
Tewodros Mulualem
Jimma Agricultural Research Center, P.O. Box 192, Jimma, Ethiopia
Zelalem Bekeko
Haramaya University, School of Plant Sciences, P.O. Box 138, Dire Dawa, Ethiopia
Asian Journal of Agricultural Research
Year: 2015 | Volume: 9 | Issue: 2 | Page No.: 38-54







ABSTRACT
The study is intended for highlighting the physiological mechanisms associated with breeding of cassava towards moisture stress tolerance. It describes important points including: The special effects of moisture stress, mechanisms of adaptation of crops to moisture stress, moisture stress and improvement on cassava, reproductive biology of cassava, current breeding goal, methods of screening and breeding of cassava for moisture stress environment, current achievement on conventional breeding on cassava and physiological response of cassava in moisture stress environment. The information summarized in this study may serve as important guideline in cassava improvement towards moisture stress tolerance.
PDF Abstract XML References Citation
Received: January 12, 2015;
Accepted: March 09, 2015;
Published: April 07, 2015
How to cite this article
Tewodros Mulualem and Zelalem Bekeko, 2015. Assessment of Conventional Breeding on Cassava and its Physiological Adaptive
Mechanisms: Implication for Moisture Stress. Asian Journal of Agricultural Research, 9: 38-54.
DOI: 10.3923/ajar.2015.38.54
URL: https://scialert.net/abstract/?doi=ajar.2015.38.54
DOI: 10.3923/ajar.2015.38.54
URL: https://scialert.net/abstract/?doi=ajar.2015.38.54
INTRODUCTION
Cassava (Manihot esculenta Crantz) is a tropical woody shrub. It is dicotyledonous plant belonging to the botanical family Euphorbiaceae (Onwueme and Charles, 1994). It is originated from North East Brazil with an additional centre of origin in Central America (Nassar, 2002). Though the domestication of cassava started from these areas, it cultivated in tropical and subtropical areas of the world (Elias et al., 2001). Edible roots from this crop are usually harvested annually.
Cassava is being cultivated as the main source of energy and as the most important staple food crop for over 900 million of people in the world (Cock, 1985; Beyene, 2012). It is the second most important staple crop in Africa after maize. It is the 6th most consumed crops in the world (Benesi et al., 2004). The world production of cassava amounts to about 225 Mmt per year, of which 121 Mmt comes from tropical Africa (FAO., 2006).
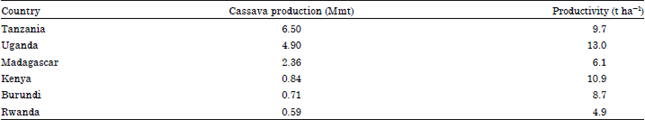
Nigeria is the world’s largest producer of cassava, accounting for over 18.67% of the world total output. In 2006, Nigeria alone produced 42 million tones of cassava representing 34.71% of cassava produced in Africa (FAO., 2006). The other top cassava producer countries are Tanzania (6.5 Mmt), Uganda (4.93 Mmt), Madagascar (2.36 Mmt), Kenya (0.84 Mmt), Burundi (0.71 Mmt) and Rwanda (0.59 Mmt) in 2005 and 2006 (Table 1). Cassava was first introduced to Ethiopia by the British. Although reliable statistical information on the area and production of cassava in Ethiopia is deficient, the crop has been cultivated, mainly, in the South, South west and Western parts of the country since its introduction. Its use as a potential food crop in Ethiopia has increased during and after the 1984 famine (Beyene, 2012). The crop is well known in its principal ability to produce economic yields under relatively marginal rainfall and soil conditions.
| Table 1: | Total annual production of cassava for the period from 2005-2006 in East Africa (Source of data: FAO (2006)) |
 | |
The consumption pattern varies within countries by regions and by income of the population group. Humans can consume cassava roots, young leaves and the tips of stems as vegetables. Most rural poor consumes boiled or baked cassava roots while people with more economic means tend to eat cassava roots as fried chips or as a snack food (Woolfe, 1992; Subbarao et al., 2005).
Apart from food, cassava is very versatile and its derivatives and starch are applicable in many types of products such as, confectionery, sweeteners, glues, plywood, textiles, paper, biodegradable products, monosodium glutamate and drugs. Cassava chips and pellets are used in animal feed and alcohol production. According to Kapinga et al. (1998), there is an increased interest in using high quality cassava flour as a substitute for cassava flour in food and non-food product industries (Ekanayake et al., 1997).
Cassava can be grown in adverse environments and be able to yield remarkably. Despite its ability to grow under sub-optimal conditions, its high yield in small area of land, multipurpose advantage and importance as a security crop in densely populated area, the cultivation and consumption of this potential crop in Africa including in Ethiopia is almost negligible. This could be attributed presence of knowledge gaps on breeding techniques to select best materials and so as to reduce its toxic compound, lack of adaptable, resistant technologies for various stresses, postharvest-handling and processing technique are a few to mention. Therefore, this review study is initiated to assess the conventional breeding on cassava to get desirable traits for moisture stress and high yield and quality. The information so derived could be exploited in devising further production strategies and to develop new breeding techniques on improving cassava for moisture stress environment (Janssens, 2001).
ECONOMIC IMPORTANCE OF CASSAVA
Cassava accounts for approximately one-third of the total staple food production and provides over 50% of the energy for more than 300 million people in sub-Saharan Africa. In Central Africa alone, cassava is estimated to provide over 1000 kcal day-1 to 30 million people (Cock, 1985). The storage roots form the basic carbohydrate component of the diet and the leaves which contain appreciable amounts of vitamins, minerals and proteins are consumed as a preferred green vegetable in many parts of Africa, providing protein, min erals and vitamins (Hahn et al., 1990). The crop is grown almost exclusively as food in many tropical areas of Africa where rainfall exceeds 600 mm over a period of at least 2-3 months and altitudes range from sea level to 1800 m. It is particularly important in those areas where food supply is constantly threatened by environmental constraints such as moisture stress and pest outbreaks, because of its ability to grow under conditions considered as suboptimal for the majority of food crops. It can be harvested any time from 6-24 months after planting and can be left in the ground as a food reserve for household food security in times of famine, drought and war. According to Global cassava production project the total cassava yield in 2004 to be around 192 million tones, as was recorded in 2003 with Africa contributing 103 million.
