
Jamsari
Plant Breeding Section, Faculty of Agriculture, Andalas University, 25163, Padang, West Sumatera, Indonesia
Lily Syukriani
Plant Breeding Section, Faculty of Agriculture, Andalas University, 25163, Padang, West Sumatera, Indonesia
Hashlin Pascananda Utami
Plant Breeding Section, Faculty of Agriculture, Andalas University, 25163, Padang, West Sumatera, Indonesia
Friedrich Herberg
Department of Biochemistry, Kassel University, Heinrich-Plett Str. 40; 34132, Building AVZ I, 2.Stock, Kassel, Germany
Wolfgang Nellen
Department of Genetics, Kassel University, Heinrich-Plett Str. 40; 34132, Kassel, Germany
Istino Ferita
Plant Breeding Section, Faculty of Agriculture, Andalas University, 25163, Padang, West Sumatera, Indonesia
Asian Journal of Agricultural Research
Year: 2015 | Volume: 9 | Issue: 1 | Page No.: 23-32







ABSTRACT
Artificial inoculation is an important method for understanding the plant-virus interaction. Currently available techniques for virus artificial infection are whitefly based inoculation, grafting, mechanical, agroinoculation and particle bombardment. In fact, almost all the available inoculation methods do not result in ideal efficiency nor practicality. The purpose of this study is to establish an injection based technique for routine artificial inoculation of Geminivirus particle into chilli pepper plant. The following treatments were applied in this study: Different ratios of extract-buffer (0, 20 and 60% w/v) and injection at four different positions (no inoculation, shoot, middle of main vein and petiol). The presence of Geminivirus particles in plants whether for inoculum source and infection effectivity was confirmed using the polymerase chain reaction. Some morphological observations were also applied in measurement of infection effectivity. Overall, the incidence of infection was 18% and among these, the average intensity of disease was 53% in the four week post inoculation. Geminivirus transmission by injection technique described in this study could be a simple and effective method for geminivirus artificial infection. However, accuracy, simplicity and uniformity have to be accommodated for further application of this technique.
PDF Abstract XML References Citation
Received: November 12, 2014;
Accepted: December 31, 2014;
Published: February 21, 2015
How to cite this article
Jamsari, Lily Syukriani, Hashlin Pascananda Utami, Friedrich Herberg, Wolfgang Nellen and Istino Ferita, 2015. Injection Technique Could as a New Promising Method for Artificial Infection of Geminivirus Particles in Chili Pepper (Capsicum annuum L.). Asian Journal of Agricultural Research, 9: 23-32.
DOI: 10.3923/ajar.2015.23.32
URL: https://scialert.net/abstract/?doi=ajar.2015.23.32
DOI: 10.3923/ajar.2015.23.32
URL: https://scialert.net/abstract/?doi=ajar.2015.23.32
INTRODUCTION
Geminivirus is one of the causal agents leading to significant losses of important crops in many tropical and sub tropical countries worldwide (Varma and Malathi, 2003). In chilli-pepper cultivation, Geminivirus is believed to be the only agent causing Pepper yellow leaf curl disease (PepYLCD) (Jamsari and Pedri, 2013). Despite intensive breeding efforts covering vector and geminiviruses, no satisfactory results to achieve resistance have so far been published.
Until now, studies on the biological characterization of Geminivirus, their host range and their transmission mode have been extensively undertaken but applications are scarce. With increased spreading of the disease, studies on virus transmission mechanism have become more and more important. This includes investigations on plant-virus interaction and particularly virus resistance. To better understand the mechanisms of virus infection, a reliable artificial virus particle infection method must be established.
So far, methods for artificial virus particle infection have been applied, e.g., grafting (Bos, 1967; Palauqui et al., 1997; Ali et al., 2013), mechanical infection via wounding (Mayo et al., 2000), agroinoculation (Leiser et al., 1992; Mutterer et al., 1999), particle bombardment (Lapidot et al., 2007) and insect based vector using Bemisia tabaci for artificial infection (Inoue-Nagata et al., 2007).
Grafting methods have shown a 71.4% effectivity with incubation periods ranging from 20-29 days. Bemisi tabaci based artificial infection could produce up to 80% infection effectivity. However both methods are not reliable, since effectivity ranges from only 12-100% (Ganefianti et al., 2008). Effectivity by mechanical infection via wounding was in general very low (Honda et al., 1983).
