
Shota Katayama
Department of Agriculture and Life Science, Obihiro University of Agriculture and Veterinary Medicine, 2-11 Inada-Cho, Obihiro, Hokkaido, 080-8555, Japan
Asian Journal of Agricultural Research
Year: 2015 | Volume: 9 | Issue: 6 | Page No.: 357-361







ABSTRACT
A novel protein, bovine colostral Odorant-Binding Protein (bcOBP) was found in bovine colostrums. This protein belongs to the lipocalin superfamily, most of them are secretary proteins. Amino acid sequences of bcOBP have CxxxC motif, which is well conserved among mammalian pheromone binding proteins. Its biological function is unclear, but pheromone transport is considered. Vomeronasal organ, whose important role is supposed to be in pheromone detection was suspected to express bcOBP mRNAs. The expression was found in vomeronasal organ, nasal mucosa and olfactory epithelium. Surprisingly, the expression was also found in lacrimal grand. The presence of bcOBP mRNAs in the area around the olfactory sense and in lacrimal gland might imply a functional role on pheromone transport and on sexual behavior between male and female like Exocrine gland-secreting peptide 1 (ESP1), respectively.
PDF Abstract XML References Citation
Received: July 28, 2015;
Accepted: September 14, 2015;
Published: September 21, 2015
How to cite this article
Shota Katayama, 2015. Putative Roles of Bovine Colostral Odorant-binding Protein (bcOBP) for Pheromone Transport and Sexual Behavior. Asian Journal of Agricultural Research, 9: 357-361.
DOI: 10.3923/ajar.2015.357.361
URL: https://scialert.net/abstract/?doi=ajar.2015.357.361
DOI: 10.3923/ajar.2015.357.361
URL: https://scialert.net/abstract/?doi=ajar.2015.357.361
INTRODUCTION
Olfaction involves the binding of small, hydrophobic, volatile molecules to receptors of the nasal neuroepithelia (Buck and Axel, 1991). It generates a cascade of neurological events that transmit the information to the olfactory bulbs projecting into the brain. The first step in this process is the solubilization of these hydrophobic molecules in the hydrophilic nasal mucus. Odorant-Binding Proteins (OBPs) are thought to transport these molecules within the mucus (Pelosi, 1996). The OBPs were first identified in the nasal mucosa (Pelosi et al., 1982; Bignetti et al., 1985; Pevsner et al., 1985) in which human was able to find OBPs bind volatile compounds and expressed.
These proteins belong to the lipocalin superfamily of which biochemical structures are well suited for this function. This superfamily comprises >100 small proteins secreted in various biological fluids (Pervaiz and Brew, 1987). They contain eight consecutive β-sheets forming a barrel-shaped fold in which hydrophobic pocket is present (Flower, 1996).
Lipocalins in the mucus of the oral and nasal sphere epithelia act as biosensor proteins for the detection of environmental signals, especially pheromones. Odorants, which are chemically diverse, are distinguished at the neuroepithelium level using combinations of hundreds of receptors (Malnic et al., 1999). Some lipocalins specifically bind volatile pheromones, which can be regarded as a sub-class of odorants. These proteins are the mouse and rat urinary proteins, the hamster aphrodisin and salivary lipocalins in mouse and pig (Bocskei et al., 1992; Singer and Macrides, 1993; Marchese et al., 1998).
In 2009, a novel protein in bovine colostrum was found by Japanese research groups, which named bovine colostral Odorant-Binding Protein (bcOBP) (Fukuda et al., 2009). This protein, which consists of 172 amino acid residues including a putative 16 AA signal peptide, belongs to lipocalin superfamily considered as secretary proteins (Fukuda et al., 2009). The OBP derived from nasal mucosa showed low sequences similarity (52%) to bcOBP (Fukuda et al., 2009). Amino acid sequences of bcOBP have CxxxC motif, which is well conserved among mammalian pheromone binding proteins (Vincent et al., 2001; Srikantan et al., 2005; Stopkova et al., 2009). They proposed hypothesis that the function of bcOBP might be pheromone transport (Fukuda et al., 2009) and hence expected that vomeronasal organ (VNO) express bcOBP mRNAs.
MATERIALS AND METHODS
RNA isolation and reverse transcription: Total RNA was purified with Trizol reagent (Invitrogen) and treated with RQ1 RNase free DNase (Promega) to remove genomic DNA contamination. One microgram of total RNA was used for reverse transcription reaction with ReverTraAce (Toyobo) and random primer (Invitrogen), according to the manufacturer’s instructions. The PCR was performed with Ex Taq (Takara). Quantitative Polymerase Chain Reaction (PCR) was performed with Taqman RT-PCR using Premix Ex Taq (Takara) and analyzed with the 7300 real-time PCR system (Applied Biosystems).