In spite of the economic value of cassava, its production is limited by moisture stress in many tropical regions when the crops became an early stage. The yield reduction due to moisture stress was estimated at 80%. Under field experiment, it was observed that moisture stress for the first 30 days during at early growing stage decreased yield by 65-80% of cassava (Beyene, 2012). Application of irrigation during the early stage is an ideal and practical solution to overcome moisture stress in crop production. However, farmers do not have access to irrigation water and infrastructures. Moreover, allocation of clean water for irrigation is a big challenge because of exponential increase of population and the current global climate change. Consequently, the sustainable solution to improve cassava production is to develop and deploy moisture stress tolerance varieties. Breeding for moisture stress tolerance requires knowledge on the physiological mechanisms involved in moisture stress tolerance and the genetic control of yield and its components. Molecular breeding techniques may improve the response of selection to moisture stress tolerance. However, their efficiency greatly depends on the availability of linked physiological and morphological traits (Subbarao et al., 2005).
EFFECT OF DROUGHT
Drought is in adequacy of water availability, including precipitation and soil moisture storage capacity, in quantity and distribution during the life cycle of the plant. In drought conditions, a water potential (Ψw) of soil becomes negative because of a concentration increase of soil solutes. The movement of cell water is determined by the water potential gradient. Drought stress requires changes in plant cells and tissues to adapt to drought stress condition and continue to acquire little available water of soil (Bartels and Sunkar, 2005). The limited amount and uneven distribution of water affect growth and development of the crops. Eventually this leads food shortage in many semi-arid countries in general and developing countries in particular (Acquaah, 2007). Drought can cause the biggest loss in crop production compared to other abiotic stress factors (Ober, 2008). It affects crop production by reducing the genetic potential of a plant (Mitra, 2001). Consequently, it is responsible of the difference between the mean yield and the yield potential of a crop and the cause of the yield instability in time (Sorrels et al., 2000).
Drought affects the root distribution of the plant in the soil. Soil moisture deficit rarely increase root biomass but root to shoot dry matter ratio tends to increase. The relative biomass distribution in the root system tends to change in favour of deeper rooting at the expense of shallow rooting. As soil dries up from the top roots at the top die while existing live roots grow into depth. There are some indications that different parts of the root respond differently to soil drying. For example lateral root growth of maize and wheat was promoted with the development of moderate water deficit but not with severe deficit (Ito et al., 2006). Lateral root growth is crucial in dry land row crops; whereas it determines the capacity for extraction of soil moisture in the spacing between rows.
The breeder must be reminded of the fact that beyond soil moisture a host of other factors can impact and hinder root growth and function. These include biotic factors such as soil inhibiting pathogens, insects and nematodes as well as abiotic factors such as soil salinity, alkalinity, acidity, water logging and mineral toxicity.
MOISTURE STRESS RESISTANCE AND ITS IMPROVEMENT
Plant breeding has been successful in developing moisture stress resistant crop cultivars. However, the traditional breeding method by using yield as a selection index and performing multi-environmental yield trials has been costly and slow. Plant physiology is now incorporated into the breeding program by using physiological selection criteria relevant to the designated plant ideotype and subsequent plant performance in the target stress environment (Venuprasad et al., 2009). Genomics offer a great potential for the improvement of breeding efficiency towards water limited environments. There are still inherent problems in deploying marker assisted selection and transgenic technology into breeding program for moisture stress resistance. The potential of genomics can be realized only when it will be well synchronized with plant breeding concept, theory and methods. It has often been voiced and published that “Moisture stress resistance” is complex and therefore its improvement is difficult (Venuprasad et al., 2009).
Breeding for moisture stress resistance can basically follow an analogy of breeding for disease resistance in terms of concept and design (with few exceptions). Moisture stress resistance is approached in terms of its components, namely dehydration avoidance, dehydration tolerance and moisture stress escape. The most widespread and effective mechanism of moisture stress resistance in crop plants is dehydration avoidance which is the ability of the plant to maintain its hydration. It is controlled by plant constitutive traits and plant adaptive traits. Dehydration tolerance which is the ability to function in a dehydrated state is rare but can sometimes be important. It is shown that when stress physiology, plant genetics and knowledge of the target environment are combined it is possible to design an appropriate plant ideotype to be used as guide in breeding for the specific water limited environment (Nguyen et al., 1997).
MECHANISMS OF ADAPTATION TO DROUGHT STRESS
The effect of moisture stress is similar to drought stress; therefore it is better to describe the mechanism of adaptation as drought stress. There are four different mechanisms which help in survival of plants under moisture deficit conditions these are: Drought escape, drought avoidance, drought tolerance and moisture stress resistance.
Drought escape: The simplest way of survival under drought conditions is to escape drought. Generally, drought occurs either in the mid or late crop season. Drought escape is most common in case of plants grown in desert regions. They complete their life cycles before the onset of drought. Drought escape also plays an important role in some crop plants. For Example, yields of early varieties of crops are less affected by severe drought than late maturing ones. All these crops have determinate growth habit. In most cases, late maturing varieties give higher yield than early types especially when drought occurs early in the season.
Drought avoidance: Drought avoidance refers to ability of the plant to maintain a favourable internal water balance under moisture stress. In other words, plants which avoid drought retain high water contents in their tissues. Drought avoidance can permit a longer growth period in the crop through reduced water use or increased water uptake. However, drought avoidance leads to reduction in photosynthesis and thereby reduction in the growth of aerial parts. It leads to increase in root development and therefore, is more important than drought tolerance. In cereals, drought avoidance operates during vegetative phase while tolerance operates during reproductive phase. Drought avoidance mechanisms are of two types. First those which reduce water loss through transpiration. Such features include stomatal characteristics and shape, size and orientation of leaves. The second, those which maintain water uptake during drought period.
Drought tolerance: The ability of crop plants to withstand low tissue water content is referred to as drought tolerance. Drought tolerance is more desirable because the crop can produce more yield at lower water potential. In cereals, drought tolerance generally operates during reproductive phase. Tolerant cultivars exhibit better germination, seedling growth and photosynthesis. In sorghum, a drought resistant line exhibited higher photosynthetic rate at leaf water potential than a less drought resistant line. Drought tolerance differs from drought avoidance in several aspects.