Grafting is rather impractical, since this method requires specific expertise for preparing stocks and scions. The B. tabaci based artificial infection is tedious, since it requires preparation of insect vectors and it carries the potential danger of uncontrolled release of the viruliferous vectors into the environment. Therefore, B. tabaci based artificial infection can only be applied in specific biological safety facilities. Particle bombardment can transmit Geminivirus particles efficiently and uniformly, but requires costly equipment and can therefore not be used in many laboratories. Agroinfection is also tedious and requires special skills. Therefore, a simple and efficient alternative would be of great advantage. In this study, an injection method was described for artificial virus particle inoculation for chilli pepper which circumvents most of the problems described above.
MATERIALS AND METHODS
Plant sources inoculum: The inoculum source was originally chosen from Kopay chilli plants grown by farmers on the chilli cultivation field in Payakumbuh Regency West Sumatera-Indonesia. The choice was based on the morphological symptoms showing curling, crumpled, wrinkled, yellowing, mosaic leafs and stunted branch and was confirmed by PCR. The chosen plants were further maintained isolated in the greenhouse during the study.
Plant test material: A similar genotype as the inoculum source, namely Kopay was used in this study. Preparation of plant material to be infected was performed in an insect shielded green house. Kopay chilly seeds were treated with sterile distilled water and then sowed out in perforated plastic trays filled with sterilized soil. Two weeks old seedlings were transferred to polybags containing sterilized combination media of soil and manure at the ratio of 2:1. The seedlings were kept for further two weeks before treatment was applied. For ensuring their isolation from any insect infection each individual plant was further grown separately and maintained in an insect-proof box. Light intensity in the greenhouse during study was between 1500-2500 lux, with 29,4% relative humidity. Meanwhile, temperature was kept between 27-30°C at day and 22-24°C at night (Honda et al., 1983). The temperature condition was also maintained during incubation period after artificial infection was performed.
Geminivirus particle detection: The presence of Geminivirus particle both in the inoculum sources and artificially infected plants was observed morphologically and verified by PCR. DNA was isolated by the procedure of Doyle and Doyle (1987) and subsequently amplified as described by Rojas et al. (1993). PCR amplification was performed with the universal primers PAL1V1978 (5'-GCATCTGCAGGCCCACATYGTCTTYCCNGT-3') and PAR1C715 (5'-GATTTCTGCAGTTDATRTTYTCRTCCATCCA-3'). Successful PCR amplification was shown by the presence of an about 1,600 bp single DNA fragment.
Preparation of Geminivirus particles and injection spot: Geminivirus particles were extracted from inoculated leafs, showing yellowish and curling symptoms. The infected leaves were homogenised in a mortar with potassium phosphate buffer (0.1 M, pH 8) containing 0.1% β-mercaptoethanol, 0.2 M KH2PO4, 0.2 M K2HPO4. β-mercaptoethanol was added freshly prior to use. The extract was filtered through sterilized gauze before injection. Injection was done with sterile syringe and a 0.5 mm diameter needle. The injected volume of extract was 2 μL.
Treatment used in the study: Two variables were used in this study: The extract ratio and the injection spot position. Extract ratio was: (1) Mock (containing only phosphate buffer) (A0), (2) Ratio of 20% (w/v) of inoculum leaf and phosphate buffer (A1) and ratio of 60% (w/v) of inoculum leaf and phosphate buffer (A2). The second factor was injection at four positions, (1) Mock (B0), (2) Shoot tip (B1), (3) Middle of vein leaf (B2) and (4) Petiole of upper leaf (B3). All treatments were done in five replicates.
Analysis of artificial infection effectivity: The first indicator for infection effectivity was on the appearance of morphological symptoms. Leaves of infected plants showing curling and yellowish colour were assumed as “Infected”. Verification was also done by PCR technique using the primer pair PAL1V1978 and PAR1C715 (Rojas et al., 1993).
RESULTS AND DISCUSSION
The PCR analysis by using primer pair PAL1V1978 and PAR1C715 as described by Rojas et al. (1993) successfully detected Geminivirus particles in the inoculums source as a 1,600 bp PCR product (Fig. 1a-b).
Morphological symptoms: Effect of injection was observed one Week Post Inoculation (WPI). Observation was based on leaf morphological symptom and recorded by photographic documentation. Selected examples are shown in Fig. 2. All leaves showing virus symptoms were confirmed by PCR as described above.
