
Oke Kingsley Oyediran
Amity Institute of Microbial Technology (AIMT), Amity University Rajasthan, Kant Kalwar, NH-11, Jaipur 303002, India
LiveDNA: 234.25151
Aseri Gajendar Kumar
Amity Institute of Microbial Technology (AIMT), Amity University Rajasthan, Kant Kalwar, NH-11, Jaipur 303002, India
Jain Neelam
Amity Institute of Biotechnology (AIB), Amity University Rajasthan, Kant Kalwar, NH-11, Jaipur 303002, India
Asian Journal of Agricultural Research
Year: 2018 | Volume: 12 | Issue: 1 | Page No.: 10-18







ABSTRACT
Background and Objective: Arbuscular mycorrhizal fungi (AMF) symbiosis is a major survival strategy for plants growing under environmental stresses. Present study aimed at investigating diversity and drivers of AMF in rhizosphere of tomato at arid and semi-arid regions of Indian desert. Materials and Methods: Soil and feeder root samples were collected from 24 tomato farms in 6 districts of arid and semi-arid regions of the desert during 2017 cropping season. Spores were analyzed using morphological and molecular (Illumina Miseq sequencing platform) methods. Results: About 18 species viz. 8 of Glomus, 3 each of Acaulospora and Sclerocystis, 2 each of Scutellospora and Gigaspora were isolated. Glomus mossae, G. intraradices and G. fasciculatum had highest frequency of occurrence (100%) followed by Gigaspora albida, G. margarita and A. bireculata (83% each), while other species ranged between 33-66%. Spore population showed strong positive correlations with root colonization, organic carbon and rainfall, fairly positive correlation with sand, pH, nitrogen and potassium, weak correlations with temperature, silt, clay and electrical conductivity and negatively significant correlation with phosphorous. Conclusion: Glomus species were dominant AMF, spore population and root colonization were higher and lower in arid and semi-arid districts respectively, while major drivers of AMF diversity were edapho-climatic factors.
PDF Abstract XML References Citation
How to cite this article
Oke Kingsley Oyediran, Aseri Gajendar Kumar and Jain Neelam, 2018. Arbuscular Mycorrhizal Fungi Associated with Rhizosphere of Tomato Grown in Arid and Semi-arid Regions of Indian Desert. Asian Journal of Agricultural Research, 12: 10-18.
DOI: 10.3923/ajar.2018.10.18
URL: https://scialert.net/abstract/?doi=ajar.2018.10.18
DOI: 10.3923/ajar.2018.10.18
URL: https://scialert.net/abstract/?doi=ajar.2018.10.18
INTRODUCTION
In the arid and semi-arid regions of Indian desert, drought, extreme temperature and progressive salinization of land are among the major abiotic stresses limiting plant growth and productivity1. The desert spreads over 1, 96,150 km2, occupying about 70% of western Rajasthan; lying between 25°45' N latitude and 70°75' E longitude2. Today, not only problems of abiotic stresses serious are in these degraded agroecosystems, it seems they are inevitably worsening3. Although, plants have a variety of biochemical and physiological mechanisms to cope with adverse environmental conditions, these mechanisms are often not sufficient enough to tolerate elevated and consistent environmental stress situations4. Furthermore, a number of conventional strategies such as selection and genetic engineering methods5, improvement of soil management, irrigation techniques6 and chemical priming7 had been introduced to improve stress tolerance in crops in this agriculturally vulnerable ecosystems with little outcomes.
Interestingly, in addition to increasing nutrient and mineral availability and uptake, different mechanisms have been proposed to be involved in tolerance to abiotic and biotic stresses8 alleviation by AMF plants6, increased ability for nutrient capturing and cycling and enhancement of plant health through increased protection against pathogen attack. Auge et al.9, further reported that AMF symbiosis is capable of influencing stomatal conductance (gs) that exerts positive control over water exchange rates and facilitation of water stable aggregates formation10. Several findings have also confirmed improved growth performances of AM-colonized salt stressed vegetable crop species such as clover, tomato, sweet basil, cucumber and lettuce11,12-14. Thus, a deep understanding of the composition and diversity of AMF in this ecologically stressed ecosystem would serve as a fundamental tool for delving into its functional profile and agricultural importance.
