
E. Abhilash Joseph
Department of Botany, Interuniversity Centre for Plant Biotechnology, University of Calicut, Kerala, 673635, India
V.V. Radhakrishnan
Department of Botany, Genetics and Plant Breeding Division, University of Calicut, Kerala, 673635, India
K.V. Mohanan
Department of Botany, Interuniversity Centre for Plant Biotechnology, University of Calicut, Kerala, 673635, India
Department of Botany, Genetics and Plant Breeding Division, University of Calicut, Kerala, 673635, India
Asian Journal of Agricultural Research
Year: 2015 | Volume: 9 | Issue: 6 | Page No.: 315-324







ABSTRACT
Rice plants respond to salinity stress in different ways and accumulation of phenolic compounds is one of them. An experiment was carried out presently to analyze the variation of accumulation of phenolics in some native rice cultivars of North Kerala, India in response to salt stress. Five cultivars namely Orthadian, Orkazhama, Kuthiru, Kuttusan and Chovvarian collected from one of the saline rice tracts of Kerala and two cultivars namely Kunhutty and Veliyan collected from one of the non-saline rice tracts of Kerala were used for the study. The plants were treated with gradually increasing concentrations of NaCl varying from 0-200 mM starting from the 45th day of growth onwards mimicking the gradually increasing salt content of the salinity prone rice farms of the study area and the quantity of total phenolics was investigated in each case. The cultivars collected from both the saline and non-saline rice tracts showed gradual increase in the accumulation of total phenolics in response to increase in salt stress. The cultivars showed differential variation in the accumulation of total phenolics in relation to salt stress showing that the character was cultivar specific. Among the cultivars studied, Kuthiru showed the highest quantity of phenolic compounds and Orkazhama showed the highest percentage of increase of phenolics over the control plants in relation to increase in salt stress. Salt stress tolerant cultivars can be further screened for the production of higher quantity of phenolics in response to salt stress with an objective of improving the antioxidant production potential of such cultivars.
PDF Abstract XML References Citation
Received: July 15, 2015;
Accepted: August 19, 2015;
Published: September 21, 2015
How to cite this article
E. Abhilash Joseph, V.V. Radhakrishnan and K.V. Mohanan, 2015. Variation in Accumulation of Phenolics in Some Native Rice Cultivars of North Kerala, India in Response to Salt Stress. Asian Journal of Agricultural Research, 9: 315-324.
DOI: 10.3923/ajar.2015.315.324
URL: https://scialert.net/abstract/?doi=ajar.2015.315.324
DOI: 10.3923/ajar.2015.315.324
URL: https://scialert.net/abstract/?doi=ajar.2015.315.324
INTRODUCTION
Rice (Oryza sativa L.) is the most important staple cereal food of nearly 50-80% of the world’s population either directly as human food or indirectly as animal feed (Osawa et al., 1992; Zhai et al., 2001; Zhou et al., 2002; Conforti and Sarris, 2011). Asia represents about 90% of global rice production and consumption (Osawa et al., 1992; Salekdeh et al., 2002). It is a semi-aquatic tropical C3 crop and hence it requires high level of soil moisture (McKersie and Lesham, 1994). Nutritional quality of rice has received more attention in the developing countries, where monotonous consumption of rice may lead to deficiencies of essential minerals, vitamins and other nutritional components (Asamarai et al., 1996; Bouis et al., 2003). Salinity is a prevalent abiotic stress that limits the productivity and geographical distribution of plants worldwide and induces a wide range of adverse metabolic responses in higher plants. High concentrations of salts disrupt homeostasis in water potential and ion distribution in plants (Dash and Panda, 2001). Soil salinity is becoming a serious problem in many coastal, arid and irrigated rice production systems. Increasing salinity of agricultural irrigation water together with progressive salinization of agricultural land is of increasing importance to agriculture because it limits the distribution of plants in certain natural habitats. The impact of salinity is the most serious in countries where all or most of the agricultural production is based on irrigation and when agriculture is a substantial part of the national economy (Mc Williams, 1986; Ghassemi et al., 1995).