Drought resistance: Drought resistance is the sum of drought avoidance and drought tolerance. In other words, drought resistance refers to the ability of crop plants to give good yield under moisture deficit conditions. Drought resistance is measured in terms of various mechanisms associated with drought tolerance and yield under soil moisture deficit. In winter wheat, both avoidance and tolerance features are important for drought resistance.
Recovery: Because drought varies in duration, some species are able to rebound (recover) after a brief drought episode. Traits that enhance recovery from drought include vegetative vigor, tillering and long growth duration.
Reproductive biology of cassava: Commercial propagation of cassava is by stem cuttings. However, sexual reproduction is a key element for conventional breeding, is common and relatively easy to achieve (Alves, 2002). Most breeding programs generate new genetic variation through crossing. Controlled pollinations generate full-sib progenies. Alternatively, in polycross nurseries insects do the pollinations. In this case the exact origin of the pollen is not known and half-sib families are produced. A certain proportion of seed from polycross nurseries may be the result of self-pollinations. Occasionally botanical seed has been used for commercial propagation (Rajendran et al., 2000) but it is not a generalized practice. Propagation from true seed occurs occasionally in farmers’ fields and as such, is also the starting point for the generation of useful genetic diversity (Elias et al., 2001; Pujol et al., 2002). Efficient farmers have been found to be key players in this informal genetic improvement process (Sambatti et al., 2001).
Cassava is a monoecious species, with female flowers opening 10-14 days before the male flowers on the same branch. Self-pollination can occur because male and female flowers on different branches or on different plants of the same genotypes can open simultaneously (Jennings and Iglesias, 2002). Flowering depends on the genotype and environmental conditions. Branching occurs when an inflorescence is formed. Because erect, non-branching types are frequently preferred by farmers, the crossing of elite clones in certain regions may become more difficult because of the scarcity of their flowers.
Synchronization of flowering remains a difficult issue in cassava breeding. Some clones flower relatively early at 4 or 5 Months After Planting (MAP), whereas others flower only at 8-10 MAP. Because of this and the time required for the seed to mature, it takes generally more than a year to obtain seeds of a planned cross. On average, between one and two seeds (out of the three possible formed in the trilocular fruit) per pollination are obtained. Several publications illustrate the procedures for controlled pollinations in cassava (Jennings and Iglesias, 2002). Seeds often have a dormancy period of a few months after maturity and they require relatively high temperatures (30-35°C) for optimum germination (Elias et al., 2001). There is no evidence of incompatibility, so crosses can be done easily (except for the scarcity or absence of flowers in certain genotypes). There is no evidence of self-incompatibility either, so it is technically possible to make self-pollinations and obtain viable botanical seed. Male sterility is a frequent phenomenon and is currently being used to measure pollen flow.
Drought stress on cassava: Cassava is sensitive to drought during the establishment stage, i.e., from planting to about 3 months after planting, was studied with special attention to root system development. Plants exposed to drought, regardless of whether early (from 9-44 Days After Planting (DAP)) or late (from 44-82 DAP), in the establishment period had significantly fewer leaves and lower shoot Dry Weight (DW) than well-watered controls. Late droughted plants manifested similar inhibition of shoot growth with those continuously droughted from 9-82 DAP. With regards to root development, early drought caused reduction in the number and length of adventitious roots, number of first, second and third order lateral roots and total root DW. The value of these parameters generally increased after the plants were rewatered. Late drought caused reduction in the number and length of the adventitious roots and the number of first and second order lateral roots but not the third order lateral roots which appeared to be promoted instead by drought. The total root DW also increased in late-droughted plants due to the thickening of adventitious roots. Overall, the results show that cassava has sensitivity to drought stress during its establishment period.
CURRENT BREEDING GOALS ON CASSAVA
Breeding objectives depend on the ultimate use of the crop. In cassava, however, increased yield, drought resistance, multiple pest and disease resistance, desirable agronomic traits such as appropriate plant architecture, early bulking of storage roots, with high dry-matter content, low cyanide content resistance drought and other a biotic stress and consumer preference traits, e.g., easy peeling and early vigour in plant growth (for high foliage yield for leaf vegetable) have been the main breeding objectives. Recently breeding for improved micronutrient content has been emphasized (Gregorio, 2002).
Broad-based populations from different agro-ecologies are developed through recurrent selection (intra and inter specific population improvement techniques) and backcrossing as well as multiple crossing schemes. Superior individuals are selected as parents and segregating families are generated by multiple crossings among these elite clones, complementing one another for various agronomic, consumer-quality and major pest resistance traits. The breeding values of the parents are evaluated through progeny testing in seedling nurseries. Based on evaluations, selected parental clones or half-sib progenies are hybridized for further improvement in a recurrent selection scheme. Backcrossing has also been a useful procedure for the transfer of resistance into elite populations by providing resistant lines quickly to prevent the severe infestation of relevant pests.
High and stable productivity is also a key trait for the future but there will also be increasing opportunities and needs for developing cassava cultivars particularly suited to specific end uses. It is therefore envisaged that multipurpose varieties will gradually give way to specific varieties with special quality traits developed for different processing pathways. These changes made clear that, stable productivity cassava breeding had the opportunity of expanding and exploiting genetic variability that would generate clones with increased value for the different industrial processes where cassava can be a strategic raw material. When cassava is used as a food security crop, additional requirements need to be addressed for a variety to be adopted.
Breeding of cassava for quality: Root quality characteristics that are often considered in breeding schemes include cyanogenic potential, starch quality, protein and dry matter contents. Attempts have been made to improve the protein content of cassava roots through conventional breeding methods involving hybridization at inter-specific levels as well as by induced polyploidy and mutation (Asiedu et al., 1992). There are different researches were conducted on the quality of cassava. Based on the genomic level, the protein content of the diploid (2n) and tetraploid (4n) cassava plants and found that the average crude protein in the tetraploid was 42.3% higher than in the diploid. In contrast with this, Kawano (2003) suggested that there is no significant variability in protein content screening with in large number of cassava genotype. In line with this, Chavez et al. (2005) reported that there was no correlation between dry matter and protein contents in the root but a weak positive correlation (p = 0.14) was observed between nitrogen and cyanogenic glycoside (HCN) contents in the roots of cassava. This might be due to the genetic potential of the genotypes in that specific environment. In cassava types (sweet and beater) the content cyanogenic glycoside is highly correlated with that of the amounts of starch content in cassava root (Mulualem and Kifle, 2013).
