
D. Ruswandi
Faculty of Agriculture, Padjadjaran University, Bandung, 40600, Indonesia
Agustian
Faculty of Agriculture, Padjadjaran University, Bandung, 40600, Indonesia
E.P. Anggia
Faculty of Agriculture, Padjadjaran University, Bandung, 40600, Indonesia
A.O. Canama
College of Agriculture, University of Philippines, Los Banos, 4031, Philippines
H. Marta
Faculty of Agriculture Engineering, Padjadjaran University, Bandung, 40600, Indonesia
S. Ruswandi
Faculty of Agriculture, Padjadjaran University, Bandung, 40600, Indonesia
E. Suryadi
Faculty of Agriculture, Padjadjaran University, Bandung, 40600, Indonesia
Asian Journal of Agricultural Research
Year: 2014 | Volume: 8 | Issue: 5 | Page No.: 234-247







ABSTRACT
Drought stress is a very important factor in the reduction of maize production in Indonesia. A series of experiments were conducted to identify early maturing mutants, to study their genetic similarity and to select for mutants tolerant to drought stress. The first experiment was done in Jatinangor to identify early maturing second generation mutants (M2). The experiment was laid on augmented design with two replications in which fifteen M2 population groups with their nonmutant parentals were used as genetic materials. The second experiment was conducted in Biotechnology Laboratory, Institute of Plant Breeding, University of the Philippines at Los Banos to analyse DNA of 28 early maturing M2 using SSRs markers. The third experiment was conducted in Majalengka to select mutants tolerant to drought stress. The experiment was arranged in augmented design consisting of two replications of 18 nonmutant parental lines and 161 M3 which have been selected for early maturity. The last experiment was laboratory screening in the Plant Breeding Laboratory, University of Padjadjaran to confirm field drought tolerant mutants. The experiment was arranged in a Randomize Block Design (RBD) using 64 of M4 which showed early maturity. In the first experiment, there were 35 M2 that showed early maturity. Gamma irradiation was found to increase the phenotypic variation and diversity of plant height, ear weight, days to tasseling and days to harvesting. The SSR analysis was found to be a valuable DNA marker system to study genetic diversity of mutant and non-mutant lines. The mutant and non-mutant lines were clustered into two major cluster and seven sub-clusters based on a phylogenetic tree analysis using UPGMA. Based on the field screening for drought in the third experiment and confirmatory experiment in the laboratory, M3DR 18.8 and its progeny of M4DR 18.8.1 (selfing of 18.8) and M3DR 18.5 and its progeny, M4DR 18.5.1 (selfing of 18.5) are tolerant mutant lines as shown by their positive index of drought and their physiological response to water stress simulation using PEG. Those mutant lines could be considered for breeding program for tolerance to drought as an anticipation of global climate change.
PDF Abstract XML References Citation
Received: May 15, 2014;
Accepted: September 13, 2014;
Published: October 28, 2014
How to cite this article
D. Ruswandi, Agustian, E.P. Anggia, A.O. Canama, H. Marta, S. Ruswandi and E. Suryadi, 2014. Mutation Breeding of Maize for Anticipating Global Climate Change in Indonesia. Asian Journal of Agricultural Research, 8: 234-247.
DOI: 10.3923/ajar.2014.234.247
URL: https://scialert.net/abstract/?doi=ajar.2014.234.247
DOI: 10.3923/ajar.2014.234.247
URL: https://scialert.net/abstract/?doi=ajar.2014.234.247
INTRODUCTION
Maize is an important global staple food. Tropical countries such as Indonesia is an important determinant of the world maize grain supply since it provides over 50% of the maize production in South East Asia. However, there are 56 million hectares of drought-prone fields in Indonesia affected by global climate change.
Soil and groundwater are key environments in maize production since they are depleted through erosion, salinization, compaction, net nutrient export and diminishing water supply (Cassman, 1999). Thus, global climate change is now considered to be happening (Hillel and Rosenzweig, 2002) and is expected to cause inclined temperatures and evapotranspiration and eventually an increased incidence of drought. These trends, doubled with an extention of cropping into marginal production areas, are generating progressively more drought-prone maize production environments (Campos et al., 2004).
