
G. Kapoor
Laboratory of Parasitology and Immunology, Department of Biosciences, Himachal Pradesh University, 171005, Shimla, India
H.S. Banyal
Laboratory of Parasitology and Immunology, Department of Biosciences, Himachal Pradesh University, 171005, Shimla, India
Asian Journal of Animal Sciences
Year: 2011 | Volume: 5 | Issue: 2 | Page No.: 145-152







ABSTRACT
Present study was undertaken to purify and characterize thioredoxin reductase (TrxR) of Plasmodium berghei, a rodent malaria parasite. Plasmodium contains thioredoxin redox system that acts as efficient antioxidant system preventing damage caused by enhanced oxidative stress. Thioredoxin (Trx) functions as redox messenger in the parasite maintaining a reduced intracellular environment. Thioredoxin reductase (E.C.1.8.1.9) was analysed in rodent malaria parasite, Plasmodium berghei. Cell-free parasite showed TrxR specific activity of 0.128±0.10 U mg-1. Maximum TrxR activity was observed in cytosolic fraction of P. berghei. The parasite enzyme was purified by ammonium sulphate precipitation and Sephadex G-200. Its molecular weight was 22 kDa and the enzyme remained maximally active at pH 7.4 while the higher temperatures inactivated the enzyme. Km (Michaelis constant) and Vmax (Maximum velocity of enzyme) values for dithionitrobenzene (DTNB) substrate were 1.25 and 0.1 mM, respectively. 1-chloro-2, 4-dinitro benzene (CDNB) uncompetitively inhibited the enzyme substrate reaction and the inhibition was concentration dependent. Ki (inhibition constant) for CDNB was found to be 1.25 mM for 0.01 mM CDNB and 1.3 mM for 0.1 mM CDNB.
PDF Abstract XML References Citation
How to cite this article
G. Kapoor and H.S. Banyal, 2011. Purification and Characterization of Plasmodium berghei Thioredoxin Reductase. Asian Journal of Animal Sciences, 5: 145-152.
DOI: 10.3923/ajas.2011.145.152
URL: https://scialert.net/abstract/?doi=ajas.2011.145.152
DOI: 10.3923/ajas.2011.145.152
URL: https://scialert.net/abstract/?doi=ajas.2011.145.152
INTRODUCTION
Plasmodium is exposed to oxidative stress during its intraerythrocytic development (Becker et al., 2004). Oxidative stress is generated by the production of Reactive Oxygen Species (ROS) produced by parasite metabolism, parasitized host red blood cells and by host’s immune responses. The malaria parasite is highly sensitive to oxidant stress and has its own battery of defense tactics lined up against ROS. The parasite protects itself against this oxidative stress by a number of host or parasite encoded antioxidant enzymes like superoxide dismutase, glutathione reductase and thioredoxin reductase, vitamin C, vitamin E etc.
Plasmodium contains functional thioredoxin redox system comprising thioredoxin (Trx), nicotinamide adenine di-nucleotide phosphate reduced (NADPH), thioredoxin reductase (TrxR) and thioredoxin dependent peroxidases. Thioredoxin (Trx) is a widely expressed protein contributing towards the essential cellular functions including protection from ROS (reactive oxygen species), reduction of enzymes like ribonucleotide reductase and thioredoxin peroxidases and regulation of transcription factors (Holmgren, 2000). Thioredoxin protects cytosolic protein from aggregation or inactivation via oxidative formation of intra or inter-molecular disulphides.
Thioredoxin reductase (TrxR; E.C. 1.8.1.9) catalyses the transfer of electrons from NADPH to thioredoxin. Plasmodium TrxR (PfTrxR) differs significantly from its mammalian counterpart as the former lacks selenium (Williams et al., 2000) and this enzyme is essential for the survival of P. falciparum erythrocytic stages (Krnajski et al., 2002).