Achievements of conventional breeding on cassava: Conventional breeding has contributed to improvement in dry-matter and starch content as well as reduction in the cyanogenic glycoside content in cassava. Achievement in cassava breeding in Africa is in the development of a range of selected genotypes that combine high stable yields, agronomic and consumer quality with acceptable levels of resistance to Cassava Mosaic Disease (CMD) and Cassava Bacterial Blight (CBB). The introduction of Latin American germplasm into the breeding programs in Africa resulted in significant broadening of the genetic base of cassava in Africa. Genotypes with high efficiency for extraction of limited soil water, low levels of water loss through transpiration and resistance to severe mite attacks were selected and recombined by genetic crosses to form the basis of a breeding population for the semi-arid agro ecosystem (CIAT., 2009). Subsequently, a farmer participatory plant breeding scheme was employed to evaluate recombinant progenies and select several varieties adapted to the tropical environments of the world (Fukuda and Saad, 2001).
Currently improved cassava varieties produced 25-100% yield increases over the traditional cultivars in 41 communities of Brazil (CIAT., 2009). Recombinant seeds from the above breeding population for semi-arid ecosystems were also introduced to Sub-Saharan Africa (hereafter Africa) through IITA and evaluated in the drier areas of northern Nigeria. Evaluations revealed the potential of the germplasm to increase productivity in the dry and hot conditions of northern Nigeria. However, the presence in Africa of a major biotic constraint, CMD, a disease not found in Latin America, limited immediate use of the germplasm but required introgression of CMD resistance into the Latin American germplasm. Resistance to CMD was introduced by backcrosses and thousands of recombinant seeds were produced and distributed to participating countries in tropical Africa. Evaluation under typical farmer conditions and selection has led to the selection of 25, 32, 10, 28 and 122 improved genotypes that c ombine CMD resistance and adaptation to water stress for the semi-arid areas of different countries. The improved germplasm has extended considerably the range of cultivation of cassava beyond its traditional area in the humid and sub-humid tropics into the semi-arid zone of Africa in extensive area of land.
Conventional plant breeding has one of the highest rates of return among the investments in agricultural research. Cassava has also benefited from technological inputs in the area of breeding (Kawano, 2003). New varieties in Africa, Asia and Latin American countries have satisfied the needs of farmers, processors and consumers, bringing millions of dollars in additional income to small farmers.
Participatory plant breeding has a clear advantage for subsistence farming where many subtle criteria define the success of a given variety and its chances of adoption by farmers. The impact of conventional breeding in cassava tends to reach a maximum when and where the crop is largely used for processing. Yield productivity in tropical areas of Asia increased an average of about 0.5 t ha-1 year-1 since 1990, whereas in South America and Africa the increase was much slower with 0.09 t ha-1 year-1. In addition to increases in productivity, Kawano (2003) reported a major improvement in dry matter content of cassava varieties released in SE Asia and also demonstrated the importance of selection for adequate levels of harvest index, particularly in early stages of the selection process. Jennings and Iglesias (2002) provided an assessment of the significant progress achieved to develop cassava cultivars tolerant to the main viral diseases, bacterial blight and super-elongation diseases. Resistance to Cassava Mosaic Disease (CMD) has been deployed and analyzed from the molecular point of view (Egesi et al., 2007). Important progress in identifying and deploying tolerance/resistance to cassava diseases has also been achieved in recent years (Mcsween et al., 2006).
LIMITATION OF CONVENTIONAL BREEDING
Most of important traits in cassava are polygenic (Amma and Sheela, 1995). Variation in polygenic traits is attributed to Quantitative Trait Loci (QTLs). Quantitative traits in plants are studied using a variety of genetic models and designs including the analysis of mating designs in segregating populations estimate effective factors using biometrical techniques. Biometrical methods can also be used to estimate useful factors for quantitative traits (Lynch and Walsh, 1998). Most biometrical techniques estimate not the exact number of genes but the number of effective factors of quantitative traits. Furthermore, the detection of multiple genes for a trait using segregation analysis alone is not efficient because of the differences due to the genetic make up of genotype by environment interaction.
Breeding of cassava for moisture stress environment: The cassava plant is affected by a biotic stresses. These stresses can result in severe yield loss on cassava production. Cassava exhibits various responses to these stresses at the molecular, cellular and whole plant levels. These responses may contribute to increased tolerance to the stresses. Breeding on cassava to stress tolerance, is an imperative to identify the genes that control these traits and to understand how these genes and their products are regulated to resistance for stresses (Nassar and Ortiz, 2008).
Stable production relies on tolerance to a biotic stresses which may vary with the environment. There are a variety of abiotic factors limiting cassava productivity which would probably accentuate as a result of changes in the climate generating wider fluctuations in relevant weather parameters. Cassava is mainly grown in drought-prone areas and/or on low-fertility soils. Some traits associated with adaptation to these conditions have been suggested (Jennings and Iglesias, 2002), such as leaf structure (Lenis et al., 2006), optimum leaf area index and ideal plant architecture (Kawano, 2003). The capacity of the stems to withstand long storage periods (sometimes up to 2 months) from harvest to planting is an important trait. This characteristic affects final density of established plants and is fundamental for areas with relatively long dry spells or erratic rainfall, because the storage period may extend to the point that it compromises their viability. Basically, the main physiological mechanisms that cassava resists drought environments are shade there leaves when drought come, hinder its vegetative growth; its stomata is located under side of the leaves (it is important to reduce respiration), close its stomata before drought come and the upper part of the leaf covered by mucilaginous substances, it is important to protect water loss from the plant. Thus, these physiological mechanisms promote the plant to adapt drought full environment.
Although it is a self-inflicted reaction, post harvest physiological deterioration and the resulting short shelf life of cassava roots after harvest is frequently grouped as a biotic stress. Consequently, cassava roots need to be consumed or processed soon after harvesting (Van Oirschot et al., 2000). The short shelf life of the roots severely limits the marketing options by increasing the probability of losses, the overall marketing costs and by limiting the access to urban markets or processing canters to production sites close to them. Extending the shelf life by only 2-3 weeks would offer huge advantages to the cassava community.