Hitherto, a lot of studies have confirmed the multiple contributions of AMF in the growth and development of most plants under abiotic stresses in wild or agro-ecosystem15. However, there is little information on the stress alleviating role of these organisms in the rhizosphere of most vegetable plants such as tomato (Solanum lycopersicum) in the arid and semi-arid ecosystem of Indian desert. Tomato is a ubiquitous vegetable, with production and consumption spreading all across the globe, making it one of the best known food ingredients and one of the most beloved vegetables11. Because tomato thrives well in warm, sunny conditions with no severe frost, it took well to Indian climate and today, India has become the second largest global producer (next to China) of tomato, producing over 18 million t of tomato annually16.
Thus, using both morphological and molecular techniques, the present study aimed at investigating the diversity and main drivers of AMF in the rhizosphere of tomato grown in the arid and semi-arid agro-ecological regions of Indian desert. The new knowledge from this study would be a valuable reference for a better understanding of the diversity and unique role of AMF in symbiosis with this important vegetable in the prevailing harsh environmental conditions of Indian desert.
MATERIALS AND METHODS
Study area: Six districts in the arid and semi-arid regions of western Rajasthan were selected for the study during the tomato planting season (May-August, 2017); 4 districts from arid region (Bikaner, Jaisalmer, Barmer and Jodhpur) and two districts from the semi-arid region (Ajmer and Jaipur). From each district, four notable irrigated tomato cultivation fields were chosen for sample collection. All study districts falls within geographical coordinates of 25°-45'N latitude and 70°-75' E longitude. Largest (38,401sq. km) and smallest (22,892 km2) land areas were found in Jaisalmer and Ajmer districts, respectively. Jaipur district had the highest altitude (487 m) while lowest (229 m) was obtained in Jaisalmer district. Mean monthly temperature was highest in the arid districts (between 45°C in Jodhpur and 49°C in Jaisalmer) while the semi-arid districts recorded 39 and 40°C in Ajmer and Jaipur, respectively. However, average monthly rainfall values in the arid districts were lower (ranging from 209 mm in Jaisalmer to 363 mm in Jodhpur) compared to the semi-arid districts (Ajmer, 557 mm; Jaipur, 601 mm). Important geographical features and climatic variations of the study districts were presented in Table 1 while locations of sample collection were shown in Fig. 1.
Soil sampling and analysis: Sampling was done during the middle of July, 2017 at a depth of 0-30 cm using soil auger after scrapping off the upper layer to remove foreign particles and litters. At least 8 soil and root samples were collected from the rhizosphere of tomato plant at each location and all samples collected from one location were put in sterilized polythene bag, labelled appropriately and mixed to form composite sample for each location. Samples were immediately taken to the laboratory and stored at 4°C for further analysis.