In the dismal scenario of widespread resistance to various antimalarials there is an increasing pressure to explore novel drug targets to reduce the malaria burden. Enzymes of Plasmodium redox system are potential drug targets as their inhibition affects several vulnerable points in redox mechanism required for dealing with oxidative challenges in the host RBC. The 3D crystal structure of Plasmodium falciparum is not yet available and not much information is available on characterization of P. falciparum TrxR protein sequence (Banerjee et al., 2009).
Present study was carried out to detect the activity of TrxR in normal and P. berghei-infected erythrocytes, their fractions and cell-free parasite. Localization of TrxR in subcellular fractions of P. berghei. TrxR protein has been purified from rodent malaria parasite and some of its characteristics have been investigated in the present study.
MATERIALS AND METHODS
The study was carried at the Department of Biosciences, Himachal Pradesh University, Shimla, India from August 2004 to August 2007. All the experiments were carried out according to the procedures authorized by the Institutional Animals Ethics Committee (IAEC) of the university (IAEC/Bio/19-2005).
Parasite: Plasmodium berghei (NK-65) was maintained in white Swiss mice, Mus musculus (Balb/c) and course of parasitaemia was monitored by Giemsa stained thin blood smears (Kapoor et al., 2008).
Cell-free parasite: Blood from normal or P. berghei-infected mice was collected in citrate saline by jugular vein incision after anaesthetizing the mice with diethyl ether. The blood was centrifuged at 1,000 g for 10 min at 4°C. The red cell pellet was suspended in equal volume of phosphate buffer saline (PBS, 0.01 M), pH 7.2 and loaded onto a CF-11 cellulose (Whatmann) column (1.5x21 cm) to remove the leukocytes (Kapoor and Banyal, 2009). The eluted leukocyte free erythrocytes were centrifuged at 1,000 g, for 10 min at 4°C. A part of the settled red cell pellet was used as total erythrocyte and to the remaining equal volume of saponin (0.2% w/v) in PBS, pH 7.2 was added, the suspension incubated for 30 min at 4°C with intermittent mixing and centrifuged at 15,000 g for 20 min at 4°C. Haemolysate was aspirated and erythrocyte membranes overlying the cell-free parasite isolated from parasitized blood while in case of normal blood, haemolysate and erythrocyte membranes were collected. Cell-free parasite obtained was washed thrice with PBS, pH 7.2 (Banyal and Fitch, 1982).
Enzyme extract was prepared by suspending total erythrocyte, haemolysate, erythrocyte membranes and cell-free parasite in appropriate volume of 0.01 M PBS, pH 7.2, homogenized in Potter-Elvehjem homogenizer at 4°C and centrifuged at 1,000 g for 10 min at 4°C.
Sub-cellular fractionation: Cell-free P. berghei was homogenized at 4°C in pre-chilled 0.25 M sucrose solution. Different fractions were obtained by subjecting the homogenate to differential centrifugation (Banyal et al., 1979).
Enzyme assay: Thioredoxin reductase activity was spectrophotometrically measured by the reduction of dithionitrobenzene (DTNB) in the presence of nicotinamide adenine di-nucleotide phosphate reduced (NADPH) (Holmgren and Bjorsnstedt, 1995). The reaction mixture contained 100 mM potassium phosphate buffer, pH 7.5, 2 mM EDTA, 3 mM DTNB, 0.2 mM NADPH and appropriate volume of enzyme extract. After initiating the reaction with NADPH, the increase in absorbance was monitored at 412 nm at room temperature. Protein was determined according to Lowry et al. (1951) and specific activity was calculated as units of enzyme per mg protein.
Ammonium sulphate fractionation: The cell-free parasite homogenate was subjected to precipitation with ammonium sulphate (between 0 and 100%). The precipitates of each cut were dissolved in minimum volume of 10 mM Tris-HCl buffer, pH 7.5 and dialyzed at 4°C in same buffer containing 1 mM EDTA with changes of buffer.
Gel filtration on Sephadex G-200: Sephadex G-200 (Sigma) was swollen in distilled water for 72 h at 4°C. A glass column (1.5x21 cm) was filled with Sephadex G-200 and flow rate was maintained at 12 mL h-1. The column was equilibrated with 10 mM Tris-HCl buffer, pH 7.5 until the final absorbance difference became zero at 280 nm. The final dialyzed sample was loaded onto the column and elutions of 1.0 mL volume were collected at 4°C. In each fraction enzyme activity and protein were determined.
