
T.V. Meenambigai
Department of Animal Biotechnology, Centralized Embryo Biotechnology Unit, Tamil Nadu Veterinary and Animal Sciences University, Chennai, Tamil Nadu, India
V. Sejian
Scientist, Division of Physiology and Biochemistry, Central Sheep and Wool Research Institute, Avikanagar, Rajasthan-304501, India
LiveDNA: 91.953
ORCID: 0000-0002-8224-4521
Asian Journal of Animal Sciences
Year: 2011 | Volume: 5 | Issue: 1 | Page No.: 1-18







ABSTRACT
Embryonic Stem (ES) cells are derived from blastocyst and these cells have the capability to generate all embryonic tissues in vitro. This propensity of ES cells has acquired considerable attention in recent years due to the promising potential for future cell replacement-based therapies. The in vitro differentiation capacity of ES cells provides unique opportunities for experimental analysis of gene regulation and function during cell commitment and differentiation in early embryogenesis. The ES cells are pluripotent cell lines with the capacity of self-renewal and a broad differentiation plasticity. They are derived from pre-implantation embryos and can be propagated as a homogeneous, uncommitted cell population for an almost unlimited period of time without losing their pluripotency and their stable karyotype. The ES cell technology is of high interest for researchers associated with livestock species. Simultaneously, research activities are being focused on characteristics and differentiation potential of Somatic Stem Cells (SSCs), unraveling an unexpected plasticity of these cell types. Somatic stem cells are found in differentiated tissues and can renew themselves in addition to generating the specialized cell types of the tissue from which they originate. Additional to discoveries of SSCs in tissues that were previously not thought to contain these kinds of cells, they also appear to be capable of developing into cell types of other tissues, but have a reduced differentiation potential as compared to embryo-derived stem cells. Therefore, SSCs are referred to as multipotent rather than pluripotent. This review summarizes characteristics of pluripotent ESCs in bovines and evaluates their potentials for in vitro propagation and differentiation as well as their potential uses in cell based therapies.
PDF Abstract XML References Citation
Received: March 25, 2010;
Accepted: May 02, 2010;
Published: July 27, 2010
How to cite this article
T.V. Meenambigai and V. Sejian, 2011. Insights into Embryonic Stem Cells of Bovines. Asian Journal of Animal Sciences, 5: 1-18.
DOI: 10.3923/ajas.2011.1.18
URL: https://scialert.net/abstract/?doi=ajas.2011.1.18
DOI: 10.3923/ajas.2011.1.18
URL: https://scialert.net/abstract/?doi=ajas.2011.1.18
INTRODUCTION
Embryonic Stem Cells (ESCs) are unique pluripotent cells derived from pre-implantation blastocyst-stage embryos. They can undergo asymmetric division whereby they either duplicate themselves or differentiate into another cell-type. While adult stem cells are multipotent and can only differentiate into a limited number of cell-types, ESCs are capable of differentiating into any cell-type. ESCs can proliferate indefinitely in an undifferentiated state (Evans and Kaufman, 1981). They express specific markers or characteristics including stage specific embryonic antigens, enzymatic activities such as alkaline phosphatase and telomerase and stemness genes that are rapidly down-regulated upon differentiation, including Oct4 and Nanog (Byrne et al., 2006). Alternatively, they can differentiate in vivo in teratomas into cells representing the three major germ layers: endoderm, mesoderm or ectoderm and they can be directed to differentiate in vitro into any of the more than 200 cell types present in the adult body (Savatier et al., 1994). Another characteristic of ESCs is their apparent ability to maintain a normal karyotype through large passage numbers. Indeed, a feature of primary cell cultures is the development of abnormal karyotypes as they become senescent. Since many human diseases result from defects in a single cell type, the potential to replace defective cells by cell or tissue replacement therapy involving differentiated human ESCs (hESCs) provides a possible cure for, or at least the alleviation of symptoms of, various degenerative diseases (Srivastava and Sejian, 2010).
The ESCs promise to open a new window in human existence. They possess the unique potential to replace our cells as they age, mutate and die. This scientific advance offers us the tantalizing possibility of maintaining our bodies in a state of mental and physical well-being inconceivable even one generation ago. As we enter the new millennium, the gift we are being offered is nothing short of the chance at longer, healthier lives. Stem cell based therapies for the repair and regeneration of various tissues and organs offer a paradigm shift that may provide alternative therapeutic solution for many diseases. Although, ESCs and Induced Pluripotent Stem Cells (iPSC) are theoretically highly beneficial, there are various limitations to their use imposed by cell regulation, ethical consideration and genetic manipulation (Cauffman et al., 2005). Adult Stem Cells (ASCs) on the other hand, are more easily available with neither ethical nor immunoreactive considerations, as long as they are of autologous tissue origin. In recent years much research has been focused on Mesenchymal Stem Cells (MSc) isolation from bone marrow stroma which have been shown to possess adipogenic, osteogenic, chondrogenic, myogenic and neurogenic potentials in vitro (Woodbury et al., 2000). As ESCs offers wide opportunity for therapeutic potential in animals, this review collates and synthesizes literature on characteristics of pluripotent ESCs in bovines and evaluates their potentials for in vitro propagation and differentiation as well as their potential uses in cell based therapies.
EPIGENETICS OF ESCs
Mammalian development originates from a single cell (zygote) that upon cleavage gives rise to totipotent blastomeres of the early embryo that eventually proliferate and differentiate into the wide variety of cell phenotypes found in the adult body. The complex pattern of gene expression governing development and differentiation is tightly regulated by epigenetic modifications, i.e., modifications of chromatin not involving changes in the DNA sequence. DNA methylation and histone methylation/acetylation are well known examples of epigenetic modifications. In general, DNA methylation is associated with the silencing of gene expression. Epigenetic errors can arise randomly or under the influence of the environment and often result in disease in humans. For example, DNA methylation has become increasingly implicated in cancer, as many cancer cells contain hypermethylated DNA that in turn can lead to the silencing of tumor suppressor genes by promoter methylation (Weksberg et al., 2003). Genomic imprinting is a form of the epigenetic program that involves modification of a gene or a chromosomal region that results in absolute or preferential, monoallelic-expression of a specific parental allele. Imprinting genes tend to cluster in the genome. Figure 1 describes the derivation potentials of embryonic stem cells.