Probe sequences used in quantitative PCR: 6-carboxyfluorescin (FAM), 6-carboxytetramethylrhodamine β-actin (TAMRA), Forward, 5’-TGCCATGTATGTGGCCATCC-3’; reverse, 5’-CGCTCGGCTGTGGTGGTAA-3’ probe, 5’-FAM-ACTCTGGGGACGGGGTCA-TAMRA-3’ bcOBP, Forward, 5’-GCACATGCCAGTTTTTCTCA-3’ ; reverse, 5’-GGAATCCCCCTCTCATTGTT-3’; probe, 5’-FAM-AAGGTTCCGCCAAAGGAACCA-TAMRA-3’.
Primers for RT-PCR: Glycerol-3-Phosphate Dehydrogenase (G3PDH) Forward, 5’-CGACCACTTTGTCAAGCTCA-3’; reverse, 5’-TCAGGGCCTTAGAGATGGAA-3’ bcOBP Forward, 5’-GCACATGCCAGTTTTTCTCA-3’; reverse, 5’-TTGGCGGAACCTTCAGTTAC-3’.
Tissue collection: We used female cows. All experiments were conducted in accordance with the Guiding Principles for the Care and Use of Research Animals of the Obihiro University of Agriculture and Veterinary Medicine, Japan.
RESULTS
bcOBP mRNAs are expressed in the area around the olfactory sense: There was a hypothesis that a functional role of bcOBP was considered to pheromone transport (Fukuda et al., 2009). Vomeronasal organ, whose biological function is supposed to be in pheromone detection was suspected to express bcOBP mRNAs. An mRNA of bcOBP was known to be expressed in mammary glands (UniGene: Bt.90525). To investigate bcOBP mRNA expression patterns around the olfactory sense, we isolated mRNAs from bovine tissue samples. RT-PCR analysis revealed that the expression was found in vomeronasal organ (VNO), nasal mucosa and olfactory epithelium (Fig. 1). Our results supported the hypothesis that bcOBP might be involved in pheromone transport. Surprisingly, the expression was also found in lacrimal grand (Fig. 1). This result proposes the hypothesis that a functional role on sexual behavior between male and female like ESP 1.
 | |
| Fig. 1: | RT-PCR analysis of bcOBP gene expression in several tissues. Primers for bcOBP and G3PDH are specific for transcripts from the respective endogenous locus. G3PDH was used as a positive control |
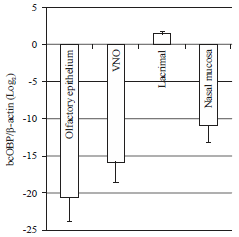 | |
| Fig. 2: | Quantitative PCR analysis of bcOBP transcription in several tissues. Transcript levels were normalized to β-actin. Data is shown in the averages and the standard deviations (n = 2) |
Quantification of bcOBP mRNA expression levels: To identify the amount of mRNA expression of bcOBP, TaqMan quantitative PCR (qPCR) was performed for the tissues in the area around the olfactory sense and for lacrimal gland. The qPCR analysis showed that the highest expression level in lacrimal gland has been observed (Fig. 2). Among olfactory sense tissues, the expressions are altered about ±5 or ±10 (Log2 conversion) (Fig. 2). In EST profile, the expression level of bcOBP mRNAs between olfactory sense tissues and lacrimal gland showed no expression at all (UniGene: Bt.90525). Our results demonstrated that bcOBP mRNA expression levels between olfactory sense tissues and lacrimal gland were not zero, but detectable. These results support the two hypothesis, (1) bcOBP might be related to sexual behavior and (2) bcOBP might be related to pheromone transport.
DISCUSSION
Prior to this study, a novel protein ESP1 was found in male mouse tear fluids and its biological function is to enhance female sexual receptive behavior through a specific vomeronasal receptor (Haga et al., 2010). This study provided us a new hypothesis that bcOBP might be related to sexual behavior between male and female. This study synthesized a biological model, which is consist of three steps, (1) bcOBP is expressed in female lacrimal gland, (2) Male cows recognize it in female through vomeronasal receptors and (3) Male sexual behavior will be enhanced. Our results demonstrated that bcOBP mRNA is abundantly expressed in lacrimal gland (Fig. 1 and 2), indicating that step (1) is correct. However, it was impossible to confirm step (2) and (3), because we could not obtain male cows and samples and not conduct experiments using them. Our model is still inconclusive and hence further study is required for proof of step (2) and (3).
