexpression of CRH exhibit muscle wasting, decreased linear growth and obesity (Stenzel-Poore et al., 1992) whereas porcine CRH was reported to be significantly associated with back fat thickness, carcass length, average daily mass gain and REA. Thus, CRH is a good positional candidate gene for fat-related traits. Buchanan et al. (2005) showed three SNPs that were associated with REA (p<0.034) and hot carcass mass (p<0.0015) in a Charolais-cross steer population. In 2007, Wibowo and others discovered a new SNP in the bovine CRH gene that was thought to be highly associated with back fat thickness in a Wagyu X Limousin F2 population (p<0.001).
Fatty Acid Binding Protein 4 (FABP4) genes: These genes showed more than two-fold expression in the muscle of newborn Wagyu-sired calves while FABP5, indicated a higher level of adipogenic differentiation in the longissimus muscle of these animals, even at this early stage of their development. FABP4 showed a similar pattern of breed-specific differences and the differential expression of both genes was confirmed by qRT-PCR (Lehnert et al., 2007). Fatty acid binding proteins are expressed at very high levels by adipocytes and are themselves, by interaction with peroxisome proliferator-activated receptors (PPAR) involved in mediating the effects of fatty acids on gene regulation (Tan et al., 2002). Genetic variation in the bovine FABP4 gene has been found to be associated with intramuscular fat content and subcutaneous fat depth (Michal et al., 2006). Recent study reported that FABP4 genes is associated with intramuscular fat (IMF) levels or marbling scores in beef (Pannier et al., 2010).
DGAT1 gene: DGAT1 is a microsomal enzyme that utilizes diacylglycerol and fatty acyl CoA as substrates in order to catalyze the final stage of triacylglycerol synthesis. Up-regulation of DGAT1 gene was validated to be co-expressed with PPAR-γ and FABPs (Taniguchi et al., 2007). The studies revealed that DGAT1 gene was significantly associated with fat related traits such as backfat thickness and marbling in beef and fat content and composition in dairy cattle (Moore et al., 2003; Thaller et al., 2003; Schennink et al., 2007). These indicate that up-regulation of DGAT1 gene may have certain functional effect upon the adipocyte development in beef cattle. This gene affect fat metabolism, including fat yield and percentage in the milk. A knock-out study showed that both male and female Dgat mice, even those fed a high fat diet, stabilized their weights and resisted fat storage which indicated the importance of DGAT1 in fat metabolism. Recent studies reported that DGAT1genes is associated with intramuscular fat levels or marbling scores in beef (Pannier et al., 2010). Intramuscular fat represents an important beef quality trait as it contributes to the juiciness and flavour of cooked meat (Shahidi, 2002; Thompson, 2004). DGAT1 and FABP4 genes were identified to be genetic factors influencing intramuscular fat deposition in muscle (Barendse, 1999; Michal et al., 2006; Thaller et al., 2003).
CONCLUSION
Advances in molecular genetics have led to the identification of genes or markers associated with genes that affect economically important traits in both dairy and beef cattle. The molecular basis of meat quality being unearthed revealed by functional genomics approaches. This will help us to gain further insight into the biological components and the development of meat quality as well as milk production. It gives greater opportunities to enhance genetic improvement program in cattle through marker-assisted selection.
Most of the traits considered in animal genetic improvement programs are quantitative traits; i.e., they are controlled by many genes, together with environmental factors while the underlying genes have minimal effects on the observable phenotype. Milk yield and growth rate in animals or yield and size in plants are typical examples of quantitative traits. In classical genetic improvement programs, selection is carried out based on observable phenotypes (of the candidates for selection and/or their relatives) but without knowing which genes are actually being selected. The development of molecular markers was therefore greeted with great enthusiasm as it was seen as a major breakthrough promising to overcome this key limitation.
REFERENCES
- Abe, T., J. Saburi, H. Hasebe, T. Nakagawa and T. Kawamura et al., 2008. Bovine QTL analysis for growth, carcass and meat quality traits in an F2 population from a cross between Japanese Black and Limousin. J. Anim. Sci., 86: 2821-2832.