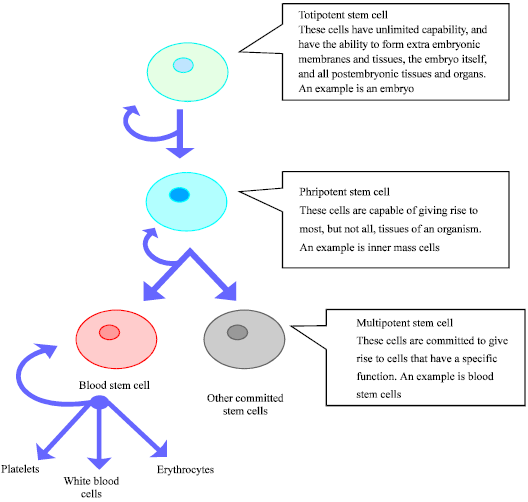 | |
| Fig. 1: | Derivation potentials of embryonic stem cells |
KEY CHARACTERISTICS OF ESCs
The ESCs are distinguished by two distinctive properties: their pluripotency and their capability to self-renew themselves indefinitely (Ying et al., 2003). ESCs are pluripotent, that is, they are able to differentiate into all derivatives of the three primary germ layers: ectoderm, endoderm and mesoderm. These include each of the more than 220 cell types in the adult body. Pluripotency distinguishes embryonic stem cells from adult stem cells found in adults; while embryonic stem cells can generate all cell types in the body, adult stem cells are multipotent and can only produce a limited number of cell types. Additionally, under defined conditions, ESCs are capable of propagating themselves indefinitely. This allows ESCs to be employed as useful tools for both research and regenerative medicine, because they can produce limitless numbers of themselves for continued research or clinical use. Because of their plasticity and potentially unlimited capacity for self-renewal, ESC therapies have been proposed for regenerative medicine and tissue replacement after injury or disease. Diseases that could potentially be treated by pluripotent stem cells include a number of blood and immune-system related genetic diseases, cancers and disorders; juvenile diabetes; parkinson's; blindness and spinal cord injuries. Besides the ethical concerns of stem cell therapy, there is a technical problem of graft-versus-host disease associated with allogeneic stem cell transplantation. However, these problems associated with histocompatibility may be solved using autologous donor adult stem cells, therapeutic cloning, stem cell banks or more recently by reprogramming of somatic cells with defined factors (e.g., induced pluripotent stem cells).
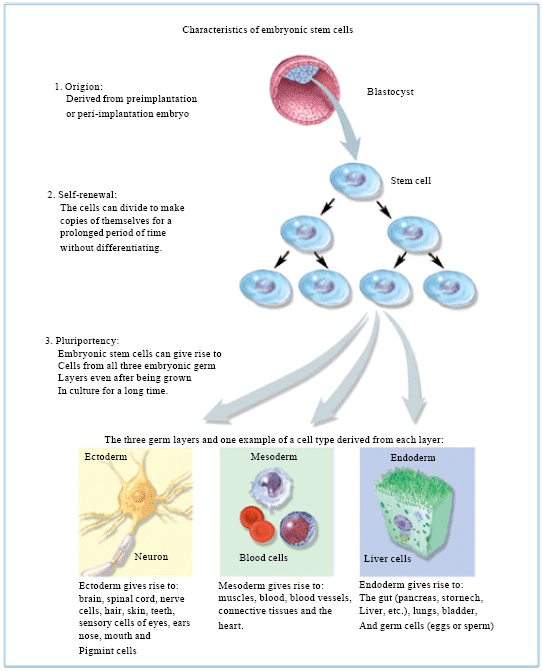 | |
| Fig. 2: | Embryonic stem cell characteristics |
Other potential uses of ESCs include investigation of early human development, study of genetic disease and as in vitro systems for toxicology testing. Figure 2 describes the key characteristics of ESCs.
STEM CELL LINEAGES
To ensure self-renewal, stem cells undergo two types of cell division. Symmetric division gives rise to two identical daughter cells both endowed with stem cell properties. Asymmetric division, on the other hand, produces only one stem cell and a progenitor cell with limited self-renewal potential. Progenitors can go through several rounds of cell division before terminally differentiating into a mature cell. It is possible that the molecular distinction between symmetric and asymmetric divisions lies in differential segregation of cell membrane proteins (such as receptors) between the daughter cells. An alternative theory is that stem cells remain undifferentiated due to environmental cues in their particular niche. Stem cells differentiate when they leave that niche or no longer receive those signals. Studies in 'Drosophila germarium have identified the signals dpp and adherens junctions that prevent germarium stem cells from differentiating. The signals that lead to reprogramming of cells to an embryonic-like state are also being investigated. These signal pathways include several transcription factors including the oncogene c-Myc (Vackova et al., 2007). Initial studies indicate that transformation of mice cells with a combination of these anti-differentiation signals can reverse differentiation and may allow adult cells to become pluripotent. However, the need to transform these cells with an oncogene may prevent the use of this approach in therapy.
ESC SOURCES
The ESC lines have been successfully isolated from mouse, monkey and human blastocysts, although outstanding derivations have also been made using embryos at pre-compaction stages (Eistetter, 1988; Delhaise et al., 1996; Strelchenko, 1996; Mitalipova et al., 2001). Most attempts to isolate and culture bESCs have been done with day 7-9 bovine blastocysts (Stice et al., 1996; Strelchenko, 1996; Cibelli et al., 1998; Iwasaki et al., 2000; Betts et al., 2001; Saito et al., 2003; Roach et al., 2006; Munoz et al., 2008) although, ESC-like cells are also isolated from day 12-14 embryos (Gjorret and Maddox-Hyttel, 2005). Yet, the optimal timing of bovine pre-implantation development to derive ESCs is still unknown. Attempts to derive bESC from zygotes and early cleavage stage embryos mostly failed (Strelchenko, 1996; Mitalipova et al., 2001), while only a single bovine embryonic cell line, generated from a two-cell embryo, has been cultured over 3 years (Mitalipova et al., 2001). Yet, when bovine morulae were used as starting material, efficient colony formation rates ranged over 60-70% (Stice et al., 1996; Strelchenko, 1996). The embryonic stage (morulae and day 7 blastocysts) used did not influence the efficiency in establishing bESC colonies. These results are nevertheless contrary to studies reporting that Day-8 hatched blastocysts yield a higher proportion of epiblast colonies than Inner Cell Masses (ICMs) isolated from day 9 blastocysts (41 and 13%, respectively) (Talbot et al., 1995).