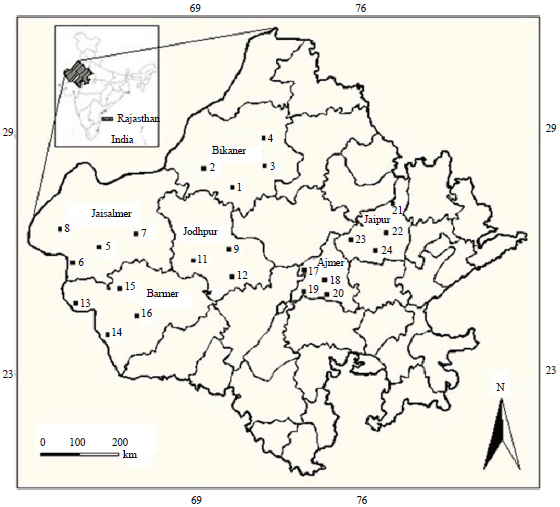 | |
| Fig. 1: | Districts showing sample collection sites |
| 1: Kolayat, 2: Nokha, 3: Bikaner, 4: Gersar, 5: Vinjorat, 6: Devikot, 7: Pokaran, 8: Jaisalmer, 9: Bandra, 10: Ranasar, 11: Jalwa, 12: Barmer, 13: Bawari, 14: Mogra, 15: Jodhpur, 16: Bilara, 17: Puskar, 18: Beawar, 19: Ajmer, 20: Kanpura, 21: Shapura, 22: Tala, 23: Chandwaji, 24: Achrol | |
| Source: www.mapsofindia.com | |
| Table 1: | Description of study districts |
 | |
| Table 2: | Soil physicochemical characteristics of the study districts |
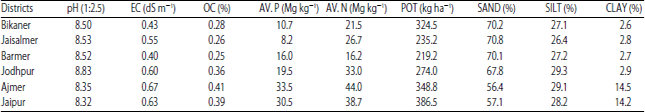 | |
| EC: Electrical conductivity, OC: Organic carbon, AV.P: Available phosphorous, AV.N: Available nitrogen, POT: Potash | |
Important soil parameters like pH and EC (soil: water ratio), organic carbon content, available nitrogen, available phosphorous, potassium content and soil texture were determined using the standard methods described by AOAC17. Values of the analyzed soil parameters were shown in Table 2.
Trap culture: The AMF were amplified using trap cultures due to the characteristic low level of spore densities in desert soils18. Five hundred gram dry weight field soil was mixed with autoclaved soil (1:1, v/v) and sown with pre-germinated, healthy and surface sterilized Triticum aestivum seeds in pots. All pots were irrigated regularly with sterile water for 45 days in green house at 25±30°C.
Isolation, morphological identification and enumeration of AMF spores: The AMF spores were isolated from rhizospheric soil samples following the Wet-Sieving and Decanting method of Gerdemann and Nicolson19. Isolated spores were kept in polyvinyl alcohol, lactic acid mixture in Melzer’s reagent and observed under the Olympus CH2Oi microscope for counting. Counted spores were expressed as number of spores per 100 g of soil. Spore identification was done based on spore size, spore color, wall layers and hyphal attachment using the specifications provided by INVAM20. After proper identification, percentage frequency of occurrence was determined using the method of Khade and Rodrique21.
Assessment of root infection and colonization: Tertiary roots were collected and washed with sterile water to remove all adhering soil debris. The roots were later cut into small pieces of approximately 1 cm length, subjected to differential staining22 and viewed under dissecting microscope (10 and 40×). Root segments containing vesicles, hartig nets, arbuscules and hyphae of endophytes were considered infected23. Root colonization was determined using the grid-line intersect method24. The root samples were placed in 7% KOH for 24 h and rinsed in water for clearing. Samples were then acidified in 3.5% HCl for 2 h and stained with 0.05% Trypan blue in 50% glycerol. Root fragments (1 cm) were mounted on slides in glycerol and viewed for intersections under the light microscope. Percentage root colonization was then expressed using the equation described by Philips and Hayman22 as:
Molecular analyses: The fast DNA isolation kit (Q-BIOgene; Heidelberg, Germany) was used to extract DNA from 50 g soil sample collected from each location. Extracted DNA were stored at -20°C for PCR reactions. DNA concentration and purity were checked using 1.0% agarose gel. Extracted sequences were amplified using the 18S rRNA gene and primer sets of AMV4.5N Forward 5'-AAGCTCGTAGTTGAATTTCG-3' and AMD R 5'-CCCAACTATCCCTATTAATCAT-3'. The PCR amplification was conducted using extracted method described by Xiao et al.25. Products of PCR were separated and purified using 1.5% agarose in 0.5×TBE and gel extraction kit (Axygen, Biosciences, NY, USA). The libraries were then sequenced using PE300 sequencing on MiSeq v3 Reagent kit (Illumina) on platform (Illumina, Inc., San Diego, CA, USA). Mothur software application (version 1.33.3) was used to analyze the DNA sequence reads. Aggregation (97%) of decoded information to operational taxonomic units (OTUs) was done using the method described by Edgar26 and blasting was done using nucleotide collection (nr nt–1) database. The blast hit with highest score was identified as equivalent species.