CrossRefPubMedDirect Link - Akerblad, P., U. Lind, D. Liberg, K. Bamberg and M. Sigvardsson, 2002. Early B-cell factor (O/E-1) is a promoter of adipogenesis and involved in control of genes important for terminal adipocyte differentiation. Mol. Cell. Biol., 22: 8015-8025.
PubMed - Barendse, W., Bunch, R.J. J.W. Kijas and M.B. Thomas, 2007. The effect of genetic variation of the retinoic acid receptor-related orphan receptor C gene on fatness in cattle. Genetics, 175: 843-853.
CrossRef - Barendse, W., B.E. Harrison, R.J. Hawken, D.M. Ferguson, J.M. Thompson, M.B. Thomas and R.J. Bunch, 2007. Epistasis between calpain 1 and its inhibitor calpastatin within breeds of cattle. Genetics, 176: 2601-2610.
CrossRef - Belury, M.A., A. Mahon and S. Banni, 2003. The conjugated linoleic acid (CLA) isomer, t10c12-CLA, is inversely associated with changes in body weight and serum leptin in subjects with type 2 diabetes mellitus. J. Nutr., 133: 257S-260S.
PubMed - Bionaz, M. and J. Loor Juan, 2008. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genomics, 9: 366-366.
Direct Link - Buchanan, F.C., C.J. Fitzsimmons, A.G. van Kessel, T.D. Thue, D.C.W. Sim and S.M. Schmutz, 2002. Association of a missense mutation in the bovine leptin gene with carcass fat content and leptin mRNA levels. Genet. Sel. Evol., 34: 105-116.
CrossRefPubMedDirect Link - Buchanan, F.C., T.D. Thue, P. Yu and D.C. Winkelman-Sim, 2005. Single nucleotide polymorphisms in the corticotrophin-releasing hormone and pro-opiomelancortin genes are associated with growth and carcass yield in beef cattle. Anim. Genet., 36: 127-131.
PubMed - Casas, E., S.N. White, T.L. Wheeler, S.D. Shackelford and M. Koohmaraie et al., 2006. Effects of calpastatin and micro-calpain markers in beef cattle on tenderness traits. J. Anim. Sci., 84: 520-525.
PubMedDirect Link - Clark, R.B., D. Bishop-Bailey, T. Estrada-Hernandez, T. Hla, L. Puddington and S.J. Padula, 2000. The nuclear receptor PPAR and immunoregulation: PPAR mediates inhibition of helper T cell responses. J. Immunol., 164: 1364-1371.
PubMed - Conte, G., M. Mele, B. Castiglioni, A. Serra and M. Viva et al., 2006. Relationship between bovine SCD polymorphism locus and mammary gland desaturation activity. Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Aug. 13-18, Belo Horizonte, Minas Gerais, Brazil, pp: 22-37.
- Cunningham, M.J., D.K. Clifton and R. A. Steuner, 1999. Leptin`s actions on the reproductive axis: Perspective and mechanisms. Biol. Reprod., 60: 216-222.
Direct Link - De-Roos, A.P.W., C. Schrooten, E. Mullaart, M.P.L. Calus and R.F. Veerkamp, 2007. Breeding value estimation for fat percentage using dense markers on Bos taurus autosome 14. J. Dairy Sci., 90: 4821-4829.
CrossRef - De Smet, S., K. Raes and D. Demeyer, 2004. Meat fatty acid composition as affected by fatness and genetic factors: A review. Anim. Res., 53: 81-98.
CrossRefDirect Link - Delavaud, C., A. Ferlay, Y. Faulconnier, F. Bocquier, G. Kann and Y. Chilliard, 2002. Plasma leptin concentration in adult cattle: Effects of breed, adiposity, feeding level and meal intake. J. Anim. Sci., 80: 1317-1328.
Direct Link - Escher, P. and W. Wahli, 2000. Peroxisome proliferator-activated receptors: insights into multiple cellular functions. Mutat. Res., 448: 121-123.