Bovine embryos from different sources have been used to isolate bESCs (Meenambigai et al., 2010; Stalin et al., 2010). Yet there is only one published report which aimed to compare the feasibility of in vitro and in vivo derived embryos for the isolation of pluripotent cells (Talbot et al.,1995). This study demonstrated that in vivo derived (IVD) blastocysts, especially from early hatching blastocysts, were shown to be a source of pluripotent epiblasts superior to their in vitro produced (IVP) counterparts. The basis for any advantage by in vivo produced blastocyst to produce ESC lines is not known, although, a number of differences in morphology, metabolic rates, gene expression and susceptibility to cooling damage (Smith et al., 2005; Lonergan et al., 2006) have been reported between IVD and IVP bovine embryos. It is possible that the reduced number of cells present in the ICM of IVP bovine embryos (Van Soom et al., 1996) might affect survival of the ICM in culture, hindering the chances to establish ESC lines from IVP embryos. In fact Anderson et al. (1994) assumed that the factor that may affect survival of porcine ICMs in culture was the number of cells of the ICMs. In vivo derived embryos might be a better source of pluripotent cells, but their use as a starting material to isolate ESC is expensive and laborious (Meenambigai et al., 2009; Prabha et al., 2010). Therefore, it would be advisable to improve the procedures to derive ESCs from IVP embryos, as well as the ability of IVPICMs to yield ESCs. An obvious way to progress in this aspect would be increasing numbers of cells in these ICMs.
DERIVATION AND CULTURE CONDITIONS FOR ESC
Table 1 depicts the different types of cell culture media for culturing embryonic stem cells. Culture conditions close to those established for murine ESC culture were successfully used to derive monkey (Thomson et al., 1995) and human ESCs (Thomson et al., 1998). Nevertheless, it soon became evident that some factors required for the maintenance of mESC pluripotency were not only dispensable in maintaining hESC pluripotency but were also detrimental. As an example, this occurred with BMP4, a member of the transforming growth factor-b (TGF-b) family involved in controlling mESC differentiation that induces differentiation of human ESCs into trophoblast cells (Xu et al., 2002). Since then, considerable amount of data have been published over differences between mouse and human pluripotency maintaining factors and signaling pathways (Renard et al., 2007) (Fig. 3). Until now, following a similar approach to primate ESC isolation, most attempts to culture bESC have been inspired by the original culture methods for mESC of Evans and Kaufman (1981).
| Table 1: | Different types of cell culture media for culturing embryonic stem cells |
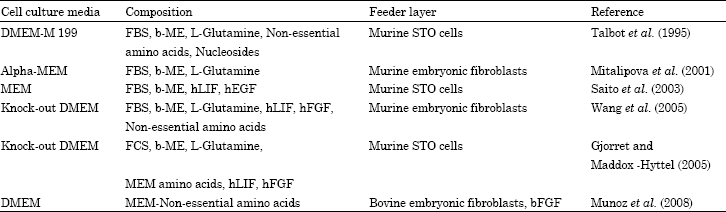 | |
| bFGF: Basic fibroblast growth factor, b-ME: b-mercaptoethanol, DMEM: Dulbecco’s Modified Eagle’s Medium, FBS: Foetal bovine serum; FCS: Foetal calf serum, hEGF: Human epidermal growth factor, hLIF: Human leukaemia inhibitory factor; MEM: Minimum essential medium | |
 | |
| Fig. 3: | Multipotential differentiation characteristics of embryonic stem cells |
Bovine ESCs are usually cultured on mouse embryonic fibroblasts (primary MEF or transformed STO cells). Culture media consists of Dulbecco’s Modified Eagle’s Medium supplemented with foetal bovine serum, L-glutamine, 2-b mercaptoethanol and different growth factors, mostly Leukaemia Inhibitory Factor (LIF) and Epidermal Growth Factor (EGF) (for comparison of some bESC culture conditions (Table 1). Yet it is likely that culture conditions suitable to maintain mESC could be inadequate to maintain undifferentiated bESC. Preliminary studies by Keefer et al. (2007) showed that the bovine ICM and its primary outgrowths express the LIF receptor and gp130 transducer. Yet, LIF did not improve the establishment and maintenance of ESCs from other ungulates (Vackova et al., 2007) although its presence in pig ESC culture medium prevented EB formation (Brevini et al., 2007). It can be speculated that, such as in hESC, stimulation of the STAT3 pathway by LIF might not induce proliferation of ungulate ESCs.
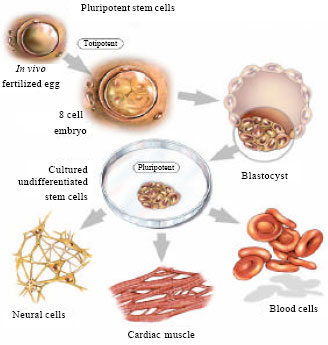
Similarly, some growth factors such as TGB-b, EGF or insulin-like growth factors (IGFs) found to suppress differentiation of mESCs ] did not inhibit differentiation of porcine is one of the possible explanations for the failure to isolate these cells. Identification of specific pluripotency signalling pathways will help to determine which growth factors are beneficial or which ones are inappropriate for establishing a successful bESC culture. Figure 3 describes the multipotential differentiation characteristics of ESCs.
SIMPLIFIED SCHEME OF STEM CELL POPULATIONS
Particular stem cell types are classified based on their differentiation potentials. The zypote and morula stages can give rise to both embryonic and extra embryonic tissues and hence can generate a complete embryo. The three germ layers, as well as embryonic germ cells, originate from the ESCs from the inner cell mass of the blastocysts. Adult stem cells produce progenitor cells and differentiated tissue. Figure 4 describes the differentiation potentials of stem cell populations.