Crucial changes in ion and water homeostasis lead to molecular damage, growth arrest and even death of the plant. Specific effects of salt stress on plant metabolism, especially on leaf senescence, have been related to the accumulation of toxic ions (Na+ and Cl-) or to K+ and Ca2+ depletion (Chutipaijit et al., 2011; Rengel, 1992). As a consequence of ion imbalance and hyperosmotic stresses which are primary effects of salt stress, secondary stresses such as oxidative damage may occur. Limited CO2 fixation due to stress conditions leads to a decrease in carbon reduction by the Calvin cycle and to a decrease in oxidized NADP+ to serve as an electron acceptor in photosynthesis. When ferredoxin is over reduced during photosynthetic electron transfer, electrons may be transferred from PS-I to oxygen to form superoxide radicals (∙O‾2) by the process called Mehler reaction which triggers chain reactions that generate more aggressive oxygen radicals (Heldt, 1997; Hsu and Kao, 2003). It is already known that these cytotoxic Active Oxygen Species (AOS) which are also generated during metabolic processes in the mitochondria, chloroplast and peroxisomes, can destroy normal metabolism through oxidative damage of lipids, proteins and nucleic acids. Lipid peroxidation, induced by free radicals, is also important in membrane deterioration (Halliwell, 1987; McCord, 2000).
Similar to other cereal grains, rice is rich in many bioactive non-nutrient compounds known as antioxidants, including phenolic compounds (Frei and Becker, 2004). There are different types of rice that contain colour pigments, such as purple rice, black rice and red rice. They contain phytochemicals that are responsible for their colours. Generally, these coloured compounds or pigments fall into a number of large groups such as chlorophylls, riboflavin, carotenoids, flavonoids and quinones. Most of these pigments are reported to form in plant for vital functions (Chiang et al., 2006). Phenolic compounds are a large group of naturally occurring and chemically diverse substances, universally distributed in the plant kingdom as secondary metabolic products with a large range of structures and functions (Harbone, 1980). They are generally characterized by an aromatic ring bearing one or more hydroxyl substituents (Martinez-Valverde et al., 2000; Liu, 2007) and include alkaloids, flavonoids, terpenoids and glycosides (Appel, 1993) and are primarily synthesized through the shikimate/chorizmate or succinyl benzoate metabolic pathway. Evidence indicates that phenolic compounds have potent antioxidant properties and free radical scavenging capabilities (Shahidi et al., 1992). They are widely distributed in medicinal plants, spices, vegetables, fruits, grains, pulses and other seeds (Stratil et al., 2007). They also exhibit a wide range of physiological properties such as anti-allergenic, anti-artherogenic, anti-microbial, antioxidant, antithrombotic, cardioprotective and vasodilatory effects (Puupponen-Pimia et al., 2001). The most important groups of phenolics are flavonoids, phenolic acids and polyphenols (King and Young, 1999). Phenolic acids contribute to allelopathic expression in numerous crops, including sorghum, wheat, oat and rice.
Flavonoids are one group of phenolics which consists of two aromatic rings linked by 3 carbons that are usually in an oxygenated heterocycle ring with one or more hydroxyl groups that may be variously elaborated with methyl, methoxyl, amino or glycosyl groups (Liu, 2004, 2007). Anthocyanins are a group of reddish to purple water soluble flavonoids that are the primary pigments in the red and black grains and have been widely identified and characterized in cereal grains (Abdel-Aal et al., 2006). There have been a few reports on the characterization of other flavonoids such as flavonols, flavones, flavanols and flavanones and these phenolic compounds are also known as antioxidants (Adom and Liu, 2002; Hu et al., 2003; Abdel-Aal et al., 2006) and have long been recognized to have protective functions against oxidative damage and are associated with reduced risk of chronic diseases (Adom and Liu, 2002; Liu, 2007). Other phytochemicals such as carotenoids, tocols and oryzanols are also antioxidants (Xu et al., 2001; Aguilar-Garcia et al., 2007; Choi et al., 2007).
