
Rika Fithri N. Buana
Microbiology Division, Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Darmaga Campus, 16680, Bogor, Indonesia
Aris Tri Wahyudi
Microbiology Division, Department of Biology, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, Darmaga Campus, 16680, Bogor, Indonesia
Nurita Toruan-Mathius
Department of Biotechnology, Plant Production and Biotechnology Division, PT. SMART Tbk, Sinarmasland Plaza, Tower II, Jl. M.H. Thamrin No.51 Kav. 22, Jakarta Pusat, 10350, Indonesia
Asian Journal of Agricultural Research
Year: 2014 | Volume: 8 | Issue: 5 | Page No.: 259-268







ABSTRACT
Basal Stem Rot (BSR) disease caused by G. boninense is one of the most serious diseases in oil palm. Many attempts have been done to prevent or reduce infection of this disease but they have not provided optimum results. Burkholderia sp. isolated from rhizosphere and root tissue of symptomless oil palm showed potentials in suppressing G. boninense growth in vitro. Five isolates of rhizosphere Burkholderia sp. and one endophyte Burkholderia sp. were used in antagonist test against G. boninense growth on PDA media. Antifungal biosynthesis related gene from Burkholderia sp. was confirmed by using PCR with pyrrolnitrin (prn), pyoluteorin (plt), phenazine (phz) and DAPG (phl) primers. Burkholderia sp. with the highest antagonist activity were applied to oil palm germinated seed for in vivo test. Endophyte Burkholderia B212 showed the highest antagonist activity against G. boninense growth in vitro with PIRG 34.38%. The B212 genome was yield an expected PCR product by using prn primers (790 bp). Sequence BLAST result showed the gene was 99% identical with B. cepacia partial prnD gene, strain ESR63. In vivo test of B212 showed that treatment of Burkholderia B212 on plant without G. boninense infection increased the height and biomass of the plant. However, B212 did not decrease the disease incidence and disease severity on G. boninense infected plant. In addition it decreased the plant height and biomass compare to control plant.
PDF Abstract XML References Citation
Received: July 22, 2014;
Accepted: October 17, 2014;
Published: October 28, 2014
How to cite this article
Rika Fithri N. Buana, Aris Tri Wahyudi and Nurita Toruan-Mathius, 2014. Control Activity of Potential Antifungal-Producing Burkholderia sp.
in Suppressing Ganoderma boninense Growth in Oil Palm. Asian Journal of Agricultural Research, 8: 259-268.
DOI: 10.3923/ajar.2014.259.268
URL: https://scialert.net/abstract/?doi=ajar.2014.259.268
DOI: 10.3923/ajar.2014.259.268
URL: https://scialert.net/abstract/?doi=ajar.2014.259.268
INTRODUCTION
Oil palm is one of the most important agricultural export crops in Indonesia besides rubber, cocoa, coffee and spices (Stads et al., 2007). The BSR has been a serious threat to the oil palm industry in Indonesia because it shortens the productive life of oil palms and causes serious economic loss. The disease is caused by a white-rot fungi G. boninense and in the past few decades has been spreading rapidly, for instance, in North Sumatra, Indonesia, this disease can lead to losses as much as 50% after repeated planting cycles (25 years) (Corley and Tinker, 2003).
The use of fungicides for fungal control did not produce significant results yet (Haas and Defago, 2005). This may be due to the fact that by the time treatment is applied, the palms may already have the disease. Antifungal-producing bacteria is a promising biocontrol agent (BCA) to overcome this disease. The BCA does not necessarily be a cure for the disease but to slowing or even to stop the disease spread by protecting the plant or enhanceing the plant defense. Endophyte BCAs have more value since, it can live inside the plant tissue through the plants lifetime.
Endophyte bacteria is bacteria which live inside the plant tissues without causing apparent harm or symptoms to the host (Munif et al., 2003). Endophyte as the internal plant habitat provide several advantages as BCA. It will be in less competition with other microorganisms, sufficient supply with the nutrients, less exposure to environmental stress factors and better translocation of bacterial metabolites throughout the host plant (Hallmann et al., 1997). Applicattion of endophyte Pseudomonas aeruginosa and B. cepacia could reduce BSR incidence up to 76% in 8 months oil palm seedling after disease inoculation (Sapak et al., 2008).