EXPRESSION PROFILING OF ESC
Morphology, as well as the capacity to differentiate in vitro through Embryoid Body (EB) formation, was one of the two defining criteria initially used to identify bESC cultures. Other traits such as small size, rounded shape or high nucleus to cytoplasm ratio were used to define bESC lines. Yet, cells belonging to trophectoderm (TE) and visceral endoderm, which usually can be found in blastocysts or isolated ICMs primary cultures, may be confounded with ESC if solely morphological features are used as evaluating criteria. Bovine blastocyst-derived TE and endoderm cell lines have been thoroughly characterized not only by morphological criteria but also by the expression tissue-specific marker. For instance, transferrin is a definitive marker for bovine blastocyst-derived endoderm cell lines (Talbot et al., 2000). Therefore, the combined use of morphological criteria and the analysis of extra-embryonic markers is suggested to truly identify bESC and/or rule out the presence of TE or visceral endoderm cells in ESC cultures. A useful strategy to characterize ESC lines is to analyze the expression of pluripotency-related molecular markers. Unfortunately, until now, no specific markers have been identified in bovine. Therefore, markers associated to pluripotency in other species (heterospecific pluripotency markers) such as stage-specific embryonic antigens (SSEA-1, -3, -4) have been used to characterize bESC. SSEAs are developmentally regulated cell surface antigens expressed by murine and human pluripotent cells (Fig. 1).
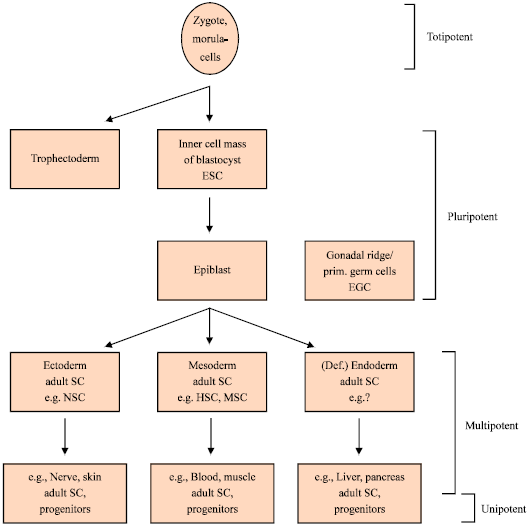 | |
| Fig. 4: | Differentiation potentials of stem cell populations. Particular stem cell types are classified based on their differentiation potentials. The zygote and morula stages can give rise to both embryonic and extra embryonic tissues and hence can generate a complete embryo. The three germ layers, as well as embryonic germ cells, originate from the embryonic stem cells from the inner cell mass of the blastocysts. Adult stem cells produce progenitor cells and differentiated tissue. Figure modified after keller (2005) |
The mESCs strongly express SSEA-1 (Solter and Knowles, 1978; Gooi et al., 1981), whereas, differentiated mESCs are characterized by the loss of SSEA-1 expression and in some instances, by the appearance of SSEA-3 and SSEA-4 (Solter and Knowles, 1979).
In contrast, hESCs typically express SSEA-3 and SSEA-4, but not SSEA-1 and their differentiation is characterized by down-regulation of SSEA-3 and SSEA-4 and up-regulation of SSEA-1 (Andrews et al., 1984; Fenderson et al., 1987). Undifferentiated hESCs also express the keratin sulphate-associated antigens TRA-1-60 and TRA-1-81 (Andrews et al., 1984). In bovine, a positive staining for SSEA-1, SSEA-3 and SSEA-4 was reported in three embryonic cell lines derived from pre-compaction embryos (Mitalipova et al., 2001). Similarly, SSEA-1 expression was also detected by Saito et al. (2003), while none of the bovine ES-like cells analysed by these authors were found positive for SSEA-3 or SSEA-4. In contrast, Wang et al. (2005) reported a positive SSEA-4 staining in the absence of SSEA-1 staining in five ESC lines. Munoz et al. (2008) reported positive staining for SSEA-4, TRA- 1-60 and TRA-1-81 in bESC-like cells.
Unfortunately, the above antigens were not only present in the ICM of bovine blastocysts but also in the TE (Fig. 1). Therefore, in bovine, these markers are not specific for undifferentiated and/or pluripotent cells. The use of such markers to characterize bESC may mislead researchers into isolating and culturing TE derived cells instead of ESCs. The expression of SSEAs, in the TE of bovine blastocysts, was unexpected, considering that for a long time SSEAs have been used to characterize undifferentiated bESCs. Nevertheless, it was not totally surprising as bovine TE cells show a slow differentiating phenotype characterized by the co-expression of epiblast-specifying genes (OCT-4, SOX-2, NANOG) and proteins (OCT-4, NANOG) and trophoblast-specific genes (CDX-2, HAND1, ETS-2, IFN-TAU, C12) (Kirchhof et al., 2000; Degrell et al., 2005; Munoz et al., 2008). Therefore in bovine, the expression of markers which are associated to pluripotency in other species (SSEA-4, TRA-1-60, TRA-1-81, OCT-4, NANOG) is not restricted to pluripotent cells (Fig. 1, 2), which is a warning to validate any pluripotency marker before its heterospecific use. An additional difficulty to characterize bESC is that available antibodies currently used to characterize ESCs are produced using mouse or human proteins as immunogens. Therefore, their ability to cross-react with the appropriated bovine protein should be evaluated before their use.
It is generally, assumed that ESC biology is regulated through transcriptional mechanisms, but the definition of a stem cell remains largely functional. The developmental capacity of ESC lines requires a set of genes that are not expressed in other cell types and knowledge of the intricate mechanisms regulating ESC pluripotentiality and differentiation potential is currently limited to a few signaling pathways (e.g., LIF, BMP, Wnt) and regulatory factors (e.g., Oct-3/4, Nanog) (Munoz et al., 2008). Theoretically, a comprehensive analysis of a cellular transcriptome (i.e., all the RNAs present in a cell type) should be sufficient to define the molecular phenotype of stem cells and establish the determinants of ESC choice. The underlying hypothesis behind these assumptions suggests that some mRNAs will be uniquely or more abundantly expressed in embryonic and/or adult stem cells than in any other cell type and that comparisons among cell populations will reveal these differences. Although several transcriptome-based (microarrays or SAGE) studies have now been published, which claim to have identified potential stemness-associated factors, a closer inspection of the data indicates that the identification of stemness factors has proved elusive (Byrne et al., 2006). This is true for both mouse and human ESCs. The reasons most frequently cited for variations among studies include cell lines, culturing conditions, array and hybridization protocols, data analysis and potentially contaminating cells.
Additionally, many of the studies in mice focused on comparisons among ESCs with adult stem cells, because of earlier studies suggesting a broader potential or plasticity of ASCs than previously believed (Niimi et al., 2005); however, this broader plasticity of primary isolates of many adult stem cells has recently been called into question (Niimi et al., 2005). The identification of stemness genes by these approaches, therefore, remains the topic of lively debate and much conjecture. Finally, the phenotype of ESCs must also involve complex processes that alter protein abundance both as a consequence of gene activation and processing (transcription, splicing, etc.), as well as regulatory events associated with translation and Post Translational Modifications (PTM). Proteomic approaches are therefore required to visualize and interpret the phenotype of undifferentiated ESCs. Figure 5 describes the combinatorial signaling pathways in maintaining mouse ESC pluripotency.