Data analysis: Operational taxonomic units (OTUs) richness, coverage, Chao’s and Shannon’s indexes were determined using Mothur software (version 1.33.3) application27 while indicator species analysis for the locations was determined using the method of Dufrene and Legendre28. Analysis of variance (one-way ANOVA) and level of significance (p<0.05 and p<0.01) were determined using the IBM SPSS Statistics V21×86 model Software Application. Pearson’s correlation co-efficient was used to assess the relationship between AMF spore population and various edapho-climatic factors.
RESULTS
Analysis of species diversity, spore population and root colonization: Results of AMF species diversity, spore population and root colonization analysis in the study districts were presented in Table 3. The number of sequence in the soil sample collected from each location ranged from 15238-29147. OTUs coverage in all district locations were up to 99% whilst OTUs number ranged from 45-81 with a genetic distance of 3%. Analysis of Chao’s index revealed higher OTUs richness in the arid district (between 70 in Jodhpur and 73 in Jaisalmer) over the semi-arid districts (Ajmer: 58, Jaipur: 57). Similarly, values obtained for AMF diversity using Shannon’s index were higher in the arid districts (ranging from 3.13 in Jodhpur to 3.74 in Bikaner) over semi-arid districts (Ajmer: 2.37, Jaipur: 2.74). The AMF spore population values of the studied tomato fields were also higher in the arid district compared to the semi-arid districts and varied between 301 (100 g–1) in Jaipur district to 494 (100g–1) in Jaisalmer district.
| Table 3: | Data on sequence analysis, AMF population and root colonization |
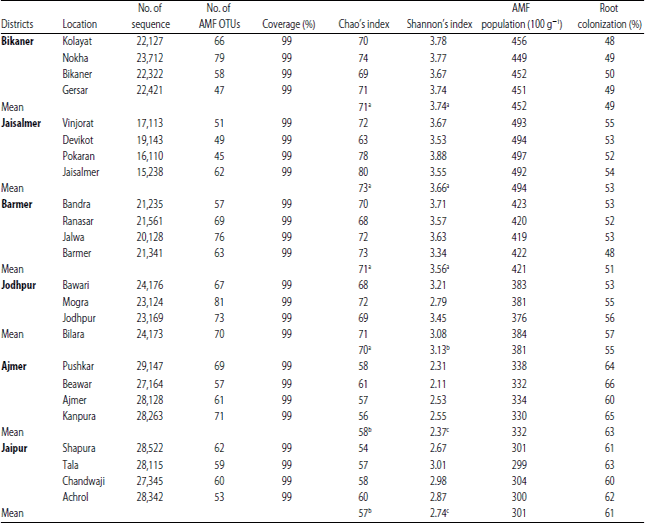 | |
| OTUs defined at cutoff of 3% difference in sequence. Same letter in the same column do not differ significantly at 5% level of probability | |
Meanwhile, higher values of percentage root colonization were recorded in the semi-arid districts (Ajmer: 63%, Jaipur: 61%) over the arid districts (between 49% in Bikaner and 55% in Jodhpur).
Assessment of AMF composition and frequency of occurrence: Variation was observed in the AMF composition among the soil samples in all the locations (Table 4). The identified OTUs were affiliated with 5 AMF genera (Acaulospora, Sclerocystis, Glomus, Scutellospora and Gigaspora) with a total of 18 AMF species. Highest number of species was recorded from Glomus (8) which served as indicator species in all the locations. This was followed by Acaulospora and Sclerocystis with 3 species each, while the genus Scutellospora and Gigaspora recorded 2 species each. Also, highest percentage frequency of occurrence (100%) was obtained among three species of Glomus viz; G. intraradices, G. mossae and G. fasciculatum followed by Gigaspora species (G. albida and G. margarita) and Acaulospora species (A. bireculata) each with 83% frequency of occurrence. The percentage frequency of occurrence of other AMF species ranged between 33-66%.