Plants have the ability to synthesize a large number of aromatic substances, most of which are phenols or their oxygen-substituted derivatives (Cowan, 1999). Most are secondary metabolites which may be in the form of simple phenols and phenolic acids, quinones, flavones, flavonoids, flavonols, tannins, terpenoids, essential oils and alkaloids. Some such as terpenoids give plants their odours and flavour; many compounds like tannins and quinones are responsible for plant pigments and others such as some alkaloids may be toxic to the consumer. In many instances, these substances serve as plant defence mechanisms against predation by insects, herbivores and microorganisms (Williams and Harborne, 1989; Cowan, 1999; Beckman, 2000). Different studies show that there are often large increases in phenolic synthesis in plants after attack by plant pathogens (De Ascensao and Dubrey, 2003). In resistant plants, phenolic based defence responses are characterized by the early and rapid accumulation of phenolics at the infection site resulting in the effective isolation of the pathogen. These physical responses may include the elaboration of cell wall thickenings and appositions such as papillae, as well as the occlusion of plant vessels.
The concentration of a particular phenolic compound within a plant tissue is dependent on season and may also vary at different stages of growth and development (Lynn and Chang, 1990; Thomas and Ravindra, 1999; Ozyigit et al., 2007). Several internal and external factors, including trauma, wounding, drought and pathogen attack affect the synthesis and accumulation of phenolics. Furthermore, the biosynthesis of phenolics in chloroplasts and their accumulation in vacuoles are enhanced on exposure to light (Kefeli et al., 2003). Photoinhibition and nutrient stresses such as deficiencies in nitrogen, phosphate, potassium, sulphur, magnesium, boron and iron, also trigger the synthesis of phenyl propanoid compounds in some plant species (Dixon and Paiva, 1995; Hollman and Katan, 1999; Balasundram et al., 2006). Studies have also shown that in rice, induction of salt stress resulted in increased synthesis of phenolics and other antioxidants. The potential of induction of antioxidant production through induction of salt stress in rice has also been reported (Daiponmak et al., 2010; Umnajkitikorn et al., 2013). Under the above circumstances, the present study has been carried out to assess the variation in the accumulation of phenolics in some native rice cultivars of North Kerala, India in response to induction of salinity stress. Incremental salinity stress has been applied mimicking the pattern of variation in the salinity levels of the salinity prone rice habitats of the study area.
MATERIALS AND METHODS
Plant materials and planting method: The experiment was conducted in the experimental rainout poly house of Department of Botany, University of Calicut, Kerala, India located at 11°35̍ N latitude and 75°48'E longitude in the first crop season of 2013. Seven native cultivars of rice including five cultivars collected from one of the saline rice habitats of Kerala namely Orthadian, Orkazhama, Kuthiru, Kuttusan and Chovvarian and two native rice cultivars collected from one of the non-saline rice habitats of Kerala namely Kunhutty and Veliyan were used for the study.
| Table 1: | Salinity treatment details |
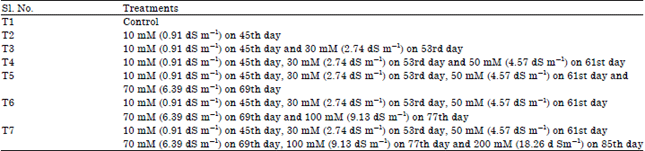 | |
Healthy mature seeds from single plant each were collected and washed in running tap water to remove infected and unfilled grains and dust particles. The seeds were soaked in distilled water and allowed to germinate in 10 cm diameter Petri dishes covered with lid under room temperature. The water was changed every day. The seeds started to germinate from the third day.
On the 10th day, required numbers of the germinated seedlings were transferred to coloured plastic pots of 25 cm diameter filled with paddy soil mixed with enriched compost in 3:1 ratio. Two seedlings were initially planted per pot and after the establishment of the seedlings the smaller among the two were removed. The plants were maintained in the experimental poly house under wetland conditions, always maintaining 3 cm of water above the soil level. The soil was fertilized with 1 g N: P: K = 18: 18: 18 per pot at fortnightly intervals starting from the 30th day. Weeding was done manually whenever required. Plants were grown in Randomized Block Design with three replications.
Experimental treatments: The experimental treatment was started from the 45th day of germination onwards starting from 10 mM (0.91 dS m–1) to 200 mM (18.26 dS m–1) aqueous solution of sodium chloride as detailed in Table 1.