Characterization of TrxR: Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE): Electrophoretic characterization of purified enzyme was done by SDS-PAGE according to method of Laemmli (1970) in a mini vertical slab gel apparatus (Genei, Banglore) as given by Banyal and Inselburg (1986) using 3% stacking and 10% separating gel. Gels were stained in silver nitrate (Merril et al., 1981).
Optimum pH for P. berghei TrxR activity was determined using 100 mM potassium phosphate buffer (pH 5.5 to 8.0).
Optimum temperature for TrxR activity was determined by incubating the enzyme alone for 10 min at different temperatures in the range of -4 to 100°C (boiling water bath). After incubation the sample was transferred onto ice bath to prevent further destruction of enzyme.
To study the effect of inhibition for 1-chloro-2, 4-dinitro benzene (CDNB) the purified enzyme was incubated with or without CDNB for 5 min at room temperature and then assayed.
RESULTS
P. berghei exhibited significant amount of thioredoxin reductase activity. P. berghei infected erythrocytes and haemolysate exhibited higher TrxR activity compared to normal erythrocytes and haemolysate (Table 1). No TrxR activity was observed in erythrocyte membranes.
The cytosolic fraction contained maximum TrxR activity, which was 5.9 times higher than the crude total parasite homogenate (Table 2). Other fractions showed very little or no TrxR activity.
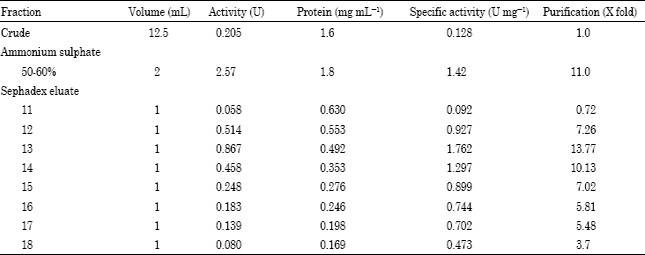
P. berghei TrxR was mainly precipitated in 50-60% ammonium sulphate cut with 11 fold purification of the enzyme. The elution profile of TrxR from Sephadex G-200 is shown in Fig. 1. The enzyme was eluted in fraction 11 to 18 with fraction 13 containing maximum TrxR activity (1.762 U mg-1). Sephadex G-200 resulted in about 54-fold purification of the parasite enzyme (Table 3).
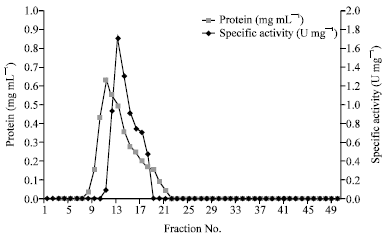
SDS-PAGE analysis of P. berghei TrxR resulted in a single band of 22 kDa (Fig. 2). The parasite enzyme was active at lower temperatures while boiling inactivated it (Table 4).
| Table 1: | Activity of thioredoxin reductase in normal and P. berghei infected erythrocytes and their fractions |
 | |
| Results are mean±SD of three experiments | |
| Table 2: | TrxR activity in subcellular fractions of P. berghei |
 | |
| Results are mean±SD of three experiments | |
| Table 3: | Purification of P. berghei TrxR by ammonium sulphate and Sephadex G-200 |
 | |
 | |
| Fig. 1: | Purification of thioredoxin reductase from cell-free P. berghei using Sephadex G-200 |