Maize is very sensitive to drought during reproductive stage i.e., flowering, silk growth, pollination and kernel set (Shaw, 1977). Water stress slows ear growth and silk emergence, more than tassel growth or anthesis, resulting in a widening interval between anthesis and silking (ASI). Under stress condition during flowering, yield of tropical maize shows a strong dependency on kernel number per plant, barrenness and ASI (Bolanos and Edmeades, 1996). Silk emergence is an indirect measure of ear growth rate and since tassel growth is little affected by drought stress, ASI becomes a reporter trait for ear and plant growth rates during the flowering period (Edmeades et al., 2000; Vega et al., 2001). In order to respond to water stress, the tolerant maize roll their leaf, close the stomata and thicken the cuticula to maintain plant turgor (Ludlow and Muchow, 1990). Based on these mechanism, maize tolerant to drought can be identified using the following traits: Grain yield, ears per plant, ASI, leaf senescence, tassel size and leaf rolling (Banziger et al., 2000).
In plant breeding, mutation is a very valuable tool for improving one or two easily identifiable traits in a well-adapted variety. By mutation, the basic genotype of the variety is usually only slightly altered while the improved trait’s is added. The time required to breed the improved variety can also be shorter than when hybridization is used to achieve the same result (Sigurbjornsson, 1977). Among the plant traits improved by mutation breeding include: Yield, flowering and ripening time, adaptability, plant type and growth habit, resistance to lodging and stem breakage, shattering and shedding resistance, tolerance to temperature, drought, heat and salinity, disease and pest resistance and quality (Aatsveit et al., 1977). Some successfull samples include: Chlorina-a tobacco cultivar, Pallas and Mari for barley cultivar and Calrose 76 of rice cultivar (Acquaah, 2007).
The detection of mutant lines can be based on their agro-morphologycal traits, biochemical content (Gallego and Martinez, 1996) and molecular markers. Molecular markers provide an effective tool for detection of desired agronomic traits since they are based on plant genotypes and thus are independent of environmental variation (He et al., 2003). Markers can help the breeders to identify the presence of desirable alleles in individual plants. Breeders do need costly and time consuming phenotypic evaluation to determine whether or not the alleles are present. Due to reasons of cost and simplicity, reliability, reproducibility, discrimination and standardization, researchers prefer Simple Sequence Repeat (SSRs) or microsatellite markers. Ruswandi et al. (2014) for example used molecular markers linked to Quantitative Trait Loci (QTL) associated with downy mildew resistance traits to select for resistance to the disease even in the absence of the pathogen.
SSRs are short, repeated DNA sequences which are co-dominant, highly reproducible and are multi-allelic variations distributed in plant genomes (Dreher et al., 2000). Since, the number of SSRs present at a particular location in the genome tends to differ among plants, SSRs can be analyzed to determine the absence or presence of specific alleles. The SSR markers are preferred molecular markers in crop breeding and are most practical markers for genomic mapping, variety identification, marker-assisted selection (He et al., 2003). It is also used for studying genetic diversity as well as genetic relationship in other crop plant species (Goldstein et al., 1995; Perera et al., 2000; Zhang et al., 2002; He et al., 2003).
The objectives of the study are: (1) To select second generation mutants (M2) based on their phenotypic performance and early maturing traits, (2) To identify the selected mutants using SSR markers and (3) To select mutants tolerant to drought stress and to confirm field drought tolerant mutants.
MATERIALS AND METHODS
Selection of early maturing of second generation mutants (M2): Genetic materials to used were fifteen M2 population groups composed of 523 M2, with their non-mutant parentals i.e., DR1, DR2, DR3, DR4, DR5, DR6, DR7, DR8, DR9, DR10, DR 12, DR 14, DR 16, DR 18 and DR 20.
Experimental design: The first experiment was carried out from September 2010 upto January 2011 in Jatinangor, West Java, Indonesia to select for early maturing second generation mutants (M2). The experiment was laid on augmented design with two replications in which fifteen M2 population groups composed of 523 M2, with their nonmutant parentals used as check varieties. The traits observed were days to silking (days after planting-dap), days to tasseling (dap), days to harvest (dap), plant height (cm), plant diameter (cm), leaf numbers and ear weight (g). Some statistic parameters were estimated to evaluate the M2 mutant population, such as: mean, standard deviation, minimum and maximum value. The difference between new mutant M2 lines and their nonmutant parental was estimated using Least Significant Increase (LSI) following Petersen (1994).
Molecular identification of selected early maturing mutants: Twenty eight 2nd generation selected mutants (M2) for early maturity and their non-mutant parental lines were germinated in a screen house. Fifteen seeds of every accession was planted in the soil box containing of fertile soil. The leaves were then prepared for DNA extraction 15 days after planting.