Determination of total phenolic content: The total phenolic content of leaf was determined using the Folin-Ciocalteu reagent (Singleton and Rossi Jr., 1965; Osawa and Namiki, 1981; Gaxiola et al., 2001). Leaf samples (0.1 g) were weighed separately (Sartorius, Germany), homogenized in 80% aqueous ethanol (Merck, India) at room temperature using a clean mortar and pestle, centrifuged in refrigerated centrifuge (Sigma, Germany) at 4°C at a speed of 10,000×g for 20 min and the supernatant was saved. The residue was re-extracted twice with 80% ethanol and supernatants were pooled, put into evaporating dishes and evaporated to dryness at room temperature. Residue was re-dissolved in 5 mL of distilled water. Hundred microliter of this extract was diluted to 3 mL with water and 0.5 mL of freshly prepared Folin-Ciocalteu reagent (Merck, India) was added. After 3 min 2 mL of 20% of sodium carbonate (w/v) (Himedia, India) was added and the contents were mixed thoroughly. The colour was developed and absorbance measured at 650 nm in a spectrometer (Thermo Scientific, USA) after 60 min using catechol as standard. The results were expressed as mg catechol/g of fresh weight of the material. The reaction was conducted in triplicate and results were averaged.
RESULTS AND DISCUSSION
From the results obtained, it is clear that total phenol content got gradually increased with progressing salt stress conditions.
| Table 2: | Variation in total phenolic content in the different rice cultivars studied (mg catechol g–1 fresh weight) |
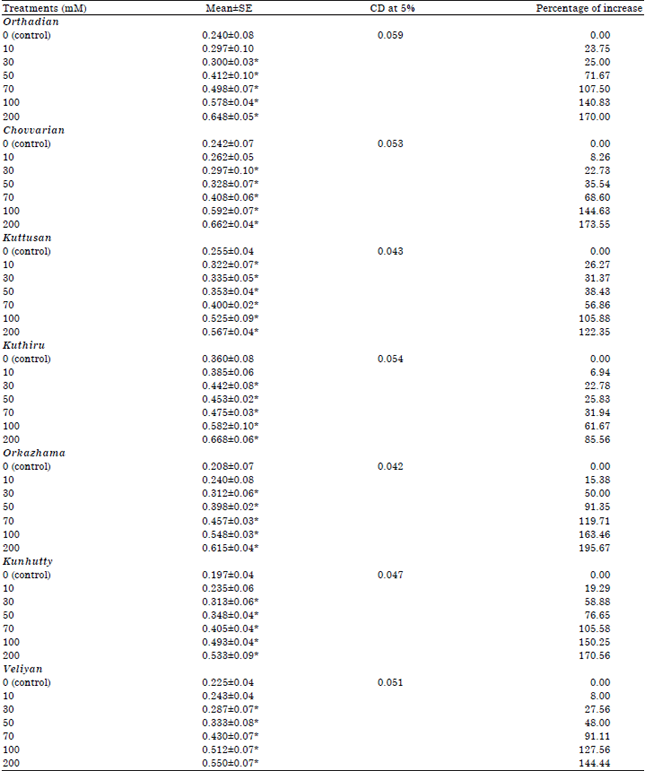 | |
| *Significant variation from the control at 5% | |
The data revealed that Orkazhama, a salinity tolerant rice cultivar from the saline rice tracts of North Kerala showed the highest percentage of increase in total phenolic content (195.67%) (Table 2 and Fig. 1-2).
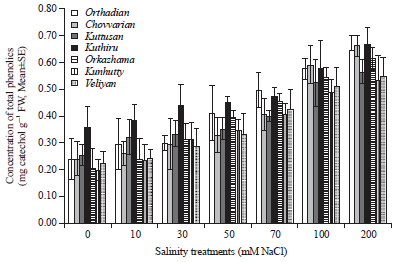 | |
| Fig. 1: | Graph showing the variation in total phenolics in different rice cultivars studied under different levels of salinity treatments |
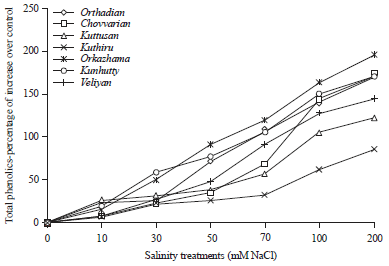 | |
| Fig. 2: | Graph showing the percentage of increase in the total phenolic content in different rice cultivars under salinity stress |
Kuthiru showed the lowest percentage of increase in total phenolic content over the control plants (85.56%). But the highest phenolic content was observed in Kuthiru followed by Chovvarian and Orthadian, the cultivars collected from the traditional saline rice tract. Phenolic content of all the cultivars increased progressively in relation to increase in salt stress and in all the cultivars the highest phenolic content was produced under the highest salt stress induced. All the varieties showed the highest phenolic content under the highest salt stress applied (200 mM NaCl). Among the cultivars studied, Kuttusan showed significant increase in phenolic content over the control starting from 10 mM salt treatment itself, while all the others showed significant variation from 30 mM salt concentration onwards.