CrossRef - Goll, D.E., V.F. Thompson, H. Li, W. Wei and J. Cong, 2003. The calpain system. Physiol. Rev., 83: 731-801.
PubMed - Graugnard, D.E., P. Piantoni, M. Bionaz, L.L. Berger, D.B. Faulkner and J.J. Loor, 2009. Adipogenic and energy metabolism gene networks in longissimus lumborum during rapid post-weaning growth in Angus and Angus x Simmental cattle fed high- or low-starch diets. BMC Genomics, 10: 142-142.
Direct Link - Grimaldi, P.A., 2001. The roles of PPARs in adipocyte differentiation. Progr. Lipid Res., 40: 269-281.
CrossRef - D'Andre Hirwa, C., W. Paul, W. Yan, C. Luo, Q. Nie, G. Yang and X. Zhang, 2009. Allelic frequency in chicken thyroid hormone responsive spot 14 alfa gene (THRSPα). Asian J. Anim. Sciences, 3: 85-91.
CrossRefDirect Link - Houseknecht, K.L., C.A. Baile, R.L. Matteri and M.E. Spurlock, 1998. The biology of leptin: A review. J. Anim. Sci., 76: 1405-1420.
PubMedDirect Link - Ji, S., G.M. Willis, R.R. Scott and M.E. Spurlock, 1998. Partial cloning and expression of the bovine leptin gene. Anim. Biotechnol., 9: 1-14.
PubMed - Jiang, Z., J.J. Michal, D.J. Tobey, T.F. Daniels, D.C. Rule and M.D. MacNeil, 2008. Significant associations of stearoyl-CoA desaturase (SCD1) gene with fat deposition and composition in skeletal muscle. Int. J. Biol. Sci., 4: 345-351.
Direct Link - Kaupe, B., H. Brandt, E.M. Prinzenberg and G. Erhardt, 2007. Joint analysis of the influence of CYP11B1 and DGAT1 genetic variation on milk production, somatic cell score, conformation, reproduction and productive lifespan in German Holstein cattle. J. Anim. Sci., 85: 11-21.
CrossRefDirect Link - Koohmaraie, M., 1992. The role of Ca(2+)-dependent proteases (calpains) in post mortem proteolysis and meat tenderness. Biochimie, 74: 239-245.
PubMed - Kuhn, C., G. Thaller, A. Winter, O.R.P. Bininda-Emonds and B. Kaupe et al., 2004. Evidence for multiple alleles at the DGAT1 locus better explains a quantitative trait locus with major effect on milk fat content in cattle. Genetics, 167: 1873-1881.
CrossRefDirect Link - Larsson, H., S. Elmstahl, G. Berglund and B. Ahren, 1998. Evidence for leptin regulation of food intake in humans. J. Clin. Endocrinol. Metab., 83: 4382-4385.
Direct Link - Law, R.E., S. Goetze, X.P. Xi, S. Jackson and Y. Kawano et al., 2000. Expression and function of PPAR in rat and human vascular smooth muscle cells. Circulation, 101: 1311-1318.
Direct Link - Lee, S.H., Y.M. Cho, S.H. Lee, B.S. Kim and N.K. Kim et al., 2008. Identification of marbling-related candidate genes in M. longissimus dorsi of high and low marbled Hanwoo(Korean Native Cattle) steers. BMB Rep., 41: 846-851.
Direct Link - Lehnert, S.A., A.A. Reverter, K.A. Byrne, Y. Wang, G.S. Nattrass, N.J. Hudson and P.L. Greenwood, 2007. Gene expression studies of developing bovine longissimus muscle from 2 different beef cattle breeds. BMC Dev. Biol., 7: 1-13.
CrossRefDirect Link - McGuire, M.A. and M.K. McGuire, 2000. Conjugated Linoleic Acid (CLA): A ruminant fatty acid with beneficial effects on human health. J. Anim. Sci., 77: 1-8.
