
Clement Wong Kiing Fook
Intitute of Tropical Agriculture, Universiti Putra Malaysia, Serdang, Selangor, Malaysia
Lai Kok Song
Department of Cell and Molecular Biology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, Serdang, Selangor, Malaysia
Wong Mui Yun
Department of Plant Pathology, Faculty of Agriculture, Universiti Putra Malaysia, Serdang, Selangor, Malaysia
Maziah Mahmood
Department of Biochemistry, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, Serdang, Selangor, Malaysia
Asian Journal of Agricultural Research
Year: 2016 | Volume: 10 | Issue: 2 | Page No.: 126-131







ABSTRACT
Determining the suitable and optimized Minimal Inhibitory Concentration (MIC) of the selectable agents is critical to successfully recover stably transformed plants. In this study, the MIC of five different antibiotics and one herbicide was determined for their inhibitory effect on the growth of a recalcitrant rice cultivar, MR219 using rice shoot apices as target tissue. Antibiotics such as kanamycin, paromomycin and neomycin did not inhibit the shoot apices growth while geneticin (G–418) and hygromycin showed complete growth inhibition at 400 and 25 mg L–1, respectively. Similarly, the herbicide basta indicated that 9 mg L–1 was sufficient to cause growth inhibition. To conclude, hygromycin and basta were suggested to the suitable selectable agent to select transformed rice shoot apices.
PDF Abstract XML References Citation
Received: December 16, 2015;
Accepted: January 21, 2016;
Published: February 15, 2016
How to cite this article
Clement Wong Kiing Fook, Lai Kok Song, Wong Mui Yun and Maziah Mahmood, 2016. Minimal Inhibitory Concentration of Common Selectable Agents on Recalcitrant Malaysian Rice Cultivar. Asian Journal of Agricultural Research, 10: 126-131.
DOI: 10.3923/ajar.2016.126.131
URL: https://scialert.net/abstract/?doi=ajar.2016.126.131
DOI: 10.3923/ajar.2016.126.131
URL: https://scialert.net/abstract/?doi=ajar.2016.126.131
INTRODUCTION
More than 90% of rice production accounts for the daily diet of more than half of the world’s population, particularly those from the Asian region. Like many other staple crops around the world, rice is susceptible to various pest, diseases and environmental changes which in turn, hampers the rice production sector leading to food security crisis (Beddington, 2010). In addition, decreasing arable land for farming and booming human population have further impeded rice production (Jiang et al., 2013). All these factors have driven the need to look for techniques to boost rice yield. Genetic engineering was sought out as one of the biotechnological alternative that has enabled researchers to insert genes from wide array of organisms in order to produce transgenic rice with higher yield, improved pest and disease resistance and improved resilience towards climate change (Shew et al., 2015).
A successful genetic transformation event is largely dependent on an effective and efficient selection system. Transformed plant cells can be selected through negative selection in which untransformed cells are killed or inhibited. Conversely, only transformed cells containing the expressed selectable marker gene will confer resistance to the toxic products by means of detoxification (Moazez et al., 2013). For efficient use of selectable agents, it is essential to determine the MIC of the agent needed to eliminate the untransformed cells because lower concentrations will produce chimeras or escapees whereas higher concentration will inhibit transformed cells (Breyer et al., 2014). So far, the reported selectable agents that have been utilized to select transgenic rice shoot apices were kanamycin (Dey et al., 2012), hygromycin (Yookongkaew et al., 2007) and basta (Park et al., 1996). However, the systematic analysis of the MIC of these and other common selectable agents towards the recalcitrant Malaysian indica rice has not been reported. Moreover, the MIC of the selectable agents has been reported to be tissue and species specific (Htwe et al., 2014) which suggested that proper selection and optimizing the MIC is therefore crucial for genetically transformed cells to proliferate under selection pressure whilst eliminating the possibility of non-transformed cells (escapees) and chimeras. Henceforth, this study sought to systematically optimize the MIC of the selectable agents in the growth inhibition of shoot apices. Ultimately, this study will serve as a guide in the selection of suitable selectable agents for rice transformation studies.
MATERIALS AND METHODS
Plant materials: This study was carried out from 2014-2015, from October to February, in Plant Biotechnology Laboratory, UPM, Malaysia. The Malaysian rice cultivar MR219 was used throughout the study. Shoot apices derived from in vitro 4 days old rice seedlings germinated from mature seeds were used as explants.