Experimental design: The second experiment was conducted in Molecular Biology Laboratory of Institute of Plant Breeding, University of The Philippines at Los Banos from November 2010 upto December 2010. The protocol for fingerprinting using SSR markers included: DNA extraction, DNA amplification, electophoresis and visualisation following AMBIONET (Asian Maize Biotechnology Network), Philippines, CIMMYT ABC (Agricultural Biotechnology Center) (George et al., 2004). DNA was amplified using eight optimized primers, namely: Umc 1149, Bngl 137, Phi 112, Phi 113, Phi 36, Phi 126, Bngl 589 and Bngl 125.
The bivariate 1-0 data were used to estimate genetic similarity on the basis of the number of shared amplification product. The genetic similarity was estimated based on formulae described by Rohlf (2001):
where, m is the number of similar alleles, n is the total number of allele and u is the number of non similar allele.
Genetic relationship among inbred lines were evaluated with phylogenetic trees constructed by the Unweighted Pair Group Method Average (UPGMA) using NTSYS-pc software version 2.1 (Rohlf, 2001). Bootstrap analysis using winboot program was also conducted to estimate the level of significance of the phylogenetic trees.
Selection of drought tolerant mutants: Genetic materials used were 161 third generation mutants (M3) including 28 early maturing selected mutants plus others superior mutants which have been selected for yield components in second generation.
Experimental design: The third experiment was conducted in Majalengka, West Java, Indonesia during dry season from July 2011 upto October 2011 to select mutants tolerant to drought stress. The experiment was arranged in augmented design consisting of two replications of 18 nonmutant parental lines as varietal check and 161 third generation mutants (M3). The variable of drought observed in the second experiment were leaf rolling, leaf senescence, ASI (Anthesis Silking Interval) and ear per plant, whereas in the confirmatory experiment in the laboratory, the variable observed were height, root length, chlorophyll content, wet weight and dry weight of seedling (Banziger et al., 2000). Tolerance to drought was analyzed based on estimating the drought index (Lin, 1978) as:
I = b1P1+b2P2+b3P3+b4P4+b5P5+b6P6…+bnPn
Pi = (xij-mi)si
Where:
| Pi | = | Phenotypic value of the-ith traits |
| xij | = | Average of the-ith traits |
| mi | = | Mean of average of the-ith traits |
| si | = | Standard deviation of the-ith traits |
P1, P2, P3, P4, P5 and P6, respectively were phenotypic value of grain weight ha-1, number of ear per plant, ASI, leaf senescence, leaf rolling, b1, b2, b3 and b4, respectively, b4 is the weight of number of ear per plant = 3, b3 is the weight of ASI = 2 and the weight of leaf senescence = 2; and b1 is the weight of leaf rolling = 1 (Wahyudi et al., 2006).
Confirmatory of drought tolerant mutants: Sixty four of 4th generation mutants (M4) which showed early maturity (88-93 days after planting) and field tolerance to drought were used as genetic materials.
Experimental design: The last experiment was aimed to confirm field mutants tolerant to drought. The test was conducted in the Plant Breeding Laboratory, University of Padjadjaran. Sixty four of fourth generation mutants (M4) were planted on nutrient solution containing PEG 6000. The plant was arranged in a Randomize Block Design (RBD) which was replicated twice. Scot Knot multiple range test was used to differentiate the significance of the mutant lines. The media for planting contained of 4 mM CaCl2.2H2O, 6.5 mM KNO3, 0.4 mM NH4NO3, 2.5 mM MgCl2.6H2O and 0.1 mM (NH)2SO4) (Anas and Yoshida, 2000) and 15% of PEG 6000 (Wang et al., 2008). The sterile M4 maize seeds were germinated in the petridish and incubated in the culture room. The germinated seeds were then planted on the planting trays (42x32 cm) containing nutrient solution with PEG 6000. The trays were covered with plastic foam which have holes 4x4 cm for holding the maize seedlings. The traits observed included normal and abnormal seedling, seedling height (cm), seedling root (cm), chlorophyl content (CCI), fresh weight of seedling (g) and dry weight of seedling (g).