Evaluation and selection scheme used in cassava for moisture stress: Cassava genetic improvement starts with the assembly and evaluation of a broad germplasm base, followed by production of new recombinant genotypes derived from selected elite clones and careful evaluation in a set of representative environments. Selection cassava breeding began only a few decades ago and therefore, the divergence between landraces and improved germplasm is not as wide as in crops with a more extensive breeding history. As a result, landrace accessions, as explained above, probably play a more relevant role in cassava than in other crops.
Parental lines are selected based mainly on their performance and little progress has been made in using general combining ability as a criterion of parental selection. The botanical seed obtained by the different crossing schemes (Kawano, 2003) may then be planted directly in the field or first germinated in greenhouse conditions and then transplanted to the field when they are about 20-25 cm tall. Root systems in plants derived from botanical seed or vegetative cuttings may differ considerably. The taproots from seedlings tend to store fewer starches than roots from cuttings (Alves, 2002). Because of this, it is difficult to correlate the root yield of clones at later stages in the evaluation/selection process, with early results from the plants obtained from botanical seeds (Morante et al., 2005). However, when seeds are germinated in containers and later transplanted, the taproot often does not develop and the seedling-derived plant may be more similar to subsequent stake-derived plants regarding the shape of their starchy roots.
For general purpose breeding, once a set of crosses is planned, it takes two years for the crosses to be ready. Certain crosses can be made within a year but crossing blocks are generally maintained for up to two years in the field thus allowing several flowering events per plant so numerous seeds per cross and many crosses per parental line can be obtained. Most of the seed is obtained from the second flowering event about 6-8 Months After Plant (MAP) through the third or fourth event (about 14-18 MAP). The last pollinations are then made early enough for the seed to mature and be harvested and processed for germination.
The vegetative multiplication rate of cassava is low. From one plant, 5-10 cuttings typically can be obtained, although this figure varies widely by genotypes. This situation implies a lengthy process to reach the point where replicated evaluations across several locations can be conducted, just because of the time required to produce enough planting material. It takes about 5-6 years from the time the botanical seed is germinated until the evaluation/selection. One further complication in a cassava breeding program is the number of factors that can affect quality of planting material. For example, the original positioning of the vegetative cutting along the stem affects considerably the performance of the plant it originates.
Cuttings from the mid-section of the stems usually produce better performance in diverse environment than those at the top or the bottom. This variation in the performance of the plant, depending on the physiological status of the vegetative cutting, results in larger experimental errors and undesirable variation in the evaluation process. There is some variation among different cassava breeding programs regarding the numbers of genotypes and plants representing them through the different stages of selection. However, the numbers presented in Table 2 are fairly common and illustrate the different stages required to complete a selection cycle and the kind of selection pressures that are generally applied.
The first selection can be conducted in the third year on the nurseries with plants derived from botanical seed (F1 in Table 2). Because of the low correlations between the performance at this early stage of selection and when the genotypes reach replicated trials, the early selections are based on high heritability traits, such as plant type, branching habits and particularly, reaction to diseases (Morante et al., 2005).
The capacity to produce this number of cuttings is in fact another selection criterion used at the F1 stage. Clonal evaluation trials usually range from 2,000-3,000 clones. Within a given trial, however, the same number of plants is used to avoid the confounding effects between number of plants and genotypic differences. Because the competition between neighboring genotypes in the clonal evaluation trials may favor more vigorous plant architectures, selection at this stage still relies heavily on high heritability traits, such as harvest index (Kawano, 2003).
Plant type is an important selection criterion at early stages of selection; plants whose main stem does not branch until it reaches at least one meter are preferred. Other selection criteria at this stage include high dry matter and cyanogenic potential. Between 100 and 300 clones survives the clonal evaluation trial. A common feature in the first two stages of selection for most programs is that selection is frequently visual with no data recording to manage a larger number of materials at lower costs. One important trait that makes the harvest of large trials, such as the clonal evaluation trial, expensive and time demanding is the measurement of dry matter content in the roots.
The productivity of cassava depends ultimately on the amount of fresh roots produced and the dry matter content of those roots. Heritability for dry matter content is considered to be intermediate. The following stage of selection is the preliminary yield trial. At (CIAT., 2009), preliminary yield trials are currently based on the evaluation of 10 plants in three replications. The ten plants in each replication are planted in two 5-plant rows. If possible, rows are spaced only 0.8 m apart (instead of the standard 1.0 m) and one empty row is left between plots to increase within-clone competition and reduce between-clone competition. Alternatively, row spacing can be maintained at 1.0 m but then the plant-to-plant distance within the row is reduced to 0.8 m. In this case also an empty row is left separating plots with different genotypes. Large genetic variability occurs among clones, even within the same family. Although poorly performing clones are mostly eliminated at the clonal evaluation trial stage, there is still a considerable variation in the preliminary yield trial trials.
| Table 2: | Typical recurrent selection scheme for an individual target environment (a consistent reduction of genotypes is also achieved through selection) |
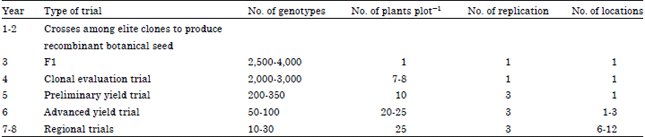 | |
| Source: Jennings and Iglesias (2002) and Ceballos et al. (2004). Cited by Lebot (2009) | |
This highlights the need for a gradual process of selection and the need to avoid strong selection pressures in early stages. With the initiation of replicated trials, the emphasis of selection shifts from high heritability traits to those of low heritability, such as root yield. Starting with preliminary yield trial and increasingly during the advanced yield trials and the regional trials, a greater weight is given to yield and its stability across locations. Cooking quality trials (relevant for the different ethnic ways cassava may be consumed) also begin at these stages, when the number of genotypes evaluated is more manageable.
The advanced yield trials are typically grown in 1-3 locations for two consecutive years. They have three replications per trial and plots are four (or five) rows with five plants per row. Yield data are taken from the six (nine) central plants of the plot and the remaining 14 (16) plants are used as source of planting material for the next season. The regional trials are conducted for at least 2 years in 3-6 locations each year. Plots have five rows with five plants per row. Yield data are taken from the nine plants from the centre. Clones that show an outstanding performance in the regional trials are released as new varieties after a few years of informal evaluation in semi-commercial evaluations with key farmers. They are also sooner rather than later incorporated into the crossing blocks as progenitors to initiate a new recurrent selection cycle.