Preparation of selectable agents: Five types of aminoglysoside antibiotics, namely kanamycin, neomycin, paromomycin, G-418 and hygromycin (Merck, Germany) and the herbicide, basta (Bayer CropScience, Malaysia) were selected for this study since they were commonly used in rice genetic transformation. All selectable agents were prepared as 50 mg mL–1 of stock solution and filter sterilized with 0.22 μM MCE syringe filter (Merck Millipore, Germany) prior to adding into cooled (50°C) autoclaved MS solid media (Murashige and Skoog, 1962).
Determination of MIC for selectable agents: Approximately 3 mm of rice shoot apices were prepared by separating the roots, endosperm and shoot tips from four days old in vitro germinated rice seedlings. Shoot apices were subjected to in vitro antibiotic and herbicide sensitivity screening at a concentration range from 1-500 mg L–1 (Table 1). The MS media without the addition of selectable agents served as the positive control. five replications with 10 explants per replicate were carried out for each screening. After two weeks of in vitro culturing, the percentage of survived explants was determined based based on the green and proliferating shoots. The MIC was determined based on the minimum concentration of each selectable agents that completely inhibited or killed the shoot apices whereby necrotic and inhibited shoot growth were observed. All experiments were repeated three times and mean values from each data set was analyzed and compared by Duncan’s multiple range test at 5% (p≤0.05).
RESULTS AND DISCUSSION
Aminoglycosides function to bind and inactivate chloroplastic ribosomes in plants and therefore, causing death in plants (Mentewab et al., 2014). This was not the case for certain aminoglycosides in this study. At all tested concentrations of kanamycin, neomycin and paromomycin, 100% of survival of shoot apices was recorded (Fig. 1a-c).
| Table 1: | Percentage survival of rice shoot apices based on 5 replicates consisting of 10 shoot apices per replicate |
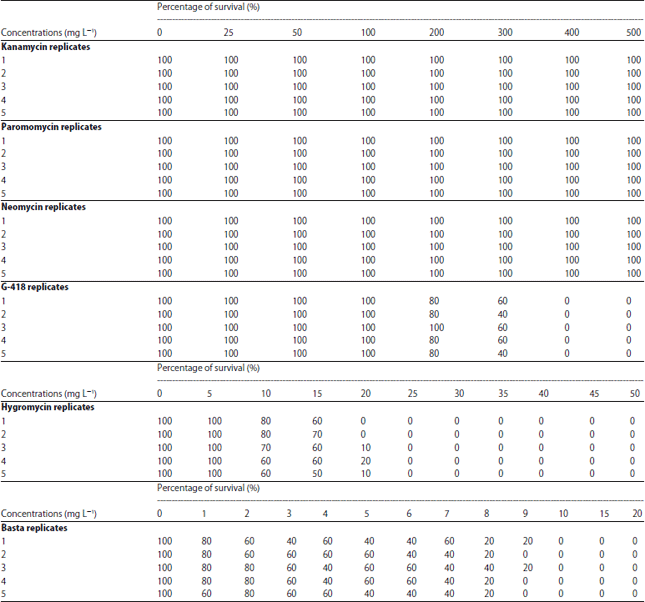 | |
Furthermore, newly produced albino plants from shoot apices were observed in kanamycin media at concentration from 200-500 mg L–1 (Fig. 2b) and such phenomenon was also observed in other studies as well (Safitri et al., 2013). It was postulated that kanamycin enters into the chloroplasts via specific transporters that disrupt chlorophyll synthesis thereby producing albino or chlorotic plants (Conte et al., 2009). So far, there are no reports on the utilization of paromomycin and neomycin in transgenic rice studies. Though found ineffective towards rice shoot apices at all tested concentrations, neomycin and paromomycin were proven to be superior in selection studies of other monocots which includes orchid (Gnasekaran et al., 2014) and maize (Que et al., 2014). Nevertheless, both were reported to be inefficient as selectable agents for rice shoot apices as demonstrated in this study (Fig. 1b, c). The insensitivity of shoot apices towards these three aminogylcosides was probably due to the presence of endogenous and non-specific phospho transferases activity in rice resulting in conferred antibiotic resistance (Mentewab et al., 2014). In addition, the higher cell wall composition in the shoot apices used in this study might have prevented the diffusion of the antibiotics into the host cells (Mentewab et al., 2014).