RESULTS AND DISCUSSION
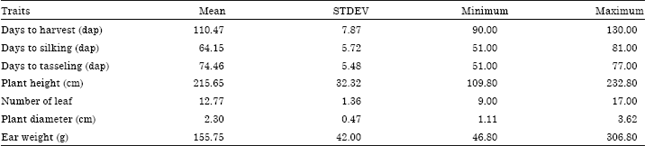
Selection of early maturing of second generation mutants (M2): Statistical parameters, such as mean, standard deviation, minimum and maximum values for reproductive and agronomic traits of second generation of mutant population are presented in Table 1. Reproductive traits which are related to early maturity traits showed broad ranges from 90-130 dap, 51-81 dap and 51-77 dap, for days to harvesting, days to silking and days to tasseling, respectively. Agronomic traits which are related to good performance also had broad ranges which are from 109.80-232.80 cm, 9-17 and 46.80-306.80 g, for plant height, number of leaf and ear weight, respectively. Based on these statistics, it can be concluded that the mutant populations are varied for every trait, including reproductive and agronomic traits. Thus, gamma ray irradiation could change the gene structure of non-mutant parental lines which eventually increase the genetic variation of the early generation of the mutants.
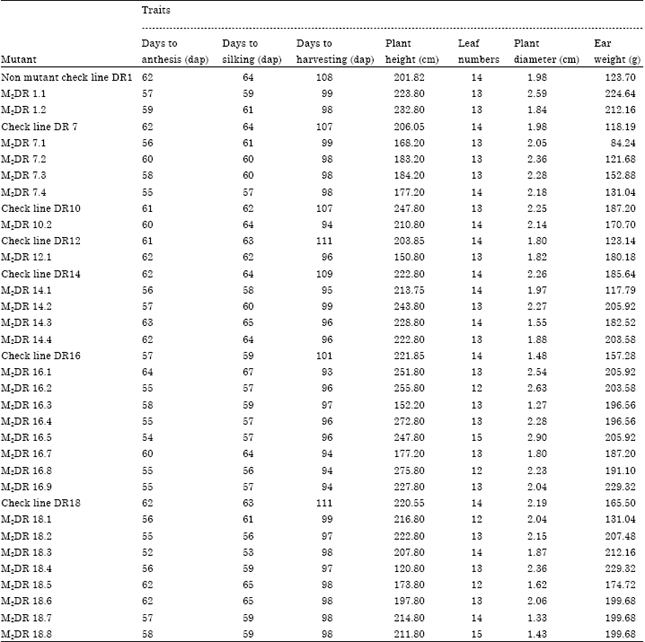
The second generation of mutant population which showed early maturity traits and high yield components as measured by indicator traits i.e., days to harvesting, days to tasseling, days to silking, together with ear weight were selected. Of 523 second generation mutants (M2), 28 mutants were selected based on their early maturity as well as agro-morphologic traits. Those selected second generation mutants are presented in Table 2. Out of 28 second generation mutant lines, 11 second generation mutants showed earlier (52-56 dap) days to anthesis than the shortest check line DR 16 (57 dap), 9 second generation mutants showed earlier (53-58 dap) days to silking than the shortest check line DR 16 (59 dap). Surprisingly, all selected second generation mutants showed earlier days to harvesting than the shortest check line DR 16 (101 dap). The result proved that genetic variation for maturity in maize population improved due to mutation. This result was similar to that of Herison et al. (2008). They reported that the chance of mutation is higher on early generation at M1 or M2 in irradiation of open pollinated population. In this early generation, segregated loci of mutated structure of chromosome (s) or gene (s) or DNA had been fixed. Therefore, selection is possible to be applied.
| Table 1: | Mean, standard deviation, minimum and maximum statistics of second generation of mutants (M2) |
 | |
| Table 2: | Twenty eight selected second generation of early maturing mutants M2 with other agronomic performance |
 | |
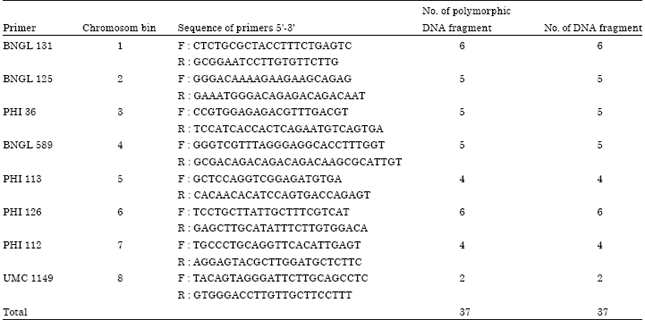
Molecular identification of selected early maturing mutants: The DNA amplification of second generation mutant population M2 DR using SSR primers is presented in Table 3. Results show that the DNA amplification of the mutant lines were polymorphic. According to Kartikaningrum et al. (2002), the number of polymorphic fragment of DNA for genetic diversity analysis is very important to determine the level of variation in the population. The total number of DNA fragment would portray the crop genome. It also reduces the bias due to the unrepresentation of particular fragments of the genome.