Members of the genus Burkholderia sp. are known for their ability to suppress soil-borne fungal pathogens by the production of various antibiotic compounds such as pyrrolnitrin and phenazines (Kirner et al., 1998). Other different antibiotics such 2,4-diacetylphloroglucinol (2,4-DAPG) and pyoluteorin has also been found responsible for suppression of soil-borne fungal pathogens (Subagio and Foster, 2003).
Antibiotic-related genes can be detected from BCAs by using Polymerase Chain Reaction (PCR) by using antibiotic specific primers which encode phenazine-1-carboxylic acid, 2,4-DAPG, pyoluteorin and pyrrolnitrin, from Pseudomonas and Bacillus genome (Zhang et al., 2005).
BCAs have been applied in many ways and on many crops species, for instance, seed dipping application on rice seeds to control bacterial blight disease caused by Xanthomonas oryzae was able to reduce the disease incidence (Suryadi et al., 2012). Besides microbial pathogen, BCA was also reported able to control plant parasitic nematodes. Munif et al. (2013) was reported seed dipping application on tomato seeds able to control Meloidogyne incognita penetration and enhanced the plant growth. Oil palm germinated seeds dipping application was also conducted by Dikin et al. (2003) to suppress Schizopyllum commune, causal agents of brown germ and seed rot in oil palm. In this study, Burkholderia sp. control activity against G. boninense growth were observed in vitro and in vivo of oil palm to analyze antifungal encoding gene from Burkholderia sp. potential.
MATERIALS AND METHODS
Burkholderia sp. isolation: Burkholderia sp. were isolated from soil near the roots and the roots that were taken from a healhty (symtompless) oil palm tree in an endemic area, in North Sumatera, Indonesia. One gram of soil was dilluted in sterlie water and poured on Nutrient Agar plates. For the roots, were surface sterilized with 90% alcohol for 1 min, 70% alcohol for 3 min and washed twice with 50 mL sterile distilled water for 30 sec. The roots were crushed aseptically and put on NA in plate. The plate were incubated for 24-36 h. Each of the grown colony were subculture and genome extracted for identification. Purified PCR product of 16S DNA were sent to 1st Base, Singapore.
Antagonist in vitro test: Antagonist test was observed to determine the Percentage Inhibition of Radial Growth (PIRG) of G. boninense (Bivi et al., 2010). Six bacterial isolates of Burkholderia sp. were selected to evaluate their efficacy in enhancing growth and inhibit the infection of BSR in oil palm in pre nursery. These Burkholderia sp. collection were isolated from rhizospher and oil palm root tissue. There were five rhizosphere Burkholderia sp. (B313, B51a, B52c, B51b, B52a) and one endophyte Burkholderia sp. (B212).
Burkholderia sp. was streaked into the PDA plate 2.5 cm from the edge of the petri dish. Agar disc cut diameter 5 mm and 5 day old G. boninense was placed 2.5 cm from the edge at the opposite side of the same petri dish. For the control plate, only G. boninense was placed in a similar manner without bacteria on a fresh petri dish. The plates were incubated at 28°C for five days. Results shown by measured the radial growth of G. boninense. The PIRG was calculated using the equation below (Zaiton et al., 2006):
Where:
| PIRG | = | Percentage inhibition of radial growth |
| R1 | = | Radial growth of G. boninense in the absence of bacteria (control) |
| R2 | = | Radial growth of G. boninense in the presence of Burkholderia sp. The three highest PIRG isolates from in vitro test were used for in vivo test |
In-vivo test: Burkholderia sp. suspensions were prepared by inoculating 24 h old cultures into Nutrient Broth (NB) and incubated for 20 h and adjusted to 108 CFU mL-1. During the preparation of mixture, equal volume of the 3 highest PIRG Burkholderia sp. were mixed. Ganoderma boninense inoculum was prepared on 6x3x15 cm sterilized oil palm fronds. The fronds were sterlized in a heat resistant plastic each before inoculated with G. boninense. After sterilization, the fronds were inoculated with a 0.5 cm diameter disc of G. boninense mycellia on agar plate. The plastic was sealed and incubated in room temperature for 3 months before used.