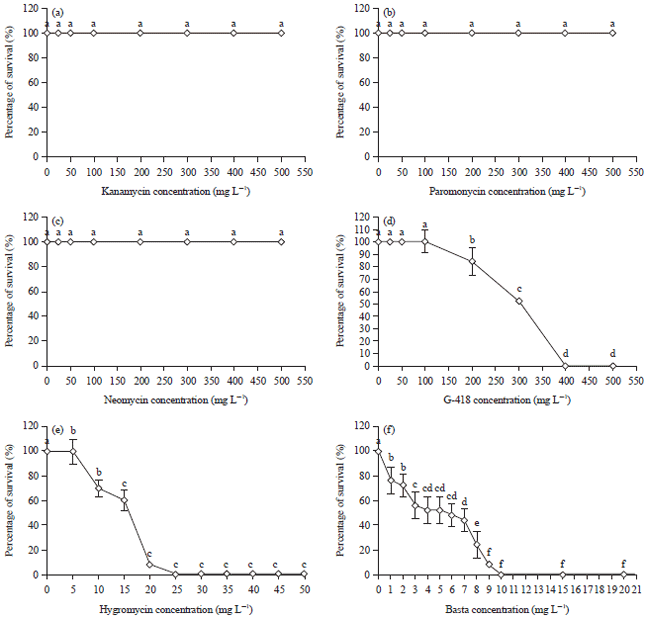 | |
| Fig. 1(a-f): | Percentage of survival of shoot apices after two weeks of culturing in varying concentrations of (a) Kanamycin, (b) Paromomycin, (c) Neomycin, (d) G-418, (e) Hygromycin and (f) Basta. Different letters indicate the values are statistically different (p≤0.05) |
In contrast, G-418 was found to be more toxic to rice cells in which the percentage of survival dropped from 52% at 300 mg L–1 to 0% at 400 mg L–1 (Fig. 1d and 2c). The G-418 was reported to contain a hydroxyl functional group at the carbon-6 in place of the amino group (NH2) which enhances its toxicity towards cells as compared to other aminoglycosides (Padilla and Burgos, 2010).
Signs of decolorized shoots, inhibited growth and even shoot death was observed in MS media supplemented with hygromycin and basta (Fig. 2d, e). For hygromycin, the survival percentage was reduced to 8.5% at 20 mg L–1 and growth was completely inhibited from 25 mg L–1 onwards showing necrotic explants (Fig. 1e). Interestingly, at higher concentrations of hygromycin (45 and 50 mg L–1), explants showed yellowing and inhibited growth instead of necrosis (Fig. 2d). There are no literatures explaining such observation. Such phenomenon might require further investigation. Comparing to G-418, rice cells are more sensitive to hygromycin at lower concentrations as evidenced in previous studies (Sultana et al., 2014; Oung et al., 2015). Unlike kanamycin, hygromycin was effectively used to produce albino-free and fertile rice transformants. The efficiency of basta was also comparable to hygromycin and G-418. Basta, a common herbicide containing the active ingredient phosphinothricin, inhibits the glutamine synthetase which directly interfere with the nitrogen assimilation in plants.
 | |
| Fig. 2(a-e): | Represents the shoot growth at different concentrations of (a) MSO (positive control), (b) Kanamycin, (c) G-418, (d) Hygromycin and (e) Basta. *MIC of each selectable agents, **Albino plants being produced, ***Necrotic explants, ****Yellowing explants, bar represents 1 cm |
At 10 mg L–1 of basta, complete death of explants was observed (Fig. 1f) which was in correspondence to previous study suggesting that effective concentration for basta was from 0.5-10 mg L–1 (Dedicova et al., 2015).
CONCLUSION
To conclude, amongst the selectable agents tested, G-418, hygromycin and basta were suitable to be used to select transformed rice. Nonetheless, the MIC determined for hygromycin (25 mg L–1) and basta (10 mg L–1) were significantly lower than G-418 (300 mg L–1) which might suggest that both hygromycin and basta can be used efficiently and economically at lower concentrations. This study has provided a systematic MIC determination of commonly used selectable agents for recalcitrant rice MR219. This serves as an essential fundamental and guide for researchers to make proper selection to ensure a successful rice transformation event.
ACKNOWLEDGMENTS
Special thanks to LRGS food security, Enhancing Sustainable Rice Production through Innovative Research (Grant No. 5525014) and UPM-GRF for the research funding and scholarship.
REFERENCES
- Beddington, J., 2010. Food security: Contributions from science to a new and greener revolution. Philosophical Trans. Royal Soc. B: Biol. Sci., 365: 61-71.