A wide array of environmental stresses seriously affect plant growth and crop yield and among that salt stress is one of the major abiotic factor that limit rice productivity. High salt content increases osmotic stress (Mohanty et al., 2002) and leads to the accumulation of excess sodium in plants (Niu et al., 1995; Zhu, 2001). In response to environmental stresses, plants accumulate more soluble osmotic molecules such as proline, betaine and soluble sugar as osmoprotectants (Zhang et al., 2004). They accumulate phenolics and other osmoprotectants also (Daiponmak et al., 2010; Umnajkitikorn et al., 2013). Here, we observed greater accumulation of phenolic compounds in the leaf tissue of all the rice cultivars studied under salt stress. This study provides a clear idea on the accumulation of phenolic compounds due to salinity stress in the rice cultivars that are cultivated in the saline as well as non-saline rice tracts of North Kerala. The cultivars collected from both the saline and non-saline rice tracts showed gradual increase in the accumulation of total phenolics in response to increase in salt stress. The cultivars showed differential variation in the accumulation of total phenolics in relation to salt stress showing that the character was cultivar specific. Among the cultivars studied, Kuthiru showed the highest quantity of phenolic compounds and Orkazhama showed the highest percentage of increase of phenolics over the control plants in relation to increase in salt stress.
Besides ionic and osmotic effects which are the primary stresses of salinity, high salt concentrations additionally induce secondary stresses such as oxidative stress due to the excessive generation of ROS in plants (Zhu, 2001; Vaidyanathan et al., 2003; Kaniuga, 2008; Xu et al., 2011). Under various stresses, ROS production increased quickly in chloroplasts, resulting in damage to the structure of plant cells (Gechev et al., 2006). The ROS are very small molecules and they are highly reactive due to the presence of unpaired valence shell electrons. When plants are exposed to abiotic stresses, their ROS levels can increase dramatically, leading to lipid peroxidation, DNA damage, denaturation of proteins and metabolic disorders. The ROS degrade polyunsaturated lipids and subsequently generate MDA which causes toxic stress in cells by forming covalent protein adducts. Furthermore, ROS triggered by various stresses also affect membrane integrity and cell compartmentation in plants. To prevent oxidative damage, plants process both enzymatic mechanisms such as APX and CAT and non-enzymatic mechanisms such as phenolic compounds for ROS scavenging (Mittler, 2002; Vaidyanathan et al., 2003; Gill and Tuteja, 2010). The present result is consistent with Chutipaijit et al. (2009), Daiponmak et al. (2010) and Ghosh et al. (2011), who found that total phenolic content increased in rice seedlings under salinity stress.
The genetic variations among crop plants are useful in providing a valuable resource in the selection of cultivars with desirable traits (Misra and Dwivedi, 2004). Anil et al. (2005) emphasized the importance of identifying the traits that impart salt tolerance to rice lines and advocated that it would have significant agronomic consequences. The results presented make it clear that total phenol content was gradually increased with progressing salt stress in all the native rice cultivars studied with much higher magnitude of increase in phenol contents in them. Moreover, salt stress tolerant cultivars can be further screened for the production of higher quantity of phenolics in response to salt stress with an objective of improving the antioxidant production potential of such cultivars.
REFERENCES
- Abdel-Aal, E.M., J.C. Young and I. Rabalski, 2006. Anthocyanin composition in black, blue, pink, purple and red cereal grains. J. Agric. Food Chem., 54: 4696-4704.
CrossRefPubMedDirect Link - Adom, K.K. and R.H. Liu, 2002. Antioxidant activity of grains. J. Agric. Food Chem., 50: 6182-6187.