The plant material was selected as the most G. boninense susceptible progeny. Two different progenies were used in this study. Briefly, oil palm germinated seeds were treated with bacterial suspension (108 CFU mL-1), dipped for 20 min (seed bacterization) and air dried for 10 min before planted.
Oil palm germinated seeds were planted in polybags (15x20 cm) regarding to the Standard Operation Prosedure (SOP) in prenursery of oil palm plantation. There were four treatments in this study. Treatment A and C were not inoculated with G. boninense and treatments C and D were inoculated with G. boninense. Treatment B and C were using seedling treated with Burkholderia sp. Treatments with G. boninense, the seedlings were placed in contact with radicula. The pots were placed under shead, watered daily and no supplementary organic fertilizer was applied for 3 months (prenursery). A destructive observation was conducted after 3 months.
The infection of G. boninense on plant can be scored by observation on signs and symptom on the treatment plants using Disease Severity Index (DSI) (Abdullah et al., 2003). The DSI was observed from the external symtomp from foliar and the roots (destructive method).
The score can be calculated by the equation of Mohd Zainudin and Abdullah (2008):
Where:
| A | = | Disease class (0, 1, 2, 3 or 4) |
| B | = | Number of plants showing that disease class per treatment |
| Table 1: | Polymerase chain reaction primers and expected amplification products from genes encoding enzymes involved in the biosynthesis of several antibiotics (Zhang et al., 2005) |
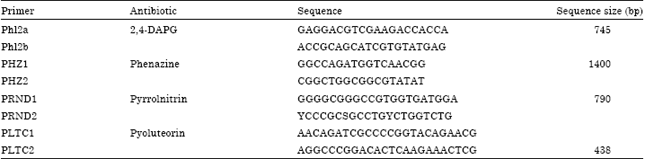 | |
Detection of antifungal gene: Antifungal gene detection was conducted by using PCR. Burkholderia sp. was prepared in liquid medium for DNA isolation by using GeneJET Genome DNA Purification Kit from Thermo Sciencetific. Specific primers for DAPG (Phl2a-Phl2b), phenazine (PHZ1-PHZ2), pyrrolnitrin (PRND1-PRND2) and pyoluteorin (PLTC1-PLTC2) were used for detection according to Zhang et al. (2005) (Table 1).
PCR product was gel extracted from agarose by using Gene JET Gel Extraction Kit (Fermentas), according to manufacturer’s instruction. After purification, the PCR products were sent to 1st Base, Singapore, to be sequenced. The antifungal genes sequences were aligned by using BioEdit software and searched for sequence similarity to other sequences which are available in the NCBI database at http://www.ncbi.nlm.nih.gov/ using Basic Local Alignment Search Tool (BLAST) algorithm. Multiple sequence alignments were performed on the selected closely related sequence accessions available using CLUSTAL W software in Mega 5.
Statistical analysis: The treatment was repeated into 13 replicates. This research used factorial design with one factor and the environmental design is complete randomized design. Statistic analysis was done by using general method linear model univariate. If there was a significant diferrence further analysis will be analyzed with á value is 5% by using SAS.
RESULTS
Antagonist in vitro test: All of the Burkholderia sp. isolates showed an inhibition activity against G. boninense growth in vitro (Table 2). Scores followed by the same letter indicated that they were insignificantly different scores. The highest inhibition activity was shown by isolate B212 with percentage PIRG was 34.38% but not significantly different with B52a and B52c which were both 27.50%. Burkholderia B313 showed the lowest activity (23.75%) and significantly different with B212 but still has an antagonist activity against G. boninense growth in vitro.