Result of correlation between AMF population and edapho-climatic factors: Statistical assessment of the relationship between AMF spore population and edapho-climatic factors of the study areas were presented in Table 5. Results revealed very strong significantly positive correlation between AMF spore population and root colonization (r = 0.913**), annual rainfall (r = 0.812**), organic carbon (r = 0.856**). A good correlation was also observed with available nitrogen (r = 0.719*), available potassium (r = 0.724*), pH (r = 0. 702*) and sand content (r = 706*). The relationships between AMF spore population and EC (r = 0.401), silt (r = 0.231), Clay (r = 0.334) and temperature (r = 0.415) were found to be very weak. However, a significantly negative correlation was observed between AMF spore population and phosphorous content (r = - 0.832**).
| Table 4: | AMF species distribution, frequency of occurrence (%) and species richness in the study districts |
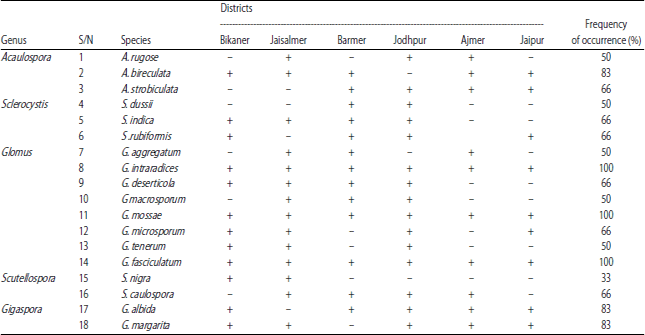 | |
| +: Present, -: Absent | |
| Table 5: | Correlation (r) between AMF spore population and various edaphic and climatic parameters |
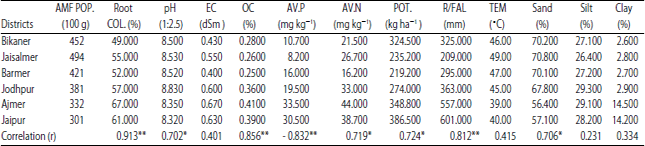 | |
*p<0.05, **p<0.01. AMFPOP: Arbuscular mycorrhizal fungi population, ROOTCOL: Root colonization, EC: Electrical conductivity, OC: Organic carbon, AV.P: Available Phosphorous, AV.N: Available nitrogen, POT: Potash, R/FAL: Rainfall, TEM: Temperature | |
DISCUSSION
The unique ability of plants in the arid zones to release large amount of amino-acids and sugars for AMF growth29, coupled with the low phosphorous content characteristic of arid soils2 might have resulted in higher diversity and AMF spore population in arid districts. High-sand content of all soil samples obtained from arid districts (Table 2) might have provided better aeration for soil humus decomposition and hyphae penetration30 resulting in accelerated fungal propagation and the consequent higher AMF diversity and spore population.
The comparatively higher AMF diversity and spore population recorded in the arid districts (Table 3) might be attributed to the prevalent cultivation practices including intensive irrigation and the use of organic manure. Adequate soil water content had been reported12 to exert obvious impacts on the distribution of AMF communities by improving the physiological status of local AMF and its ecological niche directly, probably because water is essential for reproductive and metabolic processes. In addition, cultural practice of farmyard manure application by the farmers, might have led to increased level of available nutrients, resulting in elevated number of microbial communities9.
The maximum percentage root colonization recorded in the semi-arid districts as compared to the arid districts could be due to a combination of factors prevailing in the semi-arid districts viz; higher organic carbon content, optimum levels of nutrients (available N, P and K), favorable temperature and rainfall regimes6. In addition, Bhat et al.31 confirmed that there are significant interactions between soil available phosphorous, potassium and AMF root infection and colonization. Soil phosphorous had also been reported30,32 to stimulate spore germination, hyphal growth and root infection especially under stress conditions.