The pattern of DNA fragment from the mutant lines differed from their non-mutant parental lines as shown by DNA amplification of mutant and non-mutant parental lines using BNGL 125 in Fig. 1. For example, mutant line M2DR 1.1 and M2DR 1.2, on lane 2 and 3, respectively, were different from their non-mutant parental line of DR 1 on lane 1.
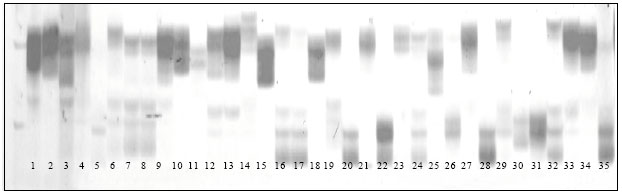 | |
| Fig. 1: | DNA amplification of 28 mutant and their non-mutant parental lines using primer BNGL 125, Notes: 1 = DR 1.C, 2=M2DR 1.1, 3 = M2DR 1.2, 4 = DR 7.C, 5 = M2DR 7.1, 6 = M2DR 7.2, 7 = M2DR 7.3, 8 = M2DR 7.4, 9 = DR 10.C, 10 = M2DR 10.2, 11 = DR 12.C, 12 = M2DR 12.1, 13 = DR 16.C, 14 = M2DR 16.1, 15 = M2DR 16.2, 16 = M2DR 16.3, 17 = M2DR 16.4, 18 = M2DR 16.5, 19 = M2DR 16.7, 20 = M2DR 16.8, 21 = M2DR 16.9, 22 = DR 14.C, 23 = M2DR 14.1, 24 = M2DR 14.2, 25 = M2DR 14.3, 26 = M2DR 14.4, 27 = DR 18.C, 28 = M2DR 18.1, 29 = M2DR 18.2, 30 = M2DR 18.3, 31 = M2DR 18.4, 32 = M2DR 18.5, 33 = M2DR 18.6, 34 = M2DR 18.7, 35 = M2DR 18.8 |
| Table 3: | Amplification of DNA using eight SSR primers |
 | |
The difference between them is due to mutation by gamma irradiation of the parental lines. The effect of gamma irradiation could randomly occur anywhere along the genome of the crop. Maize is a naturally open pollinated crop, therefore, variation of the mutant would be greater due to the crossing over of the mutated DNA fragment on the genome. As a result, second generation mutants have different DNA pattern as revealed by SSR markers eventhough they have similar ancestor and had been treated with the same dosage of gamma irradiation.
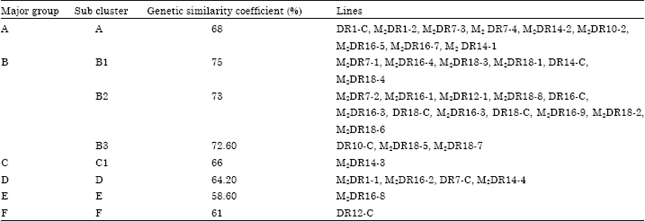
Multivariate analysis was conducted to generate a similarity matrix using Jaccard coefficient to estimate genetic diversity and relatedness among 35 maize inbred lines which is composed of 28 mutants and 7 non-mutant parental lines. Results show that the genetic relatedness among 35 maize lines was from 0.39 upto 0.95 (39% upto 95%). It can be said that genetic diversity was from 5% upto 61%. Co-phenetic coefficient (r) at 0.80 explained that the number of SSR markers were fit to make a phylogenetic tree (Rohlf, 2001). These coefficients were used to construct a phylogenetic tree/dendogram by UPGMA analysis to determine the grouping of the 28 second generation mutants and 7 non-mutant parental lines. These mutant and non-mutant lines were clustered into two major cluster and seven sub-clusters (Fig. 2 and Table 4). Results showed that second generation mutants M2DR 7-3 had close similarity with M2DR 7-2 as shown by diversity coefficient of 5% or similiraty coefficient of 95%. These mutants originated from gamma ray induction of non-mutant parental line DR7. The mutagen is effective in altering the DNA of the plant as proven in Fig. 2. Most of the mutants were grouped in different cluster compared to their non-mutant parental lines.