One week post inoculation, 60% in average (3 plants out of 5) from each treatment of A1B2, A1B3 showed morphological abnormality in leaves. The virus infection symptom was indicated by the yellowish spots formed on 70-80% of the upper leaf (mosaic), curling, abnormal leaf shape, crumpled, smaller buds and leaf edge was curved upward (cupping). The other treated plants showed normal leaf growth as compared to control plants (A0B0).
Observation in the second week post inoculation, 60% (3 plants out of 5 plants) of A2B1 showed leaf abnormalities. About 33% (4 out of 12 leafs) from every single plant of this treatment showed curling, more crumpled and intensive cupping starting from the upper leaves (buds). Mosaic yellowish colour of the abnormal leaves was not observed in this two week observation. Furthermore treatment A1B2 and A1B3 showed more advanced leaf abnormality, indicated by complete curling, crumpled and cupping with some mosaic yellowish spots in some upper leaves. In the second week post inoculation almost all (100%) of the leaves from all infected plants in A1B2 and A1B3 population showed leaf abnormality.
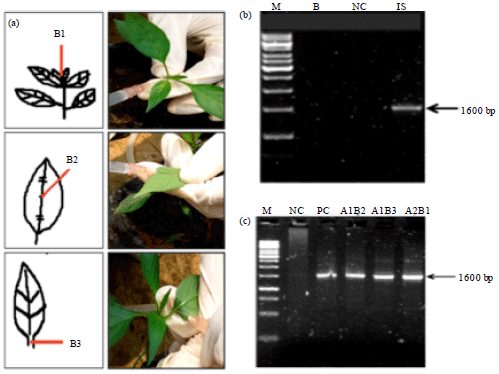 | |
| Fig. 1(a-c): | (a) Injection spot position and (b-c) Inoculum verification and inoculation effectivity via PCR based analysis. Figure shows the position for injection, B1: Shoot tip, B2: Middle of vein leaf and B3: Petiole of upper leaf, (b) PCR analysis shows the 1600 bp PCR product, indicating Geminivirus particle in inoculum sample, (c) Electropherogram of PCR products of samples from three successful treatments. M = 1 kb ladder, B: Blank, NC = Negative control, PC = Positive control, IS: Inoculum source. Treatment A1B2, A1B3 and A2B1 are putative successfully infected samples |
To verify infection effectivity, PCR was applied in all experimental units. Selected, representative results are shown in Fig. 1c. The three putative infected samples A1B2, A1B3 and A2B1 produced a single 1,600 bp fragment, whereas the other four treated plants including mocked samples did not produce any fragment. The 1,600 bp fragment from the three treatments indicated the presence of virus particles in the sample as also obtained from the positive control of the inoculum source.
Symptoms that appeared from A1B2 and A1B3 treatments were almost similar and showed more systemic spreading compared with A2B1. This characteristic was seen until the end of the observation period. The symptoms shown by A2B1 treatment tended to be more local but showed more intensive effects. The most affected leaves are in the surrounding of the injection spot. It was believed that the reason for differences in spreading of symptoms was caused by different characteristics of the tissues, in which was still meristematic and actively dividing in A2B1. On the other hand, the tissues in A1B2 and A1B3 were completely differentiated tissues. Moreover, these tissues were already taking part in photosynthesis. Leaves where injection was applied at a “Source” of photosynthetic products may distribute virus particles through the phloem vascular tissue. In contrast, shoots do not yet have the capacity as a “Source” and play a reduced role in spreading of Geminivirus particles through the vascular transportation system.
 | |
| Fig. 2: | Development of leaf symptoms from the different treatments during 4 weeks post inoculation. Circle size represent disease intensity in time frame. A: Ratio of extract, 0 = Control (mock), 1 = 1:5, 2 = 3:5; B: Injection spot position, 0 = No injection, 1 = Shoot, 2 = Midle vein, 3 = Ptiole, WPI: Week Post Inoculation |
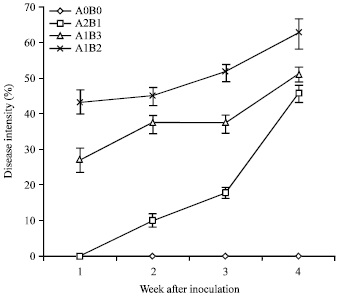 | |
| Fig. 3: | Disease intensity development from three treatments successfully infected by injection |
The argument proposed above is supported by Morra and Petty (2000). They pointed out, that once Geminivirus infects cells they will associate with the vascular system. Matthews (1970) described that virus particles will spread from one cell to the nearest cell which in turn would increase the infection focus. Both this arguments lead to the conclusion that the location of injection influences spreading of symptoms. Moreover, tissue with photosynthetic activity, will increase the probability of symptom spreading through the vascular system. The three top leaves are the most suitable tissues since they still have soft tissues. Furthermore, the leaves are also considered to be the most effective in sunlight energy absorption, so that the metabolic products during the photosynthetic processes are efficiently produced.