Conversely, the lower percentage root colonization recorded in the arid districts might be due to effect of low rainfall and higher temperature. Buenos et al.33 observed that aridity hampered root colonization and reported that in very dry environmental conditions, available water recedes to smaller pores resulting in decreased contact between available spores and water films in the soil. Van der Heidjen et al.34 further reported that lack of soil nutrients inhibited the production and separation of AMF spore for root infection and colonization.
AMF composition and frequency of occurrence: The high species number recorded from the Glomus genus (Table 4) in the study area is similar to previously published research35 that confirmed Glomus species to be the most abundant in the AMF assemblage. This could be ascribed to their unique ability to survive in both acidic and alkaline soils, co-adapt with plants to tolerate environmental challenges and produce excellent inoculum under constraint environmental conditions36.
In addition to its role in carbon allocation, the intermingling and extensive extra-radical mycelium of Glomus species also allows for a more efficient exploitation of soil nutrients and water37, thus benefiting nutrient flow through the soil-fungus-plant system that is particularly relevant in arid ecosystems33,2. Querejeta et al.5 also reported that modulation of leaf gas exchange by native, drought-adapted Glomus species is critical to the long-term performance of host plants in semi-arid environments. In further work, Querejeta et al.38 hypothesized that enhanced transpiration as well as improved plant water status were key mechanisms involved in plant growth stimulation by native Glomus species in the semi-arid soils.
The significantly positive correlation recorded between AMF spore population and organic carbon could be due to the high water and nutrient holding31,34 and buffering capacities of organic carbon39-40. Gerz et al.41 and Bagyaraj and Ashwin42, observed that AM abundance in the soil could be attributed to the availability or otherwise of its nutrient content which in turn, was credited to the amount of OC, N, P and K among others. Positive correlation observed between AMF spore population with available nitrogen and available potassium in Table 5, agrees with the findings of Timer and Den29 and Bhat et al.31 but contradicts the results of Khanam et al.43 who reported a negative correlation between soil K and AMF spore population.
CONCLUSION
The present study concludes that (1) Glomus species are the dominant species in the rhizosphere of tomato (Solanum lycopersicum) in the studied districts, (2) AMF spore population and root colonization were higher in the arid and semi-arid districts respectively and (3) Edapho-climatic factors have great driving influence on the occurrence and distribution of AMF species in the arid and semi-arid regions of Indian desert.
SIGNIFICANCE STATEMENT
This study provides knowledge on the diversity and drivers of AMF in the ecologically stressed ecosystem of arid and semi-arid regions of Indian desert and reveals promising findings that would provide baseline information for a better understanding of more important roles that AMF plays in enhancing resource allocation and increasing tolerance of tomato plant to the various environmental stresses prevalent in this agro-environment. Findings would also provide researchers with fundamental tools for delving into functional profile of AMF and its symbiotic importance with diverse crops in regions with related edapho-climatic conditions.
ACKNOWLEDGMENT
The authors are thankful to the Head of AUSIC laboratory, AIMT, Amity University Rajasthan, Jaipur for her help in the laboratory and to the very friendly farmers in all the study locations who willingly volunteered their farms for sample collection.
REFERENCES
- Kamalvanshi, M., A. Kumar, A. Jha and S.K. Dhyani, 2012. Occurrence of arbuscular mycorrhizal fungi in rhizosphere of Jatropha curcas L. in arid and semi arid regions of India. Indian J. Microbiol. 52: 492-494.
CrossRefDirect Link - Anthony, B. and R. Loren, 2006. Genetic features of population from stress-prone environments. Enhancing the Use of Crop Genetic Diversity to Manage Abiotic Stress in Agricultural Production Systems. May 23-27, 2005, International Plant Genetic Resources Institute, pp: 1-10.