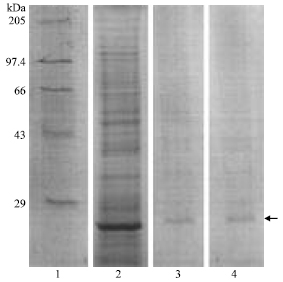
P. berghei TrxR was found to be maximally active at pH 7.4 and enzyme showed maximum activity of 0.155 U mg-1 (Fig. 3).
 | |
| Fig. 2: | SDS PAGE of TrxR purified from P. berghei; Lane 1: Protein standards; Lane 2: Cell free P. berghei homogenate; Lane 3: TrxR fraction 12; Lane 4: TrxR fraction 13 |
 | |
| Fig. 3: | Effect of pH on activity of TrxR purified from P. berghei |
| Table 4: | Effect of temperature on activity of P. berghei TrxR |
 | |
Km (Michaelis constant) and Vmax (maximum velocity of enzyme) for (dithionitrobenzene) DTNB were 1.25 and 0.1 mM, respectively (Fig. 4). CDNB (1-chloro2-4dinitrobenzene) competitively inhibited TrxR with Ki (inhibition constant) of 1.25 for 0.01 mM CDNB and 1.3 mM for 0.1 mM CDNB (Fig. 5).
 | |
| Fig. 4: | Effect of different concentration of DTNB on activity of P. berghei TrxR |
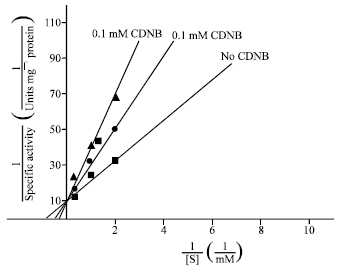 | |
| Fig. 5: | Ki determination of P. berghei TrxR using CDNB |
DISCUSSION
Significant amount of TrxR activity was observed in P. berghei-infected erythrocytes compared to normal mice erythrocytes while the erythrocyte membranes lack this enzyme. The increase in TrxR activity with P. bergehi infection indicates its role in oxidative stress. TrxR is known to be involved in maintenance of redox homeostasis and antioxidant defense in Plasmodium infection. The vital importance of thioredoxin redox cycle (comprising Trx and TrxR) is emphasized by the confirmation that TrxR knockout parasites are non viable (Krnajski et al., 2002).
Maximum activity of TrxR was observed in cytosolic fraction. Human TrxR and bovine TrxR are also reported to be cytosolic (Watabe et al., 1999). SDS-PAGE of purified TrxR revealed a subunit molecular mass of 22 kDa. TrxR exists in two different forms, low molecular weight Trxr-35 kDa found in prokaryotes, fungi, plants and protozoan parasites like Trichonomas vaginalis and high molecular weight TrxR as in mammals and P. falciparum having weight between 55 and 60 kDa (Williams et al., 2000; Coombs et al., 2004). The variation in enzyme molecular weight of rodent and human malaria parasite may be due to different species or monomer and dimmer nature of TrxR.
TrxR showed activity from -4 to 40°C while the activity declined with increase in temperature. Human TrxR becomes unstable at 40°C and is completely inactive at 60°C (Gromer et al., 1998). Parasite TrxR showed a wide range of pH activity with maximum at 7.4. Mitochondrial TrxR from bovine adrenal cortex exhibited a broad pH activity curve with peak at pH 7.5 (Watabe et al., 1999) while E. coli TrxR was maximally active at pH 7.7 (William, 1995). TrxR from rodent malaria parasite appears to be similar in characteristic like other TrxR.
Km (Michaelis constant) for di-thionitrobenzene (DTNB) of P. berghei TrxR is 1.25 mM and Vmax (Maximum velocity of enzyme) 0.1 mM. Km for DTNB of human placental TrxR is 0.4 mM (Oblong et al., 1993). CDNB competitively inhibited TrxR with Ki 1.25 mM (0.01 mM CDNB) and 1.3 mM (0.1 mM CDNB). CDNB is known to inhibit TrxR activity by covalent modification of selenoyl and thiol groups (Arner et al., 1995).
Plasmodium predominantly being intracellular utilizes minimum set of metabolic enzymes. Tremendous work has been done on enzymes of glycolytic, nucleotide and folate pathways. The antioxidant systems are important for the survival of the parasite during their erythrocytic cycle. Disruption of the antioxidant system of the parasite can be a feasible way to interfere with their development during erythrocytic schizogony. Enzymes of redox system are less studied in any species of malaria parasite and enzymes of these pathways like TrxR can be exploited as a new drug target.