The breeding scheme described above can be classified as mass phenotypic recurrent selection. No family data are used in the selection process and individual clones are evaluated and selected or discarded. It has been suggested that data are recorded in all entries at the clonal evaluation trial level (Ceballos et al., 2007) and estimates of general combining ability of the progenitors that generated the clonal evaluation trial are obtained.
Physiological response of cassava for moisture stress tolerance: Several physiological traits such as Leaf Water Potential (LWP), Osmotic Adjustment (OA), heat tolerance, desiccation tolerance, rooting depth and epicular wax (Ludlow, 1993) are known to contribute to moisture stress tolerance. Screening techniques and genetic variability based on some of these traits (LWP and OA) have been reported (Blum, 1987).
Leaf Water Potential (LWP): The physiological adaptations effective in improving tolerance to moisture stress were found to vary with plant growth stage in cassava (Ackerson et al., 1980). Before flowering, plants avoid dehydration largely by maintaining higher LWP, after flowering, plants avoid dehydration by maintaining higher turgor at a given level of moisture stress. This activity could be partly responsible for the different classification of moisture stress tolerance before and after flowering in cassava (Rosenow and Wendt, 1983). The most evident control of LWP is at the root system. Small root resistances and a large root-length density would contribute to the maintenance of a higher LWP. The root-length density increased with reduced soil moisture only at certain soil depths (Blum and Arkin, 1984). Genotypic differences in cassava root growth at early stages have been noted under moisture stress. Early-maturing cassava genotypes not only escape moisture stress but also avoid it because of reduced transpiration as a result of increased root length accompanied by reduced leaf area (high root length to leaf area ratio). Because of high sensitivity of leaf area expansion to changes in turgor, several research reports (Blum, 1987) suggested that the use of leaf area expansion as the criterion for screening the cassava genotypes for moisture stress tolerance. Large differences for leaf expansion rates among cassava cultivars and hybrids have been observed (Nassar, 2004). Some hybrid genotypes are sensitive to moisture stress at early stage, showed more reduction in leaf expansion than moisture stress tolerant. Leaf dropping and firing are also found to be a simple phenotypic trait that allows large populations to be screened (Ludlow, 1993).
Leaf rolling is an established symptom of wilting in cereals not common in cassava (Jones and Turner, 1978; Beyene, 2012) and delayed leaf rolling under moisture stress is being used as one component of a selection index for moisture stress tolerance (avoidance) in cassava (Rosenow and Wendt, 1983). Greater leaf rolling is indicative of reduced LWP in different cassava genotypes (Blum, 1987). Gaosegelwe and Kirkham (1990) suggested that LWP could be used as an easy and fast way to screen cassava genotypes for moisture stress avoidance. Under relatively mild stress, delayed leaf rolling may be associated with sustained plant growth and production. However, under severe moisture stress and heat stress conditions, greater leaf rolling may be associated with better chances for recovery when moisture stress is relieved (Blum, 1987). Stricevic and Caki (1997) showed a predawn LWP of 0.4 MPa as the threshold value for scheduling irrigation. Physiological processes are significantly decreased below this value which suggests that those genotypes that maintain predawn LWP above this level can be considered as moisture stress tolerant. However, breeding programs are slow to adopt this trait for selection.
Osmotic Adjustment (OA): Prolonged periods of water stress is a characteristic of low tissue water potential and tolerant plants adapt to low tissue water potential (OA). The genotypes with high OA retain higher turgor at a given level of plant water deficit and produce higher leaf area and subsequently support carbon assimilation. Based on this, Tangpremsri et al. (1991) concluded that the adverse effect of water stress could be reduced by selecting cassava genotypes with high OA. However, Flower et al. (1990) concluded that while moisture stress tolerant cassava cultivars had better OA and consequently less leaf rolling under stress compared with susceptible cultivars, these responses did not influence growth under very dry and hot conditions. Studies on OA have been accelerated by the use of pressure chamber method and analysis of pressure volume graphs to measure water, osmotic and turgor potentials. Thermocouple psychrometry has also aided in measurement of water and osmotic potentials (Parsons, 1982).
Variation in Osmotic Adjustment (OA) among cassava genotypes was found to range from nil to 1.6 Mpa (Rosenow et al., 1997). Landraces from dry habitats compared to those from humid regions have greater capacity for OA. Diurnal and seasonal OA to water stress have been noted in cassava (Jones and Turner, 1978). OA has direct positive effect on yield under moisture stress (Ludlow, 1993) and is largely ascribed to increase in root size, root length density and soil moisture extraction (Tangpremsri et al., 1991). Genotypic differences have been detected in predawn osmotic potentials even under mild stress.
There is a little progress in breeding for cassava tolerance using OA and other physiological traits has been documented in cassava, partly because of poor understanding of the traits conferring moisture stress tolerance and lack of procedures to impose reproducible stresses and rapid methods to measure these traits (Tangpremsri et al., 1991). It appears that individual physiological traits identified to date are not sufficiently related to overall moisture stress tolerance under field conditions to merit selection based on them (Rosenow et al., 1997).
Root architecture: The two major architectural attributes of roots concerning water uptake are root length density (total length of roots per unit soil volume) and root depth. Both root depth and root length density are determined firstly by the basic (potential) root architecture and its development in time as the plant develops and as the soil dries.
Root architecture varies extensively among crop species and genotypes. It also varies extensively with soil physical, chemical and biological characteristics. There is a basic genetic control of root growth and development. Various mutants for root growth were identified and even implicated in moisture stress resistance but the developmental interactions in the plant and plant response to the soil have a large effect on transient or final root architecture and function under moisture stress. Root plasticity is still a problem for developing a realistic root module in crop simulation models even if fractal theories are considered (Lynch, 1995). It is also a problem for the breeder in terms of setting a realistic target for selection. The minimum that one can project are some basic macroscopic potential root traits and functions that should be considered in the breeding program. It should be remembered that the root is a well organized system no less than the shoot.