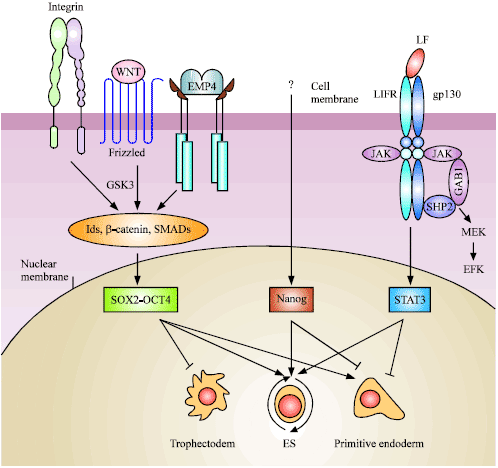 | |
| Fig. 5: | Combinatorial signaling pathways in maintaining mouse ESC pluripotency (Boiani and Scholer, 2005) |
MOLECULAR STRATEGY FOR ESC IDENTIFICATION
The pluripotent properties of ESCs are the basis of gene targeting technologies used to create mutant mouse strains (transgenic and knockout) for gene function studies in vivo. The in vitro differentiation of ESCs allows investigators to study cell differentiation and gene regulation, as well as pharmacological effects on functionally active cells (e.g., cardiomyocytes). Potentially, the greatest importance is to establish strategies for cell therapy and gene therapy. The differentiation of ESC can also provide a very useful system for the identification of genes involved in the development of a specific cell line. Commonly used methods including hanging drop and culture in suspension, result in the initial formation of multi-ESC aggregates and further differentiation into EBs (Mitsui et al., 2003). With these methods, it is difficult to detect the phenotypic change caused by one of these ES cells following genetic manipulation. As a result, previous approaches using gene-trap to identify genes involved in ESC differentiation required the isolation of genetically modified individual clones and the differentiation had to be handled separately (Mitsui et al., 2003).
Furthermore, the differentiation stage of individual EBs in the same preparation varies dramatically due to the varying size of the initial aggregate formed and subsequent processing. Although single ESC -derived Ebs can be generated in methylcellulose-based semisolid media (available from StemCell Technologies, Vancouver, BC, Canada), the EB yield is low (50-100 EBs/35-mm dish) and takes up to 20 days for differentiation (Niimi et al., 2005). To speed up large-scale gene discovery during ES cell differentiation, there is a need to develop a method for the differentiation of EBs from single ESC without the requirement of isolating cells.
ALKALINE PHOSPHATASE AND ESC-SPECIFIC MARKER STAINING
Alkaline phosphatase synthesis is specific for ESC. Alkaline phosphatase (ALPL) was detected histochemically following fixation of ES cells with 4% paraformaldehyde and nitro-blue tetrazolium chloride/5-bromo-4-chloro-3-indolylphosphate toluidine (NBT/BCIP) being used as substrate. The ALPL-expressing cells would stain dark blue. The ESCs can be immunohistochemically detected with monoclonal antibodies against mouse stage-specific embryonic antigen (SSEA) 1 (1:30), (SSEA) 4 (1:30), tumor rejection antigen gp96 (TRA1; two different antibodies used for TRA1, one recognizing a sialidase-sensitive epitope and one that reacts with an unknown epitope; 1:20), or rabbit OCT4 (1:500). The appropriate secondary antibodies, horse antimouse immunoglobulin G (IgG), goat anti-mouse IgM, or sheep anti-rabbit IgG, were used to amplify the signals. Detection of specific binding was performed with an Elite ABC peroxidase staining kit (Vector Laboratories, Inc., Burlingame, CA) and with 3,3 9-diaminobenzidine (Vector Laboratories) as substrate. Positive staining was gray-black in color. Staining controls using secondary antibodies alone also were included. The putative ES cells lines were at passages 14–16 at the time that marker expression was analyzed. The IVF blastocysts also were stained as described above. The primary antibodies for SSEA1, SSEA4 and OCT4 were localized by fluorescein isothiocyanate-conjugated goat anti-mouse IgM, horse anti-mouse IgG, or sheep anti-rabbit IgG (1:200). Finally, the samples were washed, mounted on glass slides and examined by fluorescence microscopy (Wang et al., 2005).
REGULATION OF DIFFERENTIATION
Figure 6A-F depict the ESCs derived embryoid bodies. For differentiation, ESCs were washed with 10 mL Phosphate-Buffered Saline (PBS) and then trypsinized. Well-dissociated single ESCs were transferred to ultra low cluster 6-well plate (Corning, Corning, NY, USA) at 1000-1500 cells/well in 1.5 mL LIF-free KnockOut DMEM supplemented with 20% FBS, pyruvate, nonessential amino acid and mercaptoethanol.