ACKNOWLEDGMENT
Mr. Gaurav Kapoor is thankful to ICMR New Delhi for providing JRF-SRF.
REFERENCES
- Arner, E.S., M. Bjornstedt and A. Holmgren, 1995. 1-Chloro-2,4-dinitrobenzene is an irreversible inhibitor of human thioredoxin reductase: Loss of thioredoxin disulfide reductase activity is accompanied by a large increase in NADPH oxidase activity. J. Biol. Chem., 270: 3479-3482.
Direct Link - Banyal, H.S. and C.D. Fitch, 1982. Ferriprotoporphyrin IX binding substances and the mode of action of chloroquine against malaria. Life Sci., 31: 1141-1144.
PubMed - Banyal, H.S. and J. Inselburg, 1986. Plasmodium falciparum: Induction, selection and characterization of pyrimethamine-resistant mutants. Exp. Parasitol., 62: 61-70.
CrossRef - Becker, K., L. Tilley, J.L. Vennerstrom, D. Roberts, S. Rogerson and H. Ginsburg, 2004. Oxidative stress in malaria parasite infected-erythrocytes: Host-parasite interactions. Int. J. Parasitol., 34: 163-189.
CrossRefDirect Link - Coombs, G.H., G.D. Westrop, P. Suchan, G. Puzova and R.P. Hirt et al, 2004. The amitochondriate eukaryote trichomonas vaginalis contains a divergent thioredoxin linked peroxidoxin antioxidant system. J. Biol. Chem., 279: 5249-5256.
CrossRef - Gromer, S., L.D. Arscott, C.H.Jr. Williams, R.H. Schirmer and K. Becker, 1998. Human placenta thioredoxin reductase: Isolation of the selenoenzyme, steady state kinetics and inhibition by therapeutic gold compounds. J. Biol. Chem., 273: 20096-20101.
CrossRef - Holmgren, A., 2000. Antioxidant functions of thioredoxin and glutaredoxin systems. Antioxid. Redox. Signal., 2: 811-820.
CrossRef - Holmgren, A. and T.M. Bjorsnstedt, 1995. Thioredoxin and thioredoxin reductase. Methods Enzymol., 252: 199-208.
PubMed - Kapoor, G. and H.S. Banyal, 2009. Glutathione reductase and thioredoxin reductase: Novel antioxidant enzymes from Plasmodium berghei. Korean J. Parasitol., 47: 421-424.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Merril, C.R., D. Goldman, S.A. Sedman and M.H. Ebert, 1981. Ultrasensitive stain for protein in polyacrylamide gel shows regional variation in cerebrospinal fluid proteins. Science, 211: 1437-1438.
CrossRef - Oblong, J.E., P.Y. Gasdaska, K. Sherrill and G. Powis, 1993. Purification of human thioredoxin reductase: Properties and characterization by adsorption and circular dichroism spectroscopy. Biochemistry, 32: 7271-7277.
Direct Link - Watabe, S., Y. Makino, K. Ogawa, T. Hiroi, Y. Yamanoto and S.Y. Takahashi, 1999. Mitochondrial thioredoxin reductase in bovine adrenal cortex. Its Purification, Properties, Nucleotide/Amino Acid Sequences and Identification of Selenocysteine. Eur J. Biochem., 264: 74-84.
CrossRef - William, C.H.Jr., 1995. Mechanism and structure of thioredoxin reductase from E. coli. FASEB. J., 9: 1267-1276.
Direct Link - Williams, C.H., L.D. Arscott, S. Muller, B.W. Lennon and M.L. Ludwing, et al, 2000. Thioredoxin reductase two modes of catalysis have evolved. Eur. J. Biochem., 267: 6110-6117.
CrossRef - Banerjee, A.K., N. Arora and U.S.N. Murty, 2009. Structural model of the Plasmodium falciparum Thioredoxin reductase: A novel target for antimalarial drugs. J. Vector Borne Dis., 46: 171-183.
PubMedDirect Link