| Table 4: | Grouping of 28 mutant and seven non-mutant parental lines based on phylogenetic trees using SSR |
 | |
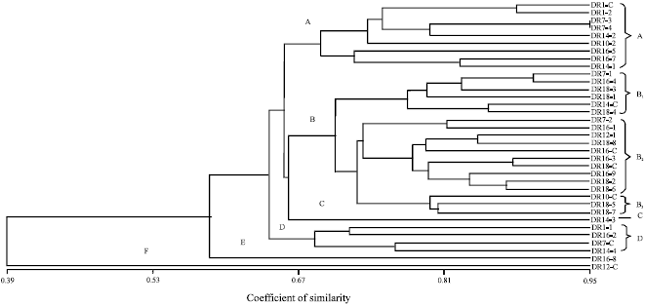 | |
| Fig. 2: | Genetic similarity of 28 second generation mutan M2 and their non-mutant parental lines using SSRs markers |
The most extreme case was shown by mutant of M2DR 16-2 that is located in a different major cluster from its non-mutant parental line of DR 16.
The phylogenetic tree on Fig. 2, delineated most of the non-mutant parental lines into groups which corresponds well with the breeding programs adopted at different institutes of release. The DR 16 and DR 18, high nutrient lines were grouped in the sub-cluster B1. These lines were mass selected by Plant Breeding Laboratory of University of Padjadjaran from CML 161 and CML 163 which have been developed by CIMMYT-Mexico as QPM lines (Ruswandi et al., 2005, 2014) whereas DR 12 and DR 14 are shown as downy mildew resistant lines, grouped into sub cluster A and F, respectively. These lines were mass selected by Plant Breeding Laboratory of University Padjadjaran from Ki 3 and P 345 which was developed by the National Institute of Agriculture Research Center of Thailand as downy mildew resistant lines (Ruswandi et al., 2005, 2014). Furthermore, DR 7 and DR 8 are non-mutant parental lines which were developed for QPM as well as downy mildew resistant lines. These lines were closely related since they are half-sib and have similar parent, i.e., Nei 9008 as female and CML 172 as male parental lines They are grouped closely in sub- cluster B3 and D. Similar to this result, clustering of genotypes according to their center of release and pedigree relationship have been reported in cotton and wheat (Iqbal et al., 1997; Mukhtar et al., 2002) and maize (Ruswandi et al., 2005).
In the present study, the SSR analysis was found to be a valuable DNA marker system to study genetic diversity of mutant and non-mutant lines. The information on genetic similarity will avoid the elite germplasm becoming genetically uniform. This can endanger the progress of long term productivity (Messmer et al., 1992). It is suggested that oriented breeding programs complementary with DNA fingerprinting can produce distinct genotype or cultivars that will maintain steady genetic improvement.
Selection of drought tolerant mutants: Results of the selection index of drought for ear weight per plant, ASI, leaf rolling and leaf senescence showed positive index of drought for 84 third generation mutant line M3. In contrast, 77 mutant line M3 showed negative index of drought. Since the mutant line with the negative index of drought showed more sensitive reaction to water stress, then only mutant lines with positive index of drought were selected as parental lines in the breeding program for drought tolerance in maize.
There were 10 third generation M3 mutant lines which were selected based on index of drought (Table 5). The M3DR 9.2 is the third generation of mutant with the highest positive index of drought at 25.49; whereas, M3DR 2.6 is the mutant with the lowest positive index of drought at-14.84. On the other hand, check variety of DR 8 showed the highest index of drought at 2.37 while DR 7 showed the lowest negative index of drought at -3.12. Based on the index of drought, drought tolerant hybrid of E6xB-11-157 at the index of drought at 7.0 which is 4.3% higher compared to the check variety tolerance of BISI-2; were selected in previous studies. Another third generation mutant lines considered as parental lines were M3DR 3.4, M3DR 8.5.2, M3DR 14.7, M3DR 18.5.1 and M3DR 18.8 with the indices of drought at 7.79, 8.85, 9.37, 6.99 and 8.23, respectively.