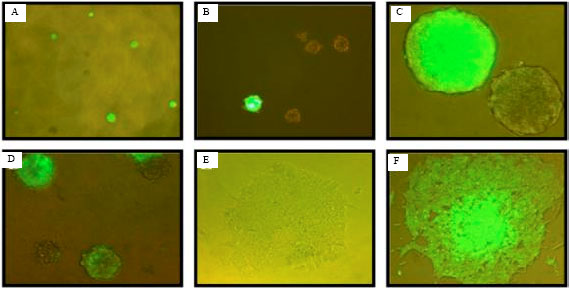 | |
| Fig. 6: | ESC derived embryoid bodies. Fluorescent images of embryoid bodies (EBs) differentiated with a plate shaker--Ordinary and green fluorescent protein-positive (GFP) embryonic stem (ES) cells were mixed at the ratio of 3:1 before differentiation. The EBs were excited at 480 nm. (A–C) 1000 mixed cells/well in 6-well plate on day 3, day 4 (10x) and day 5 (20x), respectively. (D) 2000 mixed cells/well on day 5 (10x). (E and F) GFP- and GFP single ES cell-derived EBs on day 7 after attachment to gelatin-coated dish (5x) |
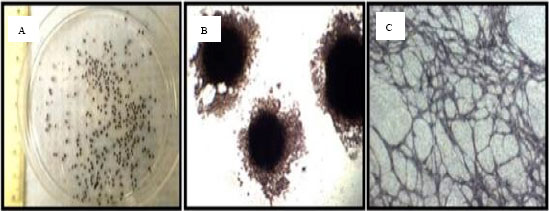 | |
| Fig. 7: | Embryoid Bodies (EBs) differentiated with a plate shaker and a spinner flask (A and B) EBs after attachment to gelatin-coated dish. (C) Vascular-like structure in EBs shown by immunochemical staining with anti-PECAM immunoglobulin G (IgG) (20x) |
To prevent any ES cells from aggregating during the differentiation, the plate was placed on a titer plate shaker (Lab-Line Instruments, Melrose Park, IL, USA) at the speed of approximately 120 rpm in a cell culture incubator for 3 days. About 250 cystic EBs were either transferred onto 100 mm 0.2% gelatin-coated dishes for attachment and further differentiation and growth or transferred into a spinner flask. To calculate the cell numbers and shaking speed, three-fourths of the ES cells were mixed with one-fourth of the ES cells that were constitutively expressing green fluorescence protein (GFP) driven by an EF-1α promoter. The cells were seeded at different densities (250-500, 500-1000, 1000-1500 and 1500-2000) and the plate was shaken at variable speeds. Only in the case where EBs were derived from single ES cells, can the whole EB be green (GFP) or not green at all. However, if EBs were derived from multiple ES cell aggregates, the EB would be partially green. As shown in Fig. 6, A, B, C and F, only at the cell density and shaking speed defined above were the whole green and non-green EBs obtained. Partially green EBs were seen if the cell density was higher and the shaking speed was too low. Although single ESC -derived EBs were obtained from the lower cell density, the size of the EB was generally smaller. Figure 7a-c describes the differentiation potential of embryoid bodies (Niimi et al., 2005).
Ordinary and green fluorescent protein-positive (GFP) Embryonic Stem (ES) cells were mixed at the ratio of 3:1 before differentiation.
DIFFERENTIATION OF BESC IN VIVO AND EX VIVO
The differentiation ability of bESC in vitro is evaluated by EB formation. The EBs are aggregates of stem cells whose development is reminiscent of early embryogenesis. Maintenance of bESC in a suspension culture (Strelchenko 1996) or in the absence of a feeder layer (Saito et al., 2003; Wang et al., 2005) initiates the formation of EBs. The EBs are composed of two layers of cells, ectoderm-like cells covered by a thin layer of endoderm like cells, with heterogeneous cellular particles within the cavity. The cells of bovine EBs give rise to a wide variety of differentiated cell types, including derivatives of the three germ layers (Saito et al., 2003; Wang et al., 2005). This ability is a proof of their pluripotent-differentiation character in vitro. The ability of bESC to participate in the embryogenesis has been proven only after a short propagation period in vitro. Chimeric transgenic calves have been born after injection of bESC (passage 3) into cleavage stage embryos (Cibelli et al., 1998), embryo aggregation with bESC (passage 9 -13) (Iwasaki et al., 2000). Yet integration of ESCs into the germ line, one of the properties used to define ESCs, have not been achieved in any of these experiments. Calves were also successfully cloned using ES-like cells as donor nuclei (short cultured ICMs or passage 14-18) (Sims and First 1994; Saito et al., 2003), but the use of ESC in nuclear transfer cannot be taken as a proof of pluripotency as cloned animals have been produced from fully differentiated somatic cell nuclei (Kato et al., 2000; Wakayama and Yanagimachi, 2001). Figure 8 describes the isolation and identification of mouse embryonic stem cells.
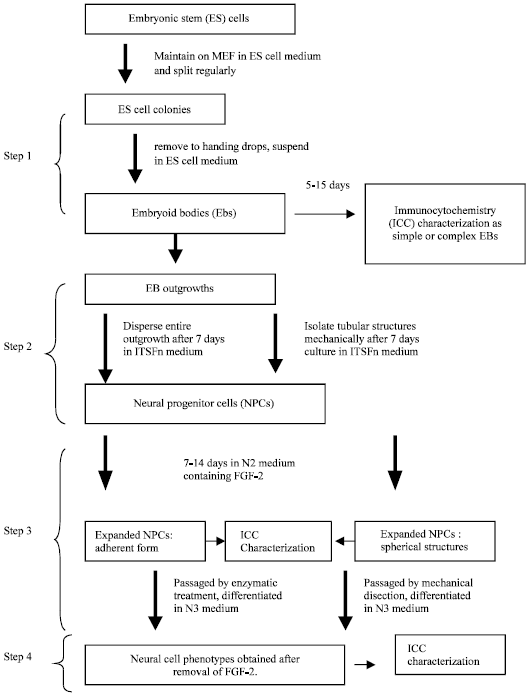 | |
| Fig. 8: | Isolation and Identification of mouse embryonic stem cells |
APPLICATIONS OF ESC
Figure 9 depicts the salient applications of ESC. The potential application of ESCs depends on the derivation of specialized lines of differentiated cells. But in real sense these are challenges which needs to be prevailed for the successful application of ESCs. For these reasons, three methods of therapeutic cloning were proposed, suitable for preparing pluripotent human embryonic stem cells with well defined genetic information from which desired differentiation would then follow:
| • | The replacement of the nucleus of an oocyte with the nucleus of an adult cell of a given subject, followed by embryonic development to the stage of blastocyst and the use of the Inner Cell Mass (ICM) in order to obtain ES cells and, from these, the desired differentiated cells |
| • | The transfer of a nucleus of a cell of a given subject into an oocyte of another animal. An eventual success in this procedure should lead to the development of a human embryo, to be used as in the preceding case |
| • | The reprogramming of the nucleus of a cell of a given subject by fusing the ES cytoplast with a somatic cell karyoplast, thus obtaining a cybrid. This is a possibility which is still under study. In any event, this method too would seem to demand a prior preparation of ES cells from human embryos. Figure 8 describes the salient applications of ESCs |
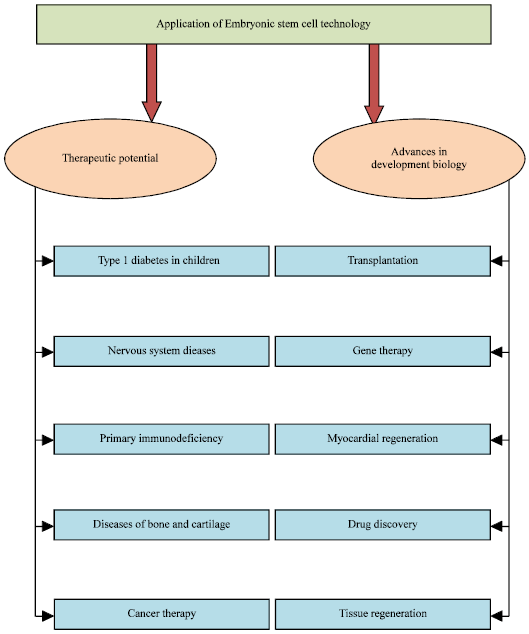 | |
| Fig. 9: | Application of ESC in therapeutics and advances in developmental biology |
REFERENCES
- Anderson G.B., S.J. Choi and R.H. Bondurant, 1994. Survival of porcine inner cell masses in culture after injection into blastocysts. Theriogenology, 42: 204-212.