There is a basic difference between crops with a tap root system such as cotton and the root system of the cereals. A single axial root system with branching is far more predictable than the cereal root system. Initially the cereal root system is composed of the seminal roots and the adventitious (crown) roots. Adventitious roots develop on a temporal cycle from axillary buds in the lower stem internodes, as in maize or sorghum. In highly tillering plants such as small grains or rice, adventitious rooting is associated with tiller production and establishment.
Profusely tillering cultivars tend to have a profuse and relatively shallow root system while limited-tillering cultivars tend to have a less profuse and relatively deeper root system, as a general design. Porter et al. (1986) described well these associations between seminal roots, adventitious roots and tiller development in wheat. When moisture stress develops some of the smaller existing tillers tend to abort and new tiller development is arrested. This is an effective mechanism for the plant to limit water use and develop deeper roots. The effect of limited tillering on root architecture and depth should not be confused with root biomass. Restricted tillering in wheat did not result in relatively greater root biomass when plants were grown in root boxes and irrigated daily (Palta et al., 2007). There is ample information in the literature indicating large genotypic differences in root depth or maximum root length among genotypes of cultivated crop species. The variation is found among cultivars, germplasm and exotic genetic resources such as landraces. Especially, molecular mapping studies of root architecture, morphology and growth with the recognition that deep penetrating roots are very important in this crop under moisture stress condition.
CONCLUSION
Cassava is a crop of major economic important and food security crop in tropical world. The crop cultivated as the main source of energy and as the most important staple food crop for over 900 million of people in many regions of Africa, Asia and South America. It is the second most important staple crop in Africa after maize. Despite in its importance, production and productivity of cassava is constrained by different biotic and abiotic factors. Consequently, conventional breeding of stress on cassava have a paramount importance for selection, development of resistant and adaptive genotypes for variable environments.
Breeding in any crop can be implemented based on its objectives and the ultimate use of the crop, for cassava, however, high yield, combining selection for moisture stress resistance and high yield, multiple pest and disease resistance, desirable agronomic traits such as appropriate plant architecture, early bulking of storage roots, with high dry-matter content, low cyanide content and consumer preference traits are the most important factors for selection and crossing on cassava. Thus, breeding associated with these traits/factors are important for the production as well as the livelihood of farmers who live in tropical and subtropical environments.
The choice of breeding method for cassava is largely dictated by its reproductive system and plant breeding goals but, generalized procedures of vegetative and crossing plants as well as established with provisions of suitable changes for the crop. Thus, identification and selection of efficient breeding methods are the key ingredients of progress in cassava breeding on stressful environment.
Physiologically, cassava has own mechanism to adapt moisture stress, by shade there leaves when moisture stress comes; hinder its vegetative growth; naturally, its stomata is located under side of the leaves and important to reduce respiration; close its stomata before moisture stress appear and the upper part of the leaf covered by mucilaginous substances are a few to mentioned. Finally, breeding based on these characters have tremendous impact on the development of genotypes for moisture stress resistance.
REFERENCES
- Ackerson, R.C., D.R. Krieg and S.J.M. Sung, 1980. Leaf conductance and osmoregulation of field-grown sorghum genotypes. Crop Sci., 20: 10-14.
CrossRef - Asiedu, R., S.K. Hahn, K.V. Bai and A.G.O. Dixon, 1992. Introgression of Genes from Wild Relatives into Cassava. In: Promotion of Root Crop-based Industries: An Incentive for Research and Development: Proceedings of the Fourth Triennial Symposium of the International Society for Tropical Root Crops-Africa Branch held in Kinshasa, Zaire, 5-8 December 1989, Akoroda, M.O. and O.B. Arene (Eds.). ISTRC-AB, IITA, Ibadan, Nigeria, ISBN-13: 9789781310850, pp: 89-91.
- Bartels, D. and R. Sunkar, 2005. Drought and salt tolerance in plants. Crit. Rev. Plant Sci., 24: 23-58.
CrossRefDirect Link - Benesi, I. R. M., M.T. Labuschagne, A.G.O. Dixon and N.M. Mahungu, 2004. Genotype X enviroment interaction effects on native cassava starch quality and potential for starch use in the commercial sector. Afr. J. Crop Sci., 12: 205-216.
CrossRefDirect Link - Blum, A. and G.F. Arkin, 1984. Sorghum root growth and water-use as affected by water supply and growth duration. Field Crops Res., 9: 131-142.
CrossRefDirect Link - Ceballos, H., C.A. Iglesias, J.C. Perez and A.G.O. Dixon, 2004. Cassava breeding: Opportunities and challenges. Plant Mol. Biol., 56: 503-516.
CrossRefDirect Link - Ceballos, H., T. Sanchez, N. Morante, M. Fregene and D. Dufour et al., 2007. Discovery of an amylose-free starch mutant in cassava (Manihot esculenta Crantz). J. Agric. Food Chem., 55: 7469-7476.
CrossRefDirect Link - Egesi, C.N., F.O. Ogbe, M. Akoroda, P. Ilona and A. Dixon, 2007. Resistance profile of improved cassava germplasm to cassava mosaic disease in Nigeria. Euphytica, 155: 215-224.
CrossRefDirect Link - Elias, M., D. McKey, O. Panaud, M.C. Anstett and T. Robert, 2001. Traditional management of cassava morphological and genetic diversity by the Makushi Amerindians (Guyana, South America): Perspectives for on-farm conservation of crop genetic resources. Euphytica, 120: 143-157.
CrossRefDirect Link - Flower, D.J., A.U. Rani and J.M. Peacock, 1990. Influence of osmotic adjustment on the growth, stomatal conductance and light interception of contrasting sorghum lines in a harsh environment. Aust. J. Plant Physiol., 17: 91-105.
CrossRefDirect Link - Gaosegelwe, P.L. and M.B. Kirkham, 1990. Evaluation of Wild, Primitive and Adapted Sorghums for Drought Resistance. In: Challenges in Dryland Agriculture: A Global Perspective: Proceedings of the International Conference on Dryland Farming, August 15-19, 1988, Amarillo/Bushland, Texas, USA., Unger, P.W., T.V. Sneed, W.R. Jordan and R.W. Jensen (Eds.). Texas Agricultural Experiment Station, College Station, TX., USA., pp: 224-226.