CrossRefDirect Link - Breyer, D., L. Kopertekh and D. Reheul, 2014. Alternatives to antibiotic resistance marker genes for in vitro selection of genetically modified plants-scientific developments, current use, operational access and biosafety considerations. Crit. Rev. Plant Sci., 33: 286-330.
CrossRefDirect Link - Conte, S., D. Stevenson, I. Furner and A. Lloyd, 2009. Multiple antibiotic resistance in Arabidopsis is conferred by mutations in a chloroplast-localized transport protein. Plant Physiol., 151: 559-573.
CrossRefDirect Link - Dedicova, B., C. Bermudez, M. Prias, E. Zuniga and C. Brondani, 2015. High throughput transformation pipeline for a Brazilian japonica rice with bar gene selection. Protoplasma, 252: 1071-1083.
CrossRefDirect Link - Dey, M., S. Bakshi, G. Galiba, L. Sahoo and S.K. Panda, 2012. Development of a genotype independent and transformation amenable regeneration system from shoot apex in rice (Oryza sativa spp. indica) using TDZ. 3 Biotech, 2: 233-240.
CrossRefDirect Link - Gnasekaran, P., J.J.J. Antony, J. Uddain and S. Subramaniam, 2014. Agrobacterium-mediated transformation of the recalcitrant Vanda Kasem's delight orchid with higher efficiency. Scient. World J.
CrossRefDirect Link - Htwe, N.N., H.C. Ling, F.Q. Zaman and M. Maziah, 2014. Plant genetic transformation efficiency of selected malaysian rice based on selectable marker gene (hptII). Pak. J. Biol. Sci., 17: 472-481.
CrossRefDirect Link - Jiang, L., X. Deng and K.C. Seto, 2013. The impact of urban expansion on agricultural land use intensity in China. Land Use Policy, 35: 33-39.
CrossRefDirect Link - Mentewab, A., K. Matheson, M. Adebiyi, S. Robinson and B. Elston, 2014. RNA-Seq analysis of the effect of kanamycin and the ABC transporter AtWBC19 on Arabidopsis thaliana seedlings reveals changes in metal content. PLOS One, Vol. 9, No. 10.
CrossRefDirect Link - Moazez, Y., H.R. Memari, H. Ofoghi, M. Roayaei and D.N. Ahmadi, 2013. Evaluation of Spirulina platensis resistance to different antibiotics to find a selectable marker for genetic transformation. Jundishapur J. Microbiol., Vol. 6, No. 7.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Oung, H.M., K.C. Lin, T.M. Wu, N.N.P. Chandrika and C.Y. Hong, 2015. Hygromycin B-induced cell death is partly mediated by reactive oxygen species in rice (Oryza sativa L.). Plant Mol. Biol., 89: 577-588.
CrossRefDirect Link - Padilla, I.M.G. and L. Burgos, 2010. Aminoglycoside antibiotics: Structure, functions and effects on in vitro plant culture and genetic transformation protocols. Plant Cell Reports, 29: 1203-1213.
CrossRefDirect Link - Park, S.H., S.R.M. Pinson and R.H. Smith, 1996. T-DNA integration into genomic DNA of rice following Agrobacterium inoculation of isolated shoot apices. Plant Mol. Biol., 32: 1135-1148.
CrossRefPubMedDirect Link - Que, Q., S. Elumalai, X. Li, H. Zhong and S. Nalapalli et al., 2014. Maize transformation technology development for commercial event generation. Front. Plant Sci., Vol. 5.
CrossRefDirect Link - Safitri, F.A., M. Ubaidillah and K.M. Min, 2013. Response of explants of calli rice (Oryza sativa L.) Japonica cv. Am. J. Plant Sci., 4: 838-843.
CrossRefDirect Link - Shew, A.M., L.L. Nalley, D.M. Danforth, B.L. Dixon, R.M. Nayga Jr., A.C. Delwaide and B. Valent, 2015. Are all GMO's the same? Consumer acceptance of cisgenic rice in India. Plant Biotechnol. J., 14: 4-7.
CrossRefDirect Link - Sultana, S., C.L. Ho, P. Namasivayam and S. Napis, 2014. Genotypic differences in response to Hygromycin effect on untransformed calli death and rice germination. Bangladesh Rice J., 18: 38-43.
Direct Link - Yookongkaew, N., M. Srivatanakul and J. Narangajavana, 2007. Development of genotype-independent regeneration system for transformation of rice (Oryza sativa ssp. indica). J. Plant Res., 120: 237-245.
CrossRefDirect Link