PubMed - Andrews, P.W., I. Damjanov, D. Simon, G.S. Banting, C. Carlin, N.C. Dracopoli and J. Fogh, 1984. Pluripotency embryonal carcinoma clones derived from human teratocarcinoma cell line Tera-2: Differentiation in vivo and in vitro. Lab. Invest., 50: 147-162.
PubMed - Betts, D., V. Bordignon, J. Hill, Q. Winger, M. Westhusin, L. Smith and W. King, 2001. Reprogramming of telomerase activity and rebuilding of telomere length in cloned cattle. Proc. Natl. Acad. Sci. USA., 98: 1077-1082.
Direct Link - Boiani, M. and H.R. Scholer, 2005. Regulatory networks in embryo-derived pluripotent stem cells. Nature Rev. Mol. Cell Biol., 6: 872-881.
CrossRef - Brevini, T.A., S. Antonini, F. Cillo, M. Crestan and F. Gandolfi, 2007. Porcine embryonic stem cells: Facts, challenges and hopes. Theriogenology, 68: S206-S213.
PubMedDirect Link - Byrne, J.A., S.M. Mitalipov and D.P. Wolf, 2006. Current progress with primate Embryonic stem cells. Curr. Stem. Cell. Res. Ther., 1: 127-138.
CrossRef - Cauffman, G., H, Van de Velde, I. Liebaers and A. Steirteghem, 2005. Oct-4 mRNA and protein expression during human preimplantation development. Mol. Human Reprod., 11: 173-181.
PubMed - Cibelli, J.B., S.L. Stice, P.J. Golueke, J.J. Kane and J. Jerry et al., 1998. Transgenic bovine chimeric offspring produced from somatic cell-derived stemlike cells. Nat. Biotechnol., 16: 642-646.
PubMed - Eistetter, H.R., 1988. A mouse pluripotent embryonal stem cell line stage-specifically regulates expression of homeo-box containing DNA sequences during differentiation in vitro. Eur. J. Cell. Biol., 45: 315-321.
PubMed - Evans, M.J. and M.H. Kaufman, 1981. Establishment in culture of pluripotential cells from mouse embryos. Nature, 292: 154-156.
CrossRef - Gooi, H.C., T. Feizi, A. Kapadia, B.B. Knowles, D. Solter and M.J. Evans, 1981. Stage-specific embryonic antigen involves alpha 1 goes to 3 fucosylated type 2 blood group chains. Nature, 292: 156-158.
CrossRef - Gjorret, J.O. and P. Maddox-Hyttel, 2005. Attempts towards derivation and establishment of bovine embryonic stem cell-like cultures. Reprod. Fertil. Dev., 17: 113-124.
CrossRef - Iwasaki, S., K.H. Campbell, C. Galli and K. Akiyama, 2000. Production of live calves derived from embryonic stem-like cells aggregated with tetraploid embryos. Biol. Reprod., 62: 470-475.
CrossRefDirect Link - Kato, Y., T. Tani and Y. Tsunoda, 2000. Cloning of calves from various somatic cell types of male and female adult, newborn and fetal cows. J. Reprod. Fertil., 120: 231-237.
PubMedDirect Link - Keefer, C.L., D. Pant, L. Blomberg and N.C. Talbot, 2007. Challenges and prospects for the establishment of embryonic stem cell lines of domestic ungulates. Anim. Reprod. Sci., 98: 147-168.
PubMed - Keller, G., 2005. Embryonic stem cell differentiation: Emergence of a new era in biology and medicine. Genes Dev., 19: 1129-1155.
CrossRef - Kirchhof, N., J.W. Carnwath, E. Lemme, K. Anastassiadis, H. Schole and H. Niemann, 2000. Expression pattern of Oct-4 in preimplantation embryos of different species. Biol. Reprod., 63: 1698-1705.
CrossRefDirect Link - Lonergan, P., T. Fair, D. Corcoran and A.C. Evans, 2006. Effect of culture environment on gene expression and developmental characteristics in IVF-derived embryos. Theriogenology, 65: 137-152.
CrossRefPubMedDirect Link - Meenambigai, T.V., R. Gopinath, A. Rajesh, A. Palanisamy, S.S. Kumar, K. Brindha and K. Kumanan, 2010. Isolation, characterization and differentiation of quail bone marrow derived mesenchymal stem cells. Proceedings of the 16th Annual Convention of Indian Society for Veterinary Immunology and Biotechnology and National Symposium on Novel Biotechnological and Immunological Intervention in Mitigation of Climate Changes on Production and Protection of Livestock and Poultry, April 8-10, Veteinary College and Research Institute, Namakkal, Tamil Nadu, India, pp: 83-83.
- Mitalipova, M., Z. Beyhan and N. First, 2001. Pluripotency of bovine embryonic cell line derived from precompacting embryos. Cloning, 3: 59-67.
PubMed - Mitsui, K., Y. Tokuzawa, H. Itoh, K. Segawa and M. Murakami et al., 2003. The homeoprotein nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell, 113: 631-642.
CrossRefPubMedDirect Link - Munoz, M., A. Rodriguez, C. De Frutos, J.N. Caamano, C. Diez, N. Facal and E. Gomez, 2008. Conventional pluripotency markers are unspecific for bovine embryonic derived cell-lines. Theriogenology, 69: 1159-1164.
Direct Link - Niimi, A., K. Mu-Young, T. Lian, L. Hong Mei and W. Xiaoyun et al., 2005. Single embryonic stem cell derived embryoid bodies for gene screening. Biotechniques, 38: 349-352.