In vivo test: The necrosis and chlorosis foliar was not seen in all treatments, though the height of the plants between treatment was seem different (Fig. 1a). The DSI after 3 months only showed in treatment C and D (25%) (Table 3). Destructive observation showed that all plants in treatment C and D were infected with G. boninense. Treatment C and D which were inoculated with G. boninense, showed a brown-blackening roots especially on the parts which colonized with the G. boninense (Fig. 1b). The roots in treatment A and C, without G. boninense, were cream-brown colored.
 | |
| Fig. 1(a-b): | (a) Plants treatment left to right (A-D) and (b) Healthy root with no appearance of fungal mycellia (left). Appearance of fungal mycellia (red arrow) |
| Table 2: | Percentage of inhibition ratio from Burkholderia sp. against Ganoderma boninense growth in vitro |
 | |
| Table 3: | Disease Severity Index (DSI) of treated plant with Ganoderma boninense |
 | |
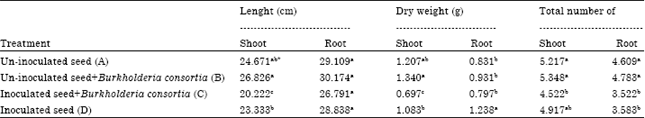
Progeny give a significant different mostly to the shoot lenght and dry weight (Table 4). Total number of roots between progeny also showed a significant difference. However, there is no correlation between progenies and treatments though in some parameter that were observed was show a significant difference.
Burkholderia sp. consortia application on inoculated seeds showed significantly lowest in shoot growth compare to control and other treatments. In all parameters that were measured, the treatment decrease the plant growth and significantly decrease shoot lenght, shoot dry weight, total number of shoot and root (Table 5). However, Burkholderia sp. consortia application on un-inoculated seeds showed the highest in shoot and root growth compare other treatments. The shoot lenght and dry weight was slightly different compare to control. The root lenght did not showed a significant different among all the treatment.
Antifungal gene detection: Primers PRND1 and PRND2 amplified the predicted 790 bp fragment from DNA of B212, B313, B51a and B52c. Primers PLTC1 and PLTC2 amplified the predicted 438 bp from DNA of Burkholderia B313, B51a and B52c (Fig. 2). Primers for phenazine and DAPG did not yield a PCR product from all isolates. This may be concluded that these isolates do not contain phenazine and DAPG biosynthesis gene.
| Table 4: | Effect of different progeny on root and leaf total number, lenght and dry weight after treatment with Ganoderma boninense and Burkholderia sp. |
 | |
| *Means within a column with the same letter are not significantly different at p<0.05 using DMRT | |
| Table 5: | Effect of dipping treatment of antagonistic bacteria on seedling of oil palm inoculated with Ganoderma boninense in prenursery at 3 months after planting |
 | |
| *Means within a column with the same letter are not significantly different at p<0.05 using DMRT | |
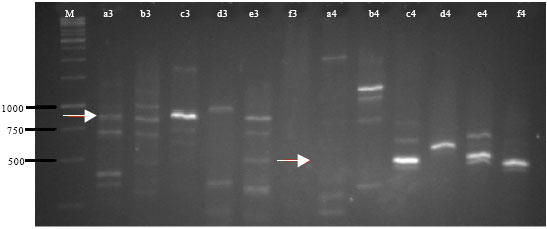 | |
| Fig. 2: | PCR product of pyrrolnitrin and pyoluteorin gene amplification. M: Marker, 3. prn primer, 4. plt primer, a: B212, b: B313, c: B51a , d: B52c, e: B51b, f: B52a. Red arrows showed the expected band size |
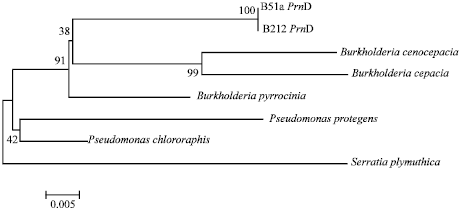 | |
| Fig. 3: | Phylogenetic tree of PrnD gene from B212 and B51a |
The PCR product were BLAST and showed a 99% identity with B. cepacia partial prnD gene, strain ESR63 (Fig. 3). It is confirmed B212 has a potency to produce pyrolnitrin and assumed pyrolnitrin is the responsible compound in inhibiting Ganoderma growth in vitro.