Confirmatory test of drought tolerant mutants: Results of the confirmatory test using nutrition with PEG-6000 showed the reduction of seedling length, chlorophyll content, root length, fresh and dry weight of seedling compared to mutants planted in control media. Scott-Knott multiple range test showed that M4 mutant lines with notation (a) have better performance than mutant lines with notation (b). The range of this group are 7.97-25.93 cm for seedling length; 5.83-15.78 for chlorophyll content, 7.23-19.62 cm for root length, 0.45-5.44 g for fresh seedling weight and 0.23-0.77 g for dry seedling weight, respectively.
Based on Scott-Knott multiple range test, the fourth generation of mutant lines M4DR 1.6.2, M4DR 3.6.1, M4DR 16.1.1, M4DR 18.1.1, M4DR 18.2.1, M4DR 18.5.1 and M4DR 18.8.1 are categorized as group (a) for all observed traits. It meant that those mutant lines performed better than the other mutant lines. In addition, M4DR 18.8.1 is the best mutant line. It showed the highest mean for all observed traits.
Seven of fourth generation mutant lines showed longer root length both in control and nutrition with PEG-6000. Those mutant lines are M4DR 1.6.2, M4DR 3.6.1, M4DR 16.1.1, M4DR 18.1.1, M4DR 18.2.1, M4DR 18.5.1 and M4DR 18.8.1. This indicated that mutant lines were more tolerant to drought than the rest of the mutant lines. Similar result was also mentioned by Effendi et al. (2010) who observed tolerance genotype performing longer root length in control media and PEG 6000 media than susceptible check variety to drought. In the mutant lines with longer root length, water stress had less effect than the mutant lines with short and less root length. Therefore, the long root mutant lines are more tolerant to drought than the short and less root ones. According to Wu and Cosgrove (2000), together with the seedling length and fresh and dry weight of seedling, the root growth is the key factor for the plant to be able to adapt in water stress. Effendi et al. (2010) explained that the drought tolerant plant incline the absorption of the water through increasing number of root and reducing the water loss through decreasing the plant canopy.
Water stress in plants reduce synthesis of chlorophyll on the leaves which consequently decrease the photosynthesis rate, increase plant temperature and increase transpiration. These cause the chlorophyll on the leaves to disintegrate. The reduction of chlorophyll content is an important physiological response of crop due to water stress. In drought tolerant plants, the response is less than on the susceptible ones.
| Table 5: | Ten best third generation maize mutant tolerant to drought based on drought index |
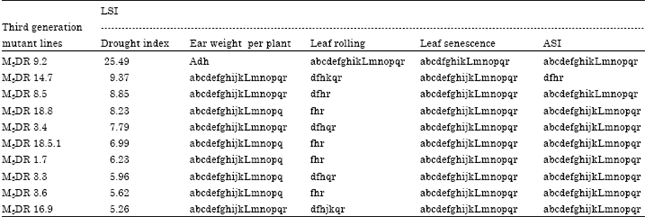 | |
a: Significantly different with non mutant line check BR153 based on LSI test at level of 5%, b: Significantly different with non mutant line check DR1 based on LSI test at level of 5%, c: Significantly different with non mutant line check DR 2 based on LSI test at level of 5%, d: Significantly different with non mutant line check DR 3 based on LSI test at level of 5%, e: Significantly different with non mutant line check DR 4 based on LSI test at level of 5%, f: Significantly different with non mutant line check DR 5 based on LSI test at level of 5%, g: Significantly different with non mutant line check DR 6 based on LSI test at level of 5%, h: Significantly different with non mutant line check DR 7 based on LSI test at level of 5%, i: Significantly different with non mutant line check DR 8 based on LSI test at level of 5%, j: Significantly different with non mutant line check DR 9 based on LSI test at level of 5%, k: Significantly different with non mutant line check DR 10 based on LSI test at level of 5%, l: Significantly different with non mutant line check DR 11 based on LSI test at level of 5%, m: Significantly different with non mutant line check DR 14 based on LSI test at level of 5%, n: Significantly different with non mutant line check DR 16 based on LSI test at level of 5%, o: significantly different with nonmutant line check DR 18 based on LSI test at level of 5%, p: Significantly different with non mutant line check SR 17 based on LSI test at level of 5%, q: Significantly different with non mutant line check SR 25 based on LSI test at level of 5%, r: Significantly different with non mutant line check SR 52 based on LSI test at level of 5%, and ns: Not significantly different with any non mutant lines check | |
| Table 6: | Seven of the fourth generation mutant lines M4 under water stress simulation with PEG-6000 nutrition based on seedling length, chlorophyll content, root length, fresh weight of seedling and dry weight of seedling |
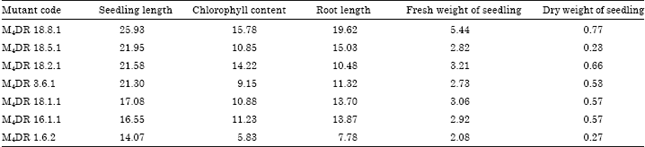 | |
The fourth generation of mutant lines of M4DR 1.6.2, M4DR 3.6.1, M4DR 16.1.1, M4DR 18.1.1, M4DR 18.2.1, M4DR 18.5.1 and M4DR 18.8.1 showed less reduction in chlorophyll content than the susceptilbe mutant lines.