Direct Link - Prabha, S., T.V. Meenambigai, S. Balasubramanian, A. Palanisamy and K. Kumanan, 2010. Glucose transporter 1 gene expression in developmentally regulated in vitro derived ovine preimplantation embryos by real time PCR. Proceedings of the XVI Annual Convention of Indian Society for Veterinary Immunology and Biotechnology and National Symposium on Novel Biotechnological and Immunological Intervention in Mitigation of Climate Changes on Production and Protection of Livestock and Poultry, April 8-10, Veteinary College and Research Institute, Namakkal, Tamil Nadu, India, pp: 84-84.
- Renard, J.P., J. Maruotti, A. Jouneau and X. Vignon, 2007. Nuclear reprogramming and pluripotency of embryonic cells: Application to the isolation of embryonic stem cells in farm animals. Theriogenology, 68: S196-S205.
Direct Link - Roach, M., L. Wang, X. Yang and X.C. Tian, 2006. Bovine embryonic stem cells. Methods Enzymol., 418: 21-37.
CrossRef - Saito, S., K. Sawai, H. Ugai, S. Moriyasu and A. Minamihashi et al., 2003. Generation of cloned calves and transgenic chimeric embryos from bovine embryonic stem-like cells. Biochem. Biophys. Res. Commun., 309: 104-113.
CrossRefDirect Link - Savatier, P., S. Huang, L. Szekely, K.G. Wiman and J. Samarut, 1994. Contrasting patterns of retinoblastoma protein expression in mouse embryonic stem cells and embryonic fibroblasts. Oncogene, 9: 809-818.
PubMed - Sims, M. and N.L.M. First, 1994. Production of calves by transfer of nuclei from cultured inner cell mass cells. Proc. Natl. Acad. Sci. USA., 91: 6143-6147.
Direct Link - Smith, S.L., R.E. Everts, X.C. Tian, F. Du and L.Y. Sung et al., 2005. Global gene expression profiles reveal significant nuclear reprogramming by the blastocyst stage after cloning. Proc. Natl. Acad. Sci. USA., 102: 17582-17587.
CrossRef - Solter, D. and B.B. Knowles, 1978. Monoclonal antibody defining a stage-specific mouse embryonic antigen (SSEA-1). Proc. Natl. Acad. Sci. USA., 75: 5565-5569.
Direct Link - Solter, D. and B.B. Knowles, 1979. Developmental stage-specific antigens during mouse embryogenesis. Curr. Top. Dev. Biol., 13: 139-165.
CrossRef - Srivastava, R.S. and V. Sejian, 2010. Recent Advances in Neuronal Stem Cell Research. In: Short Course on 'Recent Advances in Stem Cell Research, Bag, S., A.C. Majumdar, B.C. Das and G. Tarusharma (Eds.). Centre of Advanced Faculty Training in Veterinary Physiology, Division of Physiology and Climatology, Indian Veterinary Research Institute, Izatnagar, Bareilly, India, pp: 57-60.
- Stalin, V., T.V. Meenambigai, A. Palanisamy, S. Sathesh-Kumar, K. Brindha, S. Nithya and K. Kumanan, 2010. Isolation and characterization of buffalo embryonic stem cells. Proceedings of the 16th Annual Convention of Indian Society for Veterinary Immunology and Biotechnology and National Symposium on Novel Biotechnological and Immunological Intervention in Mitigation of Climate Changes on Production and Protection of Livestock and Poultry, April 8-10, Veteinary College and Research Institute, Namakkal, Tamil Nadu, India, pp: 134-134.
- Stice, S.L., N. Strelchenko, C.L. Keefer and L. Matthews, 1996. Pluripotent bovine embryonic cell lines diret embryonic development following nuclear transfer. Biol. Reprod., 54: 100-110.
CrossRefDirect Link - Talbot, N.C., A.M. Powell and Jr. C.E. Rexroad, 1995. In vitro pluripotency of epiblasts derived from bovine blastocysts. Mol. Reprod. Dev., 42: 35-52.
PubMed - Talbot, N.C., T.J. Caperna, J.L. Edwards, W. Garrett, K.D. Wells and A.D. Ealy, 2000. Bovine blastocyst-derived trophectoderm and endoderm cell cultures: interferon tau and transferring expression as respective in vitro markers. Biol. Reprod., 62: 235-247.
Direct Link - Thomson, J.A., J. Kalishman, T.G. Golos, M. Durning, C.P. Harris, R.A. Becker and J.P. Hearn, 1995. Isolation of a primate embryonic stem cell line. Proc. Natl. Acad. Sci. USA., 92: 7844-7848.
PubMed - Thomson, J.A., J. Itskovitz-Eldor, S.S. Shapiro, M.A. Waknitz, J.J. Swiergiel, V.S. Marshall and J.M. Jones, 1998. Embryonic stem cell lines derived from human blastocysts. Science, 282: 1145-1147.
CrossRef - Vackova, I., A. Ungrova and F. Lopes, 2007. Putative embryonic stem cell lines from pig embryos. J. Reprod. Dev., 53: 1137-1149.
CrossRefDirect Link - Van Soom, A., M Boerjan, M.T. Ysebaert and A. De Kruif, 1996. Cell allocation to the inner cell mass and the trophectoderm in bovine embryos cultured in two different media. Mol. Reprod. Dev., 45: 171-182.
Direct Link - Wang, L., E. Duan, L. Sung, B. Jeong, X. Yang and C. Tian, 2005. Generation and characterization of pluripotent stem cells from cloned bovine embryos. Biol. Reprod., 73: 149-155.
CrossRefDirect Link - Wakayama, T. and R. Yanagimachi, 2001. Mouse cloning with nucleus donor cells of different age and type. Mol. Reprod. Dev., 58: 376-383.
CrossRefPubMedDirect Link - Weksberg, R., A.C. Smith, J. Squire and P. Sadowski, 2003. Beckwith-Wiedemann syndrome demonstrates a role for epigenetic control of normal development. Hum. Mol. Genet., 12: R61-R68.
CrossRefDirect Link - Woodbury, D., E.J. Schwarz, D.J. Prockop and I.B. Black, 2000. Adult rat and human bone marrow stromal cellsdifferentiation into neurons. J. Neurosci Res., 61: 364-370.
PubMed - Ying, Q.L., J. Nichols, I. Chambers and A. Smith, 2003. BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell, 115: 281-292.
PubMed