Disease intensity: In order to assess the severity of the disease caused by injected virus particle, the disease intensity was calculated during 4 weeks of observation (Fig. 3). Data is based on the average of all successful plants artificially infected by injection. Intensity of the disease was calculated using equation described by Lapidot et al. (2001) as follow:
Where:
| DI | = | Disease intensity |
| Ni | = | Number of infected leaf with i-th category |
| Vi | = | Value of category attack-i |
| Z | = | Value of highest category attack |
| N | = | Number of leaf total analyzed |
The intensity categories are listed at Table 1.
The intensity of the disease development was determined during 4 weeks after treatment and their overall observation was shown in Fig. 3.
| Table 1: | Criterion scale of symptoms severity in pepper yellow leaf curl disease (PepYLCD)* |
 | |
| *After Lapidot et al. (2001) | |
Figure 3 showed that disease intensity from A1B2 and A2B1 tended to increase during 4 weeks after injection while disease intensity of A1B3 remained stable in the second and third week after treatment. The rate of disease intensity development from both A1B3 and A2B1 decreased during the second to third week after injection but again increase progressively from the third to fourth week. Meanwhile, A1B2 showed stable increase in disease intensity development rate from the beginning to the fourth week. The zero rate of disease intensity shown by A1B3 was not due to stopped spreading, but it rather caused by delayed symptoms shown by new emerging young leaves. On the other hand A2B1 showed a progressive disease intensity development from the third to fourth week (27.95%). In this case, the young new emerging leaves showed disease symptoms directly after opening from the bud showing curly, curved, mosaic and dwarf growth features.
The data obtained here is comparable with previous studies describing artificial infection methods reported by Ganefianti et al. (2008). They reached 40% disease intensity 20 days after infection. However, in nature disease intensity caused by whitefly transmission could reach up to 95.2% (Trisno et al., 2009). This is probably because whitefly movement during the feeding period causes multiple infections in a single plant. Moreover, the whitefly stylet containing virus particles directly interacts with photosynthetic products in the targeted phloem. This will increase the probability of disease intensity. Based on this assumption, the most important factor affecting inoculation effectivity depends on the direct interaction between virus particles and the photosynthetic distribution network in the plant vascular system.
Factors affecting viral injection effectivity: Three of six injection treatments (A1B2, A1B3 and A2B1) showed Geminivirus infection symptoms after seven days post inoculation. Treatment combination of A1B2 and A1B3 showed almost similar symptoms compared to A2B1. The three other treatments A1B1, A2B2 and A2B3 showed no symptomatic effect until the end of the observation time. The failure of these treatments to produce symptoms probably could be most due to inconsistency of injection depth applied in this study. Manual injection could not accurately control the depth of needle tip in micrometer scale. That’s way a mechanistic way, that can reach phloem tissue accurately should be developed. Basu and Giri (1993) described some factors that influence infection effectivity, i.e., source of inoculums, inoculums preparation method (extract preparation), stability of the virus particle along preparation steps and environmental conditions such as light intensity and temperature during incubation period. However, such those factors probably are not the case in this study, since they were handled equally.
The use of young symptomatic and active photosynthetic leaves (B2 and B3) as inoculum source showed higher chance of infection success. This is based on the fact that the young symptomatic and active photosynthetic leaves tend to have higher concentration of viral particles, since young leaves are still symptomatic and contain more photosynthetic products. Basu and Giri (1993) described, that the quality of inoculum source is widely dependent on the concentration of virus particles and the physiological state of the plant samples that in turn could affect the concentration of substances that can potentially be a virus inhibitor or inactivator.
Extract preparation is very important to obtain virus particles with a maximum titre. During extraction process of infected leaf tissue, a wide variety of metabolites from disrupted cells and their cellular debris are liberated along with viral particles that exist in the metabolic network. Some of these compounds may lead to inactivation of the virus particle or act as inhibitors in the infection process. This can be overcome by the addition of potassium phosphate buffer solution maintain a stable pH, eliminate the inhibitors and inactivators, as well as inhibit oxidation activity and phenolation. β-Mercaptoethanol helps to prevent the loss of virus particles infectivity caused by oxidation processes.