CrossRefDirect Link - Aguilar-Garcia, C., G. Gavino, M. Baragano-Mosqueda, P. Hevia and V.C. Gavino, 2007. Correlation of tocopherol, tocotrienol, γ-oryzanol and total polyphenol content in rice bran with different antioxidant capacity assays. Food Chem., 102: 1228-1232.
CrossRefDirect Link - Anil, V.S., P. Krishnamurthy, S. Kuruvilla, K. Sucharitha, G. Thomas and M.K. Mathew, 2005. Regulation of the uptake and distribution of Na+ in shoots of rice (Oryza sativa) variety Pokkali: Role of Ca2+ in salt tolerance response. Physiologia Plantarum, 124: 451-464.
CrossRefDirect Link - Appel, H.M., 1993. Phenolics in ecological interactions: The importance of oxidation. J. Chem. Ecol., 19: 1521-1552.
CrossRefDirect Link - Asamarai, A.M., P.B. Addis, R.J. Epley and T.P. Krick, 1996. Wild rice hull antioxidants. J. Agric. Food Chem., 44: 126-130.
CrossRefDirect Link - Balasundram, N., K. Sundram and S. Samman, 2006. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem., 99: 191-203.
CrossRefDirect Link - Beckman, C.H., 2000. Phenolic-storing cells: Keys to programmed cell death and periderm formation in wilt disease resistance and in general defence responses in plants? Physiol. Mol. Plant Pathol., 57: 101-110.
CrossRefDirect Link - Bouis, H.E., B.M. Chassy and J.O. Ochanda, 2003. Genetically modified food crops and their contribution to human nutrition and food quality. Trends Food Sci. Technol., 14: 191-209.
CrossRefDirect Link - Chiang, A.N., H.L. Wu, H.I. Yeh, C.S. Chu, H.C. Lin and W.C. Lee, 2006. Antioxidant effects of black rice extract through the induction of superoxide dismutase and catalase activities. Lipids, 41: 797-803.
CrossRefDirect Link - Choi, Y., H.S. Jeong and J. Lee, 2007. Antioxidant activity of methanolic extracts from some grains consumed in Korea. Food Chem., 103: 130-138.
CrossRefDirect Link - Chutipaijit, S., S. Cha-Um and K. Sompornpailin, 2009. Differential accumulations of proline and flavonoids in indica rice varieties against salinity. Pak. J. Bot., 41: 2497-2506.
Direct Link - Conforti, P. and A. Sarris, 2011. Challenges and Policies for the World Agricultural and Food Economy in the 2015 Perspective. In: Looking Ahead in World Food and Agriculture: Perspectives to 2050, Conforti, P. (Ed.). Chapter 12, Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN: 978-92-5-106903-5, pp: 509-539.
Direct Link - Cowan, M.M., 1999. Plant products as antimicrobial agents. Clin. Microbiol. Rev., 12: 564-582.
CrossRefPubMedDirect Link - Daiponmak, W., P. Theerakulpisut, P. Thanonkao, A. Vanavichit and P. Prathepha, 2010. Changes of anthocyanin cyanidin-3-glucoside content and antioxidant activity in Thai rice varieties under salinity stress. ScienceAsia, 36: 286-291.
CrossRefDirect Link - Dash, M. and S.K. Panda, 2001. Salt stress induced changes in growth and enzyme activities in germinating Phaseolus mungo seeds. Biologia Plantarum, 44: 587-589.
CrossRefDirect Link - De Ascensao, A.R.F.D.C. and I.A. Dubery, 2003. Soluble and wall-bound phenolics and phenolic polymers in Musa acuminata roots exposed to elicitors from Fusarium oxysporum f.sp. cubense. Phytochemistry, 63: 679-686.
CrossRefDirect Link - Dixon, R.A. and N.L. Paiva, 1995. Stress-induced phenylpropanoid metabolism. Plant Cell, 7: 1085-1097.
Direct Link - Frei, M. and K. Becker, 2004. Agro-biodiversity in subsistence-oriented farming systems in a Philippine upland region: Nutritional considerations. Biodivers. Conserv., 13: 1591-1610.
CrossRefDirect Link - Gaxiola, R.A., J. Li, S. Undurraga, L.M. Dang, G.J. Allen, S.L. Alper and G.R. Fink, 2001. Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc. Natl. Acad. Sci. USA. 98: 11444-11449.