DISCUSSION
Ganoderma boninense has been the most threatening disease in oil palm plantation. Ganoderma boninense has the ability to degrade lignin in the plant and decay the lower stem and sometimes the root system, leading to severe symptoms such as flattening of the crown and unopened spear leaves (Cooper et al., 2011). Many attemps have been made to control this disease including using BCAs such as bacteria.
Bacteria have various mechanism of antagonistic such as synthesizing antibiotic compounds, production of hydrolytic enzymes, siderophore production, competition for substrates and also induction of systemic resistance in the host plant will increase the plant resistance to a broad spectrum of pathogens (Kloepper and Ryu, 2006). Many soil-borne plant diseases caused by fungi and oomycetes can be controlled by strains of the genus Burkholderia (Kirner et al., 1998).
In this study endophyte Burkholderia B212 showed the highest antagonist activity against G. boninense growth (Table 2). Several strains of Pseudomonas and Burkholderia species can produced a broad-spectrum of antibiotics which play an important role in the suppression of multiple plant pathogenic fungi (De Souza and Raaijmakers, 2003).
In vivo test showed a different result from the in vitro test. In comparison to Mohd Zainudin and Abdullah (2008) and Sapak et al. (2008), the external symptom of G. boninense infection in oil palm appeared after 4 months after planting even though the disease has been invested from seedling or after 3 months planting. However, external symptom may also occur after 2 months planting with the disease.
This diferences may be caused by many factors such as pathogenicity level of G. boninense, inoculum source of G. boninense, plant tolerance and climate. White-rot fungi has been classified to the ligninolytic enzymes they express and able developed unspecific ligninolytic systems consisting of peroxidases and laccases which employ an oxidative process to degrade the wood cells (Paterson, 2007). It can be assumed that these enzymes activity can be related to G. boninense pathogenicity level. There are two type of G. boninense inoculum source used in G. boninense research, wood block and oil palm fronds. Oil palm fronds are more easy to get since it is abundance in oil palm plantation. Different climate may also effect lignin degradation which lignin degradation is less efficient at 37°C compare to 25°C (Paterson et al., 2008).
Application method of Burkholderia sp. may also effect the result of infection. Burkholderia sp. has been reported that its antifungal producing is related to quorum-sensing which is mean that the antifungal activity depends on Burkholderia sp. biomassa (Chapalain et al., 2013). Ganoderma boninense infection were found in all plants which were treated with the disease including the Burkholderia sp. consortia treatment. Burkholderia sp. consortia were expected to inhibit the disease infection but the result showed on the contrary. It is very different to other report which is using soil drenched application and the result showed that application with Burkholderia and Pseudomonas able to reduce G. boninense incidence (Sapak et al., 2008). Different method of application and number of Burkholderia sp. applied may come to a different results.
Burkholderia sp. consortia application on inoculated seeds was expected to have the highest plant growth compare to plants without Burkholderia sp. and control. However, the treatment showed the lowest plant growth compare to all other treatments. Burkholderia sp. as endophyte has the ability to penetrate the plant by using cellulase enzyme (Reinhold-Hurek et al., 2006). It can be assumed that the Burkholderia sp. has a potency to open a way for G. boninense to infect the plants. Biomass of Burkholderia sp. held an important key in suppress G. boninense infection, since, the antifungal compound is regulated by quorum sensing. This fenomena could explain the infected plants without Burkholderia sp. consortia application is higher than the infected plants with Burkholderia sp. consortia.
If compared with the shoot and root ratio (S/R) among the treatment, infected plants with Burkholderia sp. consortia shown the lowest number. Which means that in this treatment, the root has the higher biomass compare to the shoot. Burkholderia sp. consortia application gave a more affect to the roots than the shoots on infected plants. Burkholderia sp. also could acted as PGPR which can lead an indirect biocontrol agent, since it could enhanced plant growth. Burkholderia has reported as a plant growth promoting rhizobacteria (PGPR) (Compant et al., 2008).