Direct Link - Michal, J.J., Z.W. Zhang, C.T. Gaskins and Z. Jiang, 2006. The bovine fatty acid binding protein 4 gene is significantly associated with marbling and subcutaneous fat depth in Wagyu x Limousin F2 crosses Anim. Genet., 37: 400-402.
PubMed - Miyazaki, M. and J.M. Ntambi, 2003. Role of stearoyl-coenzyme a desaturase in lipid metabolism. Prostaglandins Leukot Essent Fatty Acids, 68: 113-121.
CrossRefPubMedDirect Link - Moloney, A.P., M.T. Mooney, J.P. Kouryand D.J. Troy, 2001. Producing tender and flavor some beef with enhanced nutritional characteristics. Proc. Nutr. Soc., 60: 221-229.
PubMed - Moore, S.S., C. Li, J. Basarab, W.M. Snelling and J. Kneeland et al., 2003. Fine mapping of quantitative trait loci and assessment of positional candidate genes for backfat on bovine chromosome 14 in a commercial line of Bos tausus. J. Anim. Sci., 81: 1919-1925.
PubMed - Mullen, A.M., P.C. Stapleton, D. Corcoran, R.M. Hamill and A. White, 2006. Understanding meat quality through the application of genomic and proteomic approaches. Meat Sci., 74: 3-16.
CrossRef - Nie, Q., B. Sun, D. Zhang, C. Luo and N.A. Ishag et al., 2005. High diversity of the chicken growth hormone gene and effects on growth and carcass traits. J. Heredity, 96: 698-703.
CrossRefDirect Link - Nie, Q., H. Zeng, M. Lei, N.A. Ishag, M. Fang, B. Sun, G. Yang and X. Zhang, 2004. Genomic organisation of the chicken ghrelin gene and its single nucleotide polymorphisms detected by denaturing high-performance liquid chromatography. Br. Poult. Sci., 45: 611-618.
PubMed - Nkrumah, J.D., C. Li, J.A. Basarab, S. Guercio and Y. Meng et al., 2004. Association of a single nucleotide polymorphism in the bovine leptin gene with feed intake, growth, feed efficiency, feeding behaviour and carcass merit. Can. J. Anim. Sci., 84: 211-219.
Direct Link - Nkrumah, J.D., C. Li1, D.H. Keisler, E.L. Sherman and Z. Wang et al., 2006. Polymorphisms in the leptin gene and their associations with performance, feed efficiency, and carcass merit of beef cattle. Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Aug. 13-18, Belo Horizonte, MG, Brasil, pp: 1-4.
Direct Link - Pannier, L., T.Y. Sweene, R.M. Hamill, F. Ipek, P.C. Stapleton and A.M. Mullen, 2009. Lack of an association between single nucleotide polymorphisms in the bovine leptin gene and intramuscular fat in Bos taurus cattle. Meat Sci., 81: 731-737.
CrossRef - Pethick, D.W., G.S. Harper and V.H. Oddy, 2004. Growth, development and nutritional manipulation of marbling in cattle: A review. Aust. J. Exp. Agric., 44: 705-715.
CrossRef - Rosen, E.D., P. Sarraf, A.E. Troy, G. Bradwin and K. Moore et al., 1999. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell, 4: 611-617.
PubMed - Schenkel, F.S., S.P. Miller, Z. Jiang, I.B. Mandell, X. Ye, H. Li and J.W. Wilton, 2006. Association of a single nucleotide polymorphism in the calpastatin gene with carcass and meat quality traits of beef cattle. J. Anim. Sci., 84: 291-299.
CrossRefPubMedDirect Link - Schenkel, F.S., S.P. Miller, X. Ye, S.S. Moore and J.D. Nkrumah et al., 2005. Association of single nucleotide polymorphism in the leptin gene with carcass and meat quality traits of beef cattle. J. Anim. Sci., 83: 2009-2020.