CrossRefDirect Link - Gechev, T.S., F. van Breusegem, J.M. Stone, I. Denev and C. Laloi, 2006. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. BioEssays, 28: 1091-1101.
CrossRefPubMedDirect Link - Ghosh, S.K.B., A. Missra and D.S. Gilmour, 2011. Negative elongation factor, NELF, accelerates the rate at which heat shock genes are shut-off by facilitating dissociation of heat shock factor. Mol. Cell. Biol., 31: 4232-4243.
CrossRefDirect Link - Gill, S.S. and N. Tuteja, 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem., 48: 909-930.
CrossRefDirect Link - Halliwell, B., 1987. Oxidative damage, lipid peroxidation and antioxidant protection in chloroplasts. Chem. Phys. Lipids, 44: 327-340.
CrossRefDirect Link - Hollman, P.C.H. and M.B. Katan, 1999. Dietary flavonoids: Intake, health effects and bioavailability. Food Chem. Toxicol., 37: 937-942.
CrossRefPubMedDirect Link - Hsu, S.Y. and C.H. Kao, 2003. Differential effect of sorbitol and polyethylene glycol on antioxidant enzymes in rice leaves. Plant Growth Regul., 39: 83-90.
CrossRefDirect Link - Hu, C., J. Zawistowski, W. Ling and D.D. Kitts, 2003. Black rice (Oryza sativa L. indica) pigmented fraction suppresses both reactive oxygen species and nitric oxide in chemical and biological model systems. J. Agric. Food Chem, 51: 5271-5277.
CrossRefPubMedDirect Link - Kaniuga, Z., 2008. Chilling response of plants: Importance of galactolipase, free fatty acids and free radicals. Plant Biol., 10: 171-184.
CrossRefDirect Link - King, A. and G. Young, 1999. Characteristics and occurrence of phenolic phytochemicals. J. Am. Dietetic Assoc., 99: 213-218.
CrossRefPubMedDirect Link - Kefeli, V.I., M.V. Kalevitch and B. Borsari, 2003. Phenolic cycle in plants and environment. J. Cell Mol. Biol., 2: 13-18.
Direct Link - Liu, R.H., 2004. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr., 134: 3479S-3485S.
PubMedDirect Link - Liu, R.H., 2007. Whole grain phytochemicals and health. J. Cereal Sci., 46: 207-219.
CrossRefDirect Link - Lynn, D.G. and M. Chang, 1990. Phenolic signals in cohabitation: Implications for plant development. Annu. Rev. Plant Physiol. Plant Mol. Biol., 41: 497-526.
CrossRefDirect Link - Martinez-Valverde, I., M.J. Periago and G. Ros, 2000. [Nutritional importance of phenolic compounds in the diet]. Archivos Latinoamericanos Nutricion, 50: 5-18, (In Spanish).
PubMed - McCord, J.M., 2000. The evolution of free radicals and oxidative stress. Am. J. Med., 108: 652-659.
CrossRefDirect Link - McKersie, B.D. and Y.Y. Leshem, 1994. Stress and Stress Coping in Cultivated Plants. Kluwer Academic Publishes, Dordrecht, The Netherlands, ISBN-13: 978-0792328278, Pages: 256.
Direct Link - Misra, N. and U.N. Dwivedi, 2004. Genotypic difference in salinity tolerance of green gram cultivars. Plant Sci., 166: 1135-1142.
CrossRefDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Mohanty, A., H. Kathuria, A. Ferjani, A. Sakamoto, P. Mohanty, N. Murata and A. Tyagi, 2002. Transgenics of an elite indica rice variety pusa basmati 1 harbouring the codA gene are highly tolerant to salt stress. Theoret. Applied Genet., 106: 51-57.
CrossRefDirect Link - Niu, X., R.A. Bressan, P.M. Hasegawa and J.M. Pardo, 1995. Ion homeostasis in NaCl stress environments. Plant Physiol., 109: 735-742.
PubMedDirect Link - Osawa, T. and M. Namiki, 1981. A novel type of antioxidant isolated from leaf wax of Eucalyptus leaves. Agric. Biol. Chem., 45: 735-739.