Burkholderia sp. consortia application on inoculated seed was significantly higher on root dry weight compare to other treatment. Ganoderma boninense as pathogen could induced plant defense by forming lignin as its first defence system. Lignification or cell thickness is the form of plant defense against pathogen (Xu et al., 2011) and could increase root biomass.
Total number of Burkholderia applied also play an important role in proper activity of this biocontrol agent. Schmidt et al. (2009) has reported that some of antifungal such as pyrrolnitrin is regulated by quorum sensing. Quorum sensing is a mechanism to regulate the production of antimicrobial compounds by population-density-dependent (Liu et al., 2007). Different results between in vitro and in vivo test may be caused by many factors such as plant ages by the time artificially inoculated, concentration of microbes applied and application technique.
Many research showed that Burkholderia could decrease G. boninense infection on oil palm but yet a correct application technique of this bacteria also important to get the best result. Antifungal compound is a secondary metabolite which is produce by bacteria in their stationary phase. Further research need to be conducted for the proper technique application of this Burkholderia for its optimum action.
Pyrrolnitrin encoded gene was amplified and showed that Burkholderia B212 has a potential in producing antifungal agent such as pyrrolnitrin. Pyrrolnitrin has been implicated as an important mechanism of biological control of fungal plant pathogens by several Pseudomonas strains (Hasan and Turner, 1998). Pyrollnitrin is a chlorinated phenylpyrrole antibiotic that was first isolated from Burkholderia pyrrocinia (Kloepper and Ryu, 2006) and later from other microorganisms, including Pseudomonas fluorescens, P. chlororaphis, P. aureofaciens, B. cepacia, Enterobacter agglomerans, Myxococcus fulvus and Serratia species (Hammer et al., 1999).
Pyrrolnitrin is synthesize by four protein encoded by 4 gene, prna, prnb, prnc and prnd. The Prnd gene was the final protein to form an active pyrrolnitrin compound. The prnD catalyzes the oxidation of the amino group of aminopyrrolnitrin to a nitro group to form pyrrolnitrin (Kirner et al., 1998). In some strains of Burkholderia species pyrrolnitrin biosynthesis was shown to be regulated by quorum sensing (Schmidt et al., 2009).
ACKNOWLEDGMENT
The authors would like to thank the valuable technical assistance of staff from the PT. SMART Tbk. in Microbiome Technology laboratory and Biotechnology laboratory. The authors also thank to PT. SMART Tbk for the research funding.
REFERENCES
- Bivi, M.R., M.S.N. Farhana, A. Khairulmazmi and A. Idris, 2010. Control of Ganoderma boninense: A causal agent of basal stem rot disease in oil palm with endophyte bacteria in vitro. Int. J. Agric. Biol., 12: 833-839.
Direct Link - Chapalain, A., L. Vial, N. Laprade, V. Dekimpe, J. Perreault and E. Deziel, 2013. Identification of quorum sensing-controlled genes in Burkholderia ambifaria. MicrobiologyOpen, 2: 226-242.
CrossRef - Compant, S., J. Nowak, T. Coenye, C. Clement and E. Ait Barka, 2008. Diversity and occurrence of Burkholderia spp. in the natural environment. FEMS Microbiol. Rev., 32: 607-626.
CrossRef - Cooper, R.M., J. Flood and R.W. Rees, 2011. Ganoderma boninense in oil palm plantations: Current thinking on epidemiology, resistance and pathology. Planter, 87: 515-526.
Direct Link - De Souza, J.T. and J.M. Raaijmakers, 2003. Polymorphisms within the prnD and pltC genes from pyrrolnitrin and pyoluteorin-producing Pseudomonas and Burkholderia spp. FEMS Microbiol. Ecol., 43: 21-34.