In mammalian pheromone binding proteins (Vincent et al., 2001; Srikantan et al., 2005; Stopkova et al., 2009), CxxxC motif, which was first discovered in insect pheromone binding proteins (Du and Prestwich, 1995), are well conserved as well as bcOBP. Those mammalian pheromone binding proteins are intended to be distributed in olfactory sense (Guiraudie-Capraz et al., 2005). The distribution pattern is corresponded between bcOBP (Fig. 1 and 2) and other pheromone binding proteins, thus, bcOBP might be involved in pheromone transport.
CONCLUSION
This study provides a basis for further research into the ligand specificities and other crucial knowledge.
ACKNOWLEDGMENTS
We are grateful to Dr. Yoshiyasu Kobayashi, Dr. Hisashi Inokuma and their colleagues for technical supports for tissue samplings. This study is financially supported by the Global Center of Excellence "Frontier Program for Animal Global Health and Hygiene", Ministry of Education, Culture, Sports, Science and Technology, Japan and Japan Society for the Promotion of Science.
REFERENCES
- Bignetti, E., A. Cavaggioni, P. Pelosi, K.C. Persaud, R.T. Sorbi and R. Tirindelli, 1985. Purification and characterisation of an odorant-binding protein from cow nasal tissue. Eur. J. Biochem., 149: 227-231.
CrossRefDirect Link - Bocskei, Z., C.R. Groom, D.R. Flower, C.E. Wright and S.E.V. Phillips et al., 1992. Pheromone binding to two rodent urinary proteins revealed by X-ray crystallography. Nature, 360: 186-188.
CrossRefDirect Link - Buck, L. and R. Axel, 1991. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell, 65: 175-187.
CrossRefDirect Link - Du, G. and G.D. Prestwich, 1995. Protein structure encodes the ligand binding specificity in pheromone binding proteins. Biochemistry, 34: 8726-8732.
CrossRefDirect Link - Flower, D.R., 1996. The lipocalin protein family: Structure and function. Biochem. J., 318: 1-14.
CrossRefDirect Link - Fukuda, K., A. Senda, T. Ishii, T. Urashima, M. Morita and T. Terabayashi, 2009. Short communication: Evidence for the presence of a putative odorant-binding protein in bovine colostrum. J. Dairy Sci., 92: 4992-4996.
CrossRefDirect Link - Guiraudie-Capraz, G., M.C. Slomianny, P. Pageat, C. Malosse, A.H. Cain, P. Orgeur and P.N.L. Meillour, 2005. Biochemical and chemical supports for a transnatal olfactory continuity through sow maternal fluids. Chem. Senses, 30: 241-251.
CrossRefDirect Link - Haga, S., T. Hattori, T. Sato, K. Sato and S. Matsuda et al., 2010. The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature, 466: 118-122.
CrossRefDirect Link - Malnic, B., J. Hirono, T. Sato and L.B. Buck, 1999. Combinatorial receptor codes for odors. Cell, 96: 713-723.
CrossRefDirect Link - Marchese, S., D. Pes, A. Scaloni, V. Carbone and P. Pelosi, 1998. Lipocalins of boar salivary glands binding odours and pheromones. Eur. J. Biochem., 252: 563-568.
CrossRefDirect Link - Pelosi, P., N.E. Baldaccini and A.M. Pisanelli, 1982. Identification of a specific olfactory receptor for 2-isobutyl-3-methoxypyrazine. Biochem. J., 201: 245-248.
Direct Link - Pervaiz, S. and K. Brew, 1987. Homology and structure-function correlations between alpha 1-acid glycoprotein and serum retinol-binding protein and its relatives. FASEB J., 1: 209-214.
Direct Link - Pevsner, J., R.R. Trifiletti, S.M. Strittmatter and S.H. Snyder, 1985. Isolation and characterization of an olfactory receptor protein for odorant pyrazines. Proc. Natl. Acad. Sci. USA., 82: 3050-3054.
PubMedDirect Link - Srikantan, S., V. Parekh and P.K. De, 2005. cDNA cloning and regulation of two sex-hormone-repressed hamster tear lipocalins having homology with odorant/pheromone-binding proteins. Biochim. Biophys. Acta (BBA)-Gene Struct. Exp., 1729: 154-165.
CrossRefDirect Link - Stopkova, R., D. Hladovcova, J. Kokavec, D. Vyoral and P. Stopka, 2009. Multiple roles of secretory lipocalins (MUP, OBP) in mice. Folia Zool., 58: 29-40.
Direct Link - Vincent, F., D. Lobel, K. Brown, S. Spinelli and P. Grote et al., 2001. Crystal structure of aphrodisin, a sex pheromone from female hamster. J. Mol. Biol., 305: 459-469.
CrossRefDirect Link