Direct Link - Ansari, FA. and I. Ahmad, 2018. Plant growth promoting attributes and alleviation of salinity stress to wheat by biofilm forming Brevibacterium sp. FAB3 isolated from rhizospheric soil. Saudi J. Bio. Sci.
CrossRefDirect Link - Querejeta, J.I., M.F. Allen, F. Caravaca and A. Roldan, 2006. Differential modulation of host plant δ13C and δ18O by native and nonnative arbuscular mycorrhizal fungi in a semiarid environment. New Phytol., 169: 379-387.
CrossRefDirect Link - Osakabe, Y., K. Osakabe, K. Shinozaki and L.S.P. Tran, 2014. Response of plants to water stress. Front. Plant Sci., Vol. 5.
CrossRefDirect Link - Savvides, A., S. Ali, M. Tester and V. Fotopoulos, 2016. Chemical priming against multiple abiotic stresses: Mission possible? Trend Plant Sci., 21: 329-340.
CrossRefDirect Link - Coleman-Derr, D. and S.G. Tringe, 2014. Building the crops of tomorrow: advantages of symbiont-based approaches to improving abiotic stress tolerance. Front. Microbiol., 5: 283-290.
CrossRefDirect Link - Auge, R.M., 2001. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza, 11: 3-42.
CrossRefDirect Link - Augé, R.M., H.D. Toler and A.M. Saxton, 2015. Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: a meta-analysis. Mycorrhiza, 25: 13-24.
CrossRefDirect Link - Krasensky, J. and C. Jonak, 2012. Drought, salt and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot., 63: 1593-1608.
CrossRef - Chitarra, W., C. Pagliarani, B. Maserti, E. Lumini and I. Siciliano et al., 2016. Insights on the impact of arbuscular mycorrhizal symbiosis on tomato tolerance to water stress. Plant Physiol., 71: 1009-1023.
CrossRefDirect Link - Porcel, R., R. Aroca and J.M. Ruiz-Lozano, 2012. Salinity stress alleviation using arbuscular mycorrhizal fungi: A review. Agron. Sustainable Dev., 32: 181-200.
CrossRefDirect Link - McGonigle, T.P., M.H. Miller, D.G. Evans, G.L. Fairchild and J.A. Swan, 1990. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol., 115: 495-501.
CrossRefDirect Link - Stutz, J.C., and J.B. Morton, 1996. Successive pot cultures reveal high species richness of Arbuscular endomycorrhizal fungi in arid ecosystems. Canadian J. Bot., 74: 1883-1889.
CrossRefDirect Link - Gerdemann, J.W. and T.H. Nicolson, 1963. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc., 46: 235-244.
CrossRefDirect Link - Khade, S.W. and B.F. Rodrigues, 2008. Spatial variations of AM fungi in Terminalia crenulata Roth from the western Ghat region of Goa, India. Mycorrhiza News, 20: 20-21.
Direct Link - Morton, J.B. and G.L. Benny, 1990. Revised classification of arbuscular mycorrhizal fungi (Zygomycetes): A new order, Glomales, two new suborders, Glomineae and Gigasporineae and two new families, Acaulosporaceae and Gigasporaceae, with an emendation of Glomaceae. Mycotaxon, 37: 471-491.
Direct Link - Xiao, E., V. Krumins, S. Tang, T. Xiao, Z. Ning, X. Lan, W. Sun, 2016. Correlating microbial community profiles with geochemical conditions in a watershed heavily contaminated by an antimony tailing pond. Environ. Poll., 215: 141-153.
CrossRefDirect Link - Edgar, R.C., 2013. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods, 10: 996-998.
CrossRefDirect Link - Schloss, P.D., S.L. Westcott, T. Ryabin, J.R. Hall and M. Hartmann, 2009. Introducing mothur: open-source, platform independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol., 75: 7537-7541.
CrossRefDirect Link - Dufrene, M. and P. Legendre, 1997. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monographs, 67: 345-366.