Leaf tissue, especially young leaves probably are the most sensitive tissues to be inoculated. However, even if the test plants are sensitive to the virus, several factors such as age, nutrition, temperature and light intensity also determines the degree and extent of infection.
The affinity of Geminivirus to the phloem tissue of the host plant might also be one of the reasons for the low success rate of mechanical inoculation. Morra and Petty (2000) described that Begomovirus infection is limited to the phloem tissue of plants. The 38 inoculated tissue sections contained 317 cells which were infected with Begomovirus. Among them 314 cells (about 99%) were in the vascular system while only 3 (~1%) were mesophyll cells. Further analysis of the infected cells associated with the vascular system, showed that 2 were companion cells, 18 were phloem parenchyma cells and 12 of them are bundle-sheath vessels. They did not find any DNA virus in phloem sieve elements. The nature of “Phloem limited” Geminivirus particles complicates the process of mechanical inoculation, in this case virus particles can only replicate in cells surrounding the phloem tissue. After artificial inoculation, virus particle stay in a single epidermal cell, from there the virus particles will spread to nearby cells and increasing the focus of infection (Matthews, 1970).
Rojas et al. (2005) described that only a few of the genus Begomovirus can come out of the vascular system to the surrounding mesophyll tissue in the primary affected tissues or organs infected systemically. Therefore, the accuracy of targeting cells or tissues by injection will significantly affect the effectivity of artificial infection. Injection must be located accurately to the spot where Geminivirus can replicate which is the area surrounding the phloem vascular system. However, so far there is no proper way to control injection due to varying plant tissue characteristic.
The A2B1 was injected with more virus particle (60%) but showed delay symptom (Fig. 2, 3) of virus infection. In the first week post inoculation, no symptom could be observed, just in the second week post inoculation symptoms were apparent and progressed until they almost reached the level of A1B2 and A1B3 after four weeks post inoculation. This result apparently contradicts the common hypothesis that more virus particles, cause higher infection effectivity. The concentration of virus particle thus does not have a linear correlation with the infection effectivity. This phenomenon has been termed as “Bottleneck effect” (Moury et al., 2007). The bottleneck phenomenon is an event where the virus particles that enter into the plant tissue is allowed only in a certain number of the virus particles. French and Stenger (2003) in their study of wheat streak mosaic virus found that only about 10 virus particles could contribute to systemic infection in plants after mechanical inoculation, although a few hundred particles were present at the beginning to initiate an infection. Furthermore, Moury et al. (2007) pointed out that the average aphid can transmit only 20-50% virus population existing in their stylets.
| Table 2: | Comparison of injection method with three existing Geminivirus artificial infection |
 | |
This amount is apparently low when compared to the existing virus populations in an infected plant tissue. Another possibility could be explained by reducing of virus particle persistency. Even though during inoculum extract preparation the A2B1 contained more higher titre of virus particle than A1B2 and A1B3, but in line of steps of extract preparation persistency of virus particle downgraded making incubation time of A2B1 more longer. Moury et al. (2007) even mentioned virulent-avirulent survival where avirulent strains inhibit the virulent ones to propagate.
Finally, comparison of available artificial infection (Table 2) demonstrates that injection based inoculation provides enough disease incidence with short incubation period and is practical. The whitefly based method could successfully provide the disease incidence, allowing highly efficient inoculation with short incubation period. However, maintaining an infectious whitefly population is tedious and has the risk of uncontrollable virus transmission into the environment. This makes whitefly based inoculation non-practical. The grafting method has the disadvantage of long incubation periods and laborious handling. Even though mechanical inoculation is practical, this method has a low efficiency in transmitting virus particles.
CONCLUSION
Based on disease incidence, incubation period and practicality, the injection method provides a promising alternative for Geminivirus artificial infection in chilli pepper compared to the existing available methods. However, more accurate injection into target tissue, has to be achieved for further application of this technique.
ACKNOWLEDGMENT
We gratefully thank the Directorate General of Higher Education from Indonesia Ministry of Education and Culture (DP2M) for supporting this research via scheme of Research for International Collaboration and Publication fiscal year 2014, contract number: 06/UN.16/PL/KLNPI/I/2014. We thank also the German Academic Exchange Service (DAAD) which allowed us to establish and partly fund this collaboration through IGN-TTRC (Indonesia German Network for Teaching, Training and Research Collaboration).