Direct Link - Gregorio, G.B., 2002. Progress in breeding for trace minerals in staple crops. J. Nutr., 132: 500S-502S.
Direct Link - Hahn, S., K. Bai and R. Asiedu, 1990. Tetraploids, triploids and 2n pollen from diploid interspecific crosses with cassava. Theor. Applied Genet., 79: 433-439.
CrossRefPubMedDirect Link - Ito, K., K. Tanakamaru, S. Morita, J. Abe and S. Inanaga, 2006. Lateral root development, including responses to soil drying, of maize (Zea mays) and wheat (Triticum aestivum) seminal roots. Physiol. Plant, 127: 260-267.
CrossRefDirect Link - Jones, M.M. and N.C. Turner, 1978. Osmotic adjustment in leaves of sorghum in response to water deficits. Plant Physiol., 61: 122-126.
Direct Link - Kapinga, R., A. Westby, E. Rwiza, Z. Bainbridge and A. Nsanzugwanko, 1998. Diversification of Cassava utilisation in the Lake Zone of Tanzania: A case study. Trop. Agric., 75: 125-128.
Direct Link - Kawano, K., 2003. Thirty years of cassava breeding for productivity-biological and social factors for success. Crop Sci., 43: 1325-1335.
CrossRefDirect Link - Lenis, J.I., F. Calle, G. Jaramillo, J.C. Perez, H. Ceballos and J.H. Cock, 2006. Leaf retention and cassava productivity. Field Crop Res., 95: 126-134.
CrossRefDirect Link - Mcsween, S., T. Walker, V. Salegua and R. Pitoro, 2006. Economic impact on food security of varietal tolerance to cassava brown streak disease in coastal Mozambique. Research Report Series No. 1E, Directorate of Training, Documentation and Technology Transfer, Institute of Agricultural Research of Mozambique, Maputo, Mozambique, August 2006, pp: 1-33.
- Mitra, J., 2001. Genetics and genetic improvement of drought resistance in crop plants. Curr. Sci., 80: 758-763.
Direct Link - Nassar, N.M.A., 2002. Cassava, Manihot esculenta Crantz, genetic resources: Origin of the crop, its evolution and relationships with wild relatives. Genet. Mol. Res., 1: 298-305.
Direct Link - Nguyen, H.T., R.C. Babu and A. Blum, 1997. Breeding for drought resistance in rice: physiology and molecular genetic considerations. Crop Sci., 37: 1426-1434.
CrossRefDirect Link - Palta, J.A., I.R.P. Fillery and G.J. Rebetzke, 2007. Restricted-tillering wheat does not lead to greater investment in roots and early nitrogen uptake. Field Crops Res., 104: 52-59.
CrossRefDirect Link - Porter, J.R., B. Klepper and R.K. Belford, 1986. A model (WHTROOT) which synchronizes root growth and development with shoot development for winter wheat. Plant Soil, 92: 133-145.
CrossRefDirect Link - Pujol, B., G. Gigot, G. Laurent, M. Pinheiro-Kluppel, M. Elias, M. Hossaert-McKey and D. McKey, 2002. Germination ecology of cassava (Manihot esculenta Crantz, Euphorbiaceae) in traditional agroecosystems: Seed and seedling biology of a vegetatively propagated domesticated plant. Econ. Bot., 56: 366-379.
CrossRefDirect Link - Rosenow, D.T., G. Ejeta, L.E. Clark, M.L. Gilbert, R.G. Henzell, A.K. Borell and R.C. Muchow, 1997. Breeding for pre- and post-flowering drought stress resistance in Sorghum. Proceedings of the International Conference on Genetic Improvement of Sorghum and Pearl Millet, September 22-27, 1996, Lubbock, TX., USA., pp: 400-411.
Direct Link - Sambatti, J.B.M., P.S. Martins and A. Ando, 2001. Folk taxonomy and evolutionary dynamics of cassava: A case study in Ubatuba, Brazil. Econ. Bot., 55: 93-105.
CrossRefDirect Link - Sorrels, M.E., A. Diab and M. Nachit, 2000. Comparative Genetics of Drought Tolerance. In: Durum Wheat Improvement in the Mediterranean Region: New Challenges, Royo, C., M. Nachit, N. Di Fonzo and J.L. Araus (Eds.). CIHEAM, Zaragoza, Spain, pp: 191-201.
Direct Link - Stricevic, R. and E. Caki, 1997. Relationships between available soil water and indicators of plant water status of sweet Sorghum to be applied in irrigation scheduling. Irrig. Sci., 8: 17-21.
CrossRefDirect Link - Subbarao, G.V., O. Ito, R. Serraj, J.J. Crouch and S. Tobita et al., 2005. Physiological Perspectives on Improving Crop Adaptation to Drought-Justification for a Systematic Component-based Approach. In: Handbook of Photosynthesis, Pessarakli, M. (Ed.). 2nd Edn., Marcel and Dekker, New York, USA., pp: 577-594.
- Tangpremsri, T., S. Fukai, K.S. Fischer and R.G. Henzell, 1991. Genotypic variation in osmotic adjustment in grain sorghum. II. Relation with some growth attributes. Aust. J. Agric. Res., 42: 759-767.
CrossRefDirect Link - Mulualem, T. and A. Kifle, 2013. Studies on correlation and path analysis for root yield and related traits of cassava (Manihot esculenta Crantz) in South Ethiopia. Unique Res. J. Agric. Sci., 1: 34-39.
Direct Link - Van Oirschot, Q.E.A., G.M. O'Brien, D. Dufour, M.A. El-Sharkawy and E. Mesa, 2000. The effect of pre-harvest pruning of cassava upon root deterioration and quality characteristics. J. Sci. Food Agric., 80: 1866-1873.
CrossRef - Venuprasad, R., C.O. Dalid, M. del Valle, D. Zhao and M. Espiritu, 2009. Identification and characterization of large-effect quantitative trait loci for grain yield under lowland drought stress in rice using bulk-segregant analysis. Theor. Applied Genet., 120: 177-190.
CrossRefDirect Link - Chavez, A.L., T. Sanchez, G. Jaramillo, J.M. Bedoya and J. Echeverry et al., 2005. Variation of quality traits in cassava roots evaluated in landraces and improved clones. Euphytica, 143: 125-133.
CrossRefDirect Link