CrossRefDirect Link - Dikin, A., K. Sijam, M.A. Zainal Abidin and I.A. Seman, 2003. Biological control of seedborne pathogen of oil palm, Schizopyllum commune Fr. with antagonistic bacteria. Int. J. Agric. Biol., 5: 507-512.
Direct Link - Haas, D. and G. Defago, 2005. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol., 3: 307-319.
CrossRefPubMedDirect Link - Hallmann, J., A. Quadt-Hallmann, W.F. Mahaffee and J.W. Kloepper, 1997. Bacterial endophytes in agricultural crops. Can. J. Microbiol., 43: 895-914.
CrossRefDirect Link - Hammer, P.E., W. Burd, D.S. Hill, J.M. Ligon and K.H. Pee, 1999. Conservation of the pyrrolnitrin biosynthetic gene cluster among six pyrrolnitrin-producing strains. FEMS Microbiol. Lett., 180: 39-44.
CrossRef - Hasan, Y. and P.D. Turner, 1998. The comparative importance of different oil palm tissues as infection sources for basal stem rot in replantings. Planter, 74: 119-135.
Direct Link - Kirner, S., P.E. Hammer, D.S. Hill, A. Altmann and I. Fischer et al., 1998. Functions encoded by pyrrolnitrin biosynthetic genes from Pseudomonas fluorescens. J. Bacteriol., 180: 1939-1943.
Direct Link - Liu, X., M. Bimerew, Y. Ma, H. Muller and M. Ovadis et al., 2007. Quorum-sensing signaling is required for production of the antibiotic pyrrolnitrin in a rhizospheric biocontrol strain of Serratia plymuthica. FEMS Microbiol. Lett., 270: 299-305.
CrossRef - Munif, A., J. Hallman and R.A. Sikora, 2013. The influence of endophytic bacteria on Meloidogyne incognita infection and tomato plant growth. J. ISSAAS, 19: 68-74.
Direct Link - Paterson, R.R.M., 2007. Ganoderma disease of oil palm-A white rot perspective necessary for integrated control. Crop Protect., 26: 1369-1376.
CrossRefDirect Link - Paterson, R.R.M., S. Meon, A.M.A. Zainal and N. Lima, 2008. Prospects for Inhibition of lignin degrading enzymes to control Ganoderma white rot of oil palm. Curr. Enzym. Inhib., 4: 172-179.
Direct Link - Reinhold-Hurek, B., T. Maes, S. Gemmer, M. Van Montagu and T. Hurek, 2006. An endoglucanase is involved in infection of rice roots by the not-cellulose-metabolizing endophyte Azoarcus sp. strain BH72. Mol. Plant-Microbe Interact., 19: 181-188.
CrossRef - Schmidt, S., J.F. Blom, J. Pernthaler, G. Berg, A. Baldwin, E. Mahenthiralingam and L. Eberl, 2009. Production of the antifungal compound pyrrolnitrin is quorum sensing‐regulated in members of the Burkholderia cepacia complex. Environ. Microbiol., 11: 1422-1437.
CrossRefDirect Link - Xu, L., L. Zhu, L. Tu, L. Liu and D. Yuan et al., 2011. Lignin metabolism has a central role in the resistance of cotton to the wilt fungus Verticillium dahlia as revealed by RNA-Seq-dependent transcriptional analysis and histochemistry. J. Exp. Bot., 62: 5607-5621.
CrossRef - Zaiton, S., M. Sariah and M.A. Zainal Abidin, 2006. Isolation and characterization of microbial endophytes from oil palm roots: Implication as biological control agents against Ganoderma. Planter, 82: 587-597.
Direct Link - Sapak, Z., S. Meon and Z.A.M. Ahmad, 2008. Effect of endophytic bacteria on growth and suppression of Ganoderma infection in oil palm. Int. J. Agric. Biol., 10: 127-132.
Direct Link - Zhang, Y., W.G. Fernando, T.R. de Kievit, C. Berry, F. Daayf and T.C. Paulitz, 2005. Detection of antibiotic-related genes from bacterial biocontrol agents with polymerase chain reaction. Can. J. Microbiol., 52: 476-481.
PubMed