REFERENCES
- Ali, E.M., K. Kobayashi, N. Yamaoka, M. Ishikawa and M. Nishiguchi, 2013. Graft transmission of RNA silencing to non-transgenic scions for conferring virus resistance in Tobacco. PloS ONE, Vol. 8.
CrossRefDirect Link - French, R. and D.C. Stenger, 2003. Evolution of wheat streak mosaic virus: Dynamics of population growth within plants may explain limited variation. Annu. Rev. Phytopathol., 41: 199-214.
CrossRefDirect Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Honda, Y., M. Iwaki, Y. Saito, P. Thongmeearkom, K. Kittisak and N. Deema, 1983. Mechanical transmission, purification and some properties of whitefly-borne mung bean yellow mosaic virus in Thailand. Plant Dis., 67: 801-804.
Direct Link - Inoue-Nagata, A.K., T. Nagata, A.C. de Avila and L.D.B. Giordano, 2007. A reliable begomovirus inoculation method for screening Lycopersicon esculentum lines. Horticultura Brasileira, 25: 447-450.
CrossRefDirect Link - Jamsari, J. and J. Pedri, 2013. Complete nucleotide sequence of DNA A-like genome and DNA-β of monopartite pepper yellow leaf curl virus, a dominant Begomovirus infecting Capsicum annuum in West Sumatera Indonesia. Asian J. Plant Pathol., 7: 1-14.
CrossRefDirect Link - Lapidot, M., G. Weil, L. Cohen, L. Segev and V. Gaba, 2007. Biolistic inoculation of plants with Tomato yellow leaf curl virus DNA. J. Virol. Methods, 144: 143-148.
CrossRefDirect Link - Lapidot, M., M. Friedmann, M. Pilowsky, R. Ben-Joseph and R. Cohen, 2001. Effect of host plant resistance to Tomato Yellow Leaf Curl Virus (TYLCV) on virus acquisition and transmission by its whitefly vector. J. Phytopathol., 91: 1209-1213.
CrossRefDirect Link - Leiser, R.M., V. Ziegler-Graff, A. Reutenauer, E. Herrbach and O. Lemaire et al., 1992. Agroinfection as an alternative to insects for infecting plants with beet western yellows luteovirus. Proc. Natl. Acad. Sci. USA., 89: 9136-9140.
Direct Link - Mayo, M., E. Ryabov, G. Fraser and M. Taliansky, 2000. Mechanical transmission of Potato leafroll virus. J. General Virol., 81: 2791-2795.
Direct Link - Morra, M.R. and I.T. Petty, 2000. Tissue specificity of geminivirus infection is genetically determined. Plant Cell, 12: 2259-2270.
Direct Link - Moury, B., F. Fabre and R. Senoussi, 2007. Estimation of the number of virus particles transmitted by an insect vector. Proc. Natl. Acad. Sci. USA., 104: 17891-17896.
Direct Link - Palauqui, J.C., T. Elmayan, J.M. Pollien and H. Vaucheret, 1997. Systemic acquired silencing: Transgene-specific post-transcriptional silencing is transmitted by grafting from silenced stocks to non-silenced scions. EMBO J., 16: 4738-4745.
CrossRefPubMedDirect Link - Rojas, M.R., R.L. Gilbertson, D.R. Russell and D.P. Maxwell, 1993. Use of degenerate primers in the polymerase chain reaction to detect whitefly-transmitted geminiviruses. Plant Dis., 77: 340-347.
CrossRefDirect Link - Rojas, M.R., C. Hagen, W.J. Lucas and R.L. Gilbertson, 2005. Exploiting chinks in the plant's armor: Evolution and emergence of geminiviruses. Annu. Rev. Phytopathol., 43: 361-394.
CrossRefDirect Link - Trisno, J., S.H. Hidayat, T. Habazar, I. Manti and Jamsari, 2009. Detection and sequence diversity of begomovirus associated with yellow leaf curl disease of pepper (Capsicum annuum) in West Sumatera, Indonesia. Microbiol. Indonesia, 3: 56-61.
Direct Link - Varma, A. and V.G. Malathi, 2003. Emerging geminivirus problems: A serious threat to crop production. Ann. Applied Biol., 142: 145-164.
CrossRefDirect Link