Direct Link - Osawa, T., N. Ramarathnam, S. Kawakishi and M. Namiki, 1992. Antioxidative Defence Systems Generated by Phenolic Plant Constituents. In: Phenolic Compounds in Food and Their Effects on Health. II. Antioxidants and Cancer Prevention, Huang, M.T., C.T. Ho and C.Y. Lee (Eds.). ACS Press, Washington, DC., pp: 122-143.
- Ozyigit, I.I., M.V. Kahraman and O. Ercan, 2007. Relation between explant age, total phenols and regeneration response in tissue cultured cotton (Gossypium hirsutum L.). Afr. J. Biotechnol., 6: 3-8.
Direct Link - Puupponen-Pimia, R., L. Nohynek, C. Meier, M. Kahkonen, M. Heinonen, A. Hopia and K.M. Oksman-Caldentey, 2001. Antimicrobial properties of phenolic compounds from berries. J. Applied Microbiol., 90: 494-507.
CrossRefPubMedDirect Link - Rengel, Z., 1992. The role of calcium in salt toxicity. Plant Cell Environ., 15: 625-632.
CrossRefDirect Link - Salekdeh, G.H., J. Siopongco, L.J. Wade, B. Ghareyazie and J. Bennett, 2002. A proteomic approach to analyzing drought- and salt-responsiveness in rice. Field Crops Res., 76: 199-219.
CrossRefDirect Link - Shahidi, F., P.K. Janitha and P.D. Wanasundara, 1992. Phenolic antioxidants. Crit. Rev. Food Sci. Nutr., 32: 67-103.
CrossRefDirect Link - Singleton, V.L. and J.A. Rossi, 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic., 16: 144-158.
CrossRefDirect Link - Stratil, P., B. Klejdus and V. Kuban, 2007. Determination of phenolic compounds and their antioxidant activity in fruits and cereals. Talanta, 71: 1741-1751.
CrossRefDirect Link - Chutipaijit, S., S. Cha-Um and K. Sompornpailin, 2011. High contents of proline and anthocyanin increase protective response to salinity in Oryza sativa L. spp. indica. Aust. J. Crop Sci., 5: 1191-1198.
Direct Link - Umnajkitikorn, K., B. Faiyue and K. Saengil, 2013. Enhancing antioxidant properties of germinated Thai rice (Oryza sativa L.) cv. Kum Doi Saket with salinity. J. Rice Res., Vol. 1.
Direct Link - Thomas, P. and M.B. Ravindra, 1999. Shoot tip culture in mango: Influence of medium, genotype, explant factors, season and decontamination treatments on phenolic exudation, explant survival and axenic culture establishment. J. Hortic. Sci. Biotechnol., 72: 713-722.
Direct Link - Vaidyanathan, H., P. Sivakumar, R. Chakrabarty and G. Thomas, 2003. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)-differential response in salt-tolerant and sensitive varieties. Plant Sci., 165: 1411-1418.
CrossRefDirect Link - Xu, Z., N. Hua and J.S. Godber, 2001. Antioxidant activity of tocopherols, tocotrienols and γ-oryzanol components from rice bran against cholesterol oxidation accelerated by 2,2'-azobis(2-methylpropionamidine) dihydrochloride. J. Agric. Food Chem., 49: 2077-2081.
CrossRefPubMedDirect Link - Xu, X.Y., R. Fan, R. Zheng, C.M. Li and D.Y. Yu, 2011. Proteomic analysis of seed germination under salt stress in soybeans. J. Zhejiang Univ. Sci. B, 12: 507-517.
CrossRefDirect Link - Zhai, C.K., C.M. Lu, X.Q. Zhang, G.J. Sun and K.J. Lorenz, 2001. Comparative study on nutritional value of Chinese and North American wild rice. J. Food Compos. Anal., 14: 371-382.
CrossRefDirect Link - Zhang, J.Z., R.A. Creelman and J.K. Zhu, 2004. From laboratory to field. Using information from Arabidopsis to engineer salt, cold and drought tolerance in crops. Plant Physiol., 135: 615-621.
Direct Link - Zhou, Z., K. Robards, S. Helliwell and C. Blanchard, 2002. Ageing of stored rice: Changes in chemical and physical attributes. J. Cereal Sci., 35: 65-78.
CrossRefDirect Link