PubMedDirect Link - Schenkel, F.S., S.P. Miller, S.S. Moore, C. Li and A. Fu et al., 2006. Association of SNPs in the leptin and leptin receptor genes with different fat depos in beef cattle. Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Aug. 13-18, Belo Horizonte, MG, Brasil, pp: 1-4.
Direct Link - Schennink, A., W.M. Stoop, M.H.P.W. Visker, J.M.L. Heck and H. Bovenhuis et al., 2007. DGAT1 underlies large genetic variation in milk-fat composition of dairy cows. Anim. Genet., 38: 467-473.
CrossRef - Spiegelman, B.M., 1997. Peroxisome proliferator-activated receptor gamma: A key regulator of adipogenesis and systemic insulin sensitivity. Eur. J. Med. Res., 28: 457-464.
PubMed - Stenzel-Poore, M.P., V.A. Cameron, J. Vaughan, P.E. Sachiko and W. Vale, 1992. Development Cushing`s Syndrome in corticotropin-releasing factor transgenic mice. Endocrinology, 130: 3378-3386.
Direct Link - Stone, R.T., S.M. Kappes and C.W. Beattie, 1996. The bovine homologue of the obese gene maps to chromosome 4. Mamm. Genome, 7: 399-400.
CrossRef - Tan, N.S., N.S. Shaw, N. Vinckenbosch, P. Liu and R. Yasmin et al., 2002. Selective cooperation between fatty acid binding proteins and peroxisome proliferator-activated receptors in regulating transcription. Mol. Cell Biol., 22: 5114-5127.
CrossRefDirect Link - Taniguchi, M., T. Utsugi, K. Oyama, H. Mannen and M. Kobayashi et al., 2004. Genotype of stearoyl-CoA desaturase is associated with fatty acid composition in Japanese Black cattle. Mamm. Genome, 14: 142-148.
CrossRefDirect Link - Thaller, G., C. Kuhn, A. Winter, G. Ewald and O. Bellmann et al., 2003. DGAT1, a new positional and functional candidate gene for intramuscular fat deposition in cattle. Anim. Genet., 34: 354-357.
CrossRefPubMedDirect Link - Thompson, J.M., 2004. The effects of marbling on flavour and juiciness scores of cooked beef, after adjusting to a constant tenderness. Aust. J. Exp. Agric., 44: 645-652.
CrossRef - Tontonoz, P., E. Hu and B.M. Spiegelman, 1994. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell, 79: 1147-1156.
PubMed - Wang, Y.H., N.I. Bower, A. Reverter, S.H. Tan and N. De Jager et al., 2008. Gene expression patterns during intramuscular fat development in cattle. J. Anim Sci.
CrossRefDirect Link - Wood, I.A., G. Moser, D.L. Burrell, K.L. Mengersen and D.J. Hetzel, 2006. A meta-analytic assessment of a thyroglobulin marker for marbling in beef cattle. Genet. Sel. Evol., 38: 479-494.
CrossRefDirect Link - Woods, S.C., R.J. Seeley, D. Jr. Porte and M.W. Schwartz, 1998. Signals that regulate food intake and energy homeostasis. Science, 280: 1378-1383.
CrossRefDirect Link - Zhang, Y., R. Proenca, M. Maffei, M. Barone, L. Leopold and J.M. Friedman, 1994. Positional cloning of the mouse obesity gene and its human homologue. Nature, 372: 425-432.
PubMed - Bennett, M.K., Y.K. Seo, S. Datta, D.J. Shin and T.F. Osborne, 2008. Selective binding of sterol regulatory element-binding protein isoforms and co-regulatory proteins to promoters for lipid metabolic genes in liver. J. Biol. Chem., 283: 15628-15637.
PubMed - Lord, G.M., G. Matarese, J.K. Howard, R.J. Baker, S.R. Bloom and R.I. Lechler, 1998. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature, 394: 897-901.
PubMed - Milanesi, E., L. Nicoloso and P. Crepaldi, 2007. Stearoyl CoA desaturase (SCD) gene polymorphisms in Italian cattle breeds. J. Anim. Breed. Genet., 125: 63-67.
PubMed