CrossRefDirect Link - Timer, L.W. and R.F.L. Den, 1980. The relationship of mycorrhizal infection to phosphorous-Induced copper deficiency in sour orange seedlings. New Phytol., 85: 15-23.
Direct Link - Torrecillas, E., M.D.M. Alguacil, A. Roldán, G. Díaz, A. Montesinosnavarro and M.P. Torres, 2014. Modularity reveals the tendency of arbuscular mycorrhizal fungi to interact differently with generalist and specialist plant species in gypsum soils. Appl. Environ. Microbiol., 80: 5455-5466.
CrossRefDirect Link - Bhat, B.A., S. Muzamilhamad and T. Avinash, 2014. Impact of various edaphic factors on AMF spore population and diversity in Catharanthus roseus at Gwalior, India. Int. J. Plant Sci., 9: 1-6.
Direct Link - Zhang, H., S. Wei, W. Hu, L. Xiao and M. Tang, 2017. Arbuscular mycorrhizal fungus Rhizophagus irregularis increased potassium content and expression of genes encoding potassium channels in Lycium barbarum. Front. Plant Sci., Vol. 8.
CrossRefDirect Link - Bueno, C.G., M. Moora, M. Gerz, J. Davison, M. Öpik et al., 2017. Plant mycorrhizal status, but not type, shifts with latitude and elevation in Europe. Global Ecol. Biogeogr., 26: 690-699.
CrossRefDirect Link - Der Heijden, V., M.G., F.M. Martin, M.A. Selosse and I.R. Sanders, 2015. Mycorrhizal ecology and evolution: The past, the present and the future. New Phytol., 205: 1406-1423.
CrossRefDirect Link - Gerz, M., C.G. Bueno, M. Zobeland M. Moora, 2016. Plant community mycorrhization in temperate forests and grasslands: Relations with edaphic properties and plant diversity. J. Veg. Sci., 27: 89-99.
CrossRefDirect Link - Soudzilovskaia, N.A., J.C. Douma, A.A. Akhmetzhanova, P.M. van Bodegom, W.K Cornwell et al., 2015. Global patterns of plant root colonization intensity by mycorrhizal fungi explained by climate and soil chemistry. Global Ecol. Biogeogr., 24: 371-382. https://doi.org/10.1111/geb.12272-382.
CrossRefDirect Link - Bennett, J.A., H. Maherali, K.O. Reinhart, Y. Lekberg, M.M. Hart and J.N. Klironomos, 2017. Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science, 355: 181-184.
CrossRefDirect Link - Querejeta, J.I., M.F. Allen, M.M. Alguacil and A. Roldan, 2007. Plant isotopic composition provides insight into mechanisms underlying growth stimulation by AM fungi in a semiarid environment. Funct. Plant Biol., 34: 683-691.
CrossRefDirect Link - Cheng, L., F.L. Booker, C. Tu, K.O. Burkey and L. Zhou et al., 2012. Arbuscular mycorrhizal fungi increase carbon decomposition under elevated C02. Science, 337: 1084-1087.
CrossRefDirect Link - Bonfim, J.A., R.L.F. Vasconcellos, T. Gumiere, D.D.L.C. Mescolotti, F. Oehl and E.J.B.N. Cardoso, 2016. Diversity of arbuscular mycorrhizal fungi in a Brazilian Atlantic forest Toposequence. Microbiol. Ecol., 71: 164-172.
CrossRefDirect Link - Gerz, M., B.C. Guillermo, W.A. Ozinga, M. Zobeland M. Moora, 2018. Niche differentiation and expansion of plant species are associated with mycorrhizal symbiosis. J. Ecol., 106: 254-264.
CrossRefDirect Link - Khanam, D., M.A.U. Mridha, A.R.M. Solaiman and T. Hossain, 2006. Effect of edaphic factors on root colonization and spore population of arbuscular mycorrhizal fungi. Bull. Inst. Trop. Agric. Kyushu Univ., 29: 97-104.
Direct Link