Table 6 shows the selected fourth generation mutant lines based on their seedling length, root length, chlorophyll content, fresh and dry weight of seedlings. The selected mutant lines performed less reduction using the observed traits. Based on these physiological responses, these mutant lines are more tolerant to drought than the rest of the mutant lines.
Based on the result of selection both in the field experiment in Majalengka and the confirmatory test using nutrition with PEG-6000 in the laboratory, it is shown that M3DR 18.8 and its progeny of M4DR 18.8.1 (selfing of M3DR 18.8) and M3DR 18.5 and its progeny, M4DR 18.5.1 (selfing of M3DR 18.5) are tolerant mutant lines as shown by their positive index of drought and their physiological response to water stress simulation using PEG. These mutant lines could be considered for breeding program for tolerance to drought.
CONCLUSION
Gamma irradiation was found to increase the phenotypic variation and diversity of plant height, leaf number, diameter of stem, ear weight, days to tasseling and days to harvesting. In the present study, the SSR analysis was found to be a valuable DNA marker system to study genetic diversity of mutant and non-mutant lines. The mutant and non-mutant lines were clustered into two major cluster and seven sub-clusters based on a phylogenetic tree analysis using UPGMA.
There were 28 second generation mutants (M2) that showed early maturity. Based on the positive selection index of drought, the following third generation mutant lines (M3) were expected to be drought tolerant under the field experiment in Majalengka, West Java, namely M3DR 1.7, M3DR 3.3, M3DR 3.4, M3DR 3.6, M3DR 8.5, M3DR 9.2, M3DR 14.7, M3DR 16.9, M3DR 18.8 and M3DR 18.5.1. There were 7 fourth generation parental mutant lines (M4) which showed consistency in drought tolerance during confirmatory experiment in the laboratory. These fourth generation parental mutant lines included M4DR 1.6.2, M4DR 3.6.1, M4DR 16.1.1, M4DR 18.1.1, M4DR 18.2.1, M4DR 18.5.1 and M4DR 18.8.1. In addition, M3DR 18.8 and its progeny of M4DR 18.8.1 (selfing of M3DR 18.8) and M3DR 18.5 and its progeny, M4DR 18.5.1 (selfing of M3DR 18.5) are tolerant mutant lines as shown by their positive index of drought and their physiological response to water stress simulation using PEG. Those mutant lines could be considered for breeding program for tolerance to drought.
ACKNOWLEDGEMENT
The authors would like to put into words their appreciation to the Directorate General Higher Education, Ministry of Culture and Education, Republic Indonesia and Universitas Padjadjaran for the research funding through Hibah Penelitian Pasca Sarjana 2010-2012 and PUPTN 2013-2014 granted to the 1st author.
REFERENCES
- Anas and T. Yoshida, 2000. Screening of Al-tolerant sorghum by hematoxylin staining and growth response. Plant Prod. Sci., 3: 246-253.
Direct Link - Banziger, M., G.O. Edmeades, D.L. Beck and M.R. Bellon, 2000. Breeding for Drought and Nitrogen Stress Tolerance in Maize: From Theory to Practice. CIMMYT, Mexico, ISBN: 970-648-46-3, Pages: 68.
Direct Link - Wang, B., Z. Li, A.E. Eneji, X. Tian, Z. Zhai, J. Li and L. Duan, 2008. Effects of coronatine on growth, gas exchange traits, chlorophyll content, antioxidant enzymes and lipid peroxidation in maize (Zea mays L.) seedlings under simulated drought stress. Plant Prod. Sci., 11: 283-290.
CrossRefDirect Link - Bolanos, J. and G.O. Edmeades, 1996. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crops Res., 48: 65-80.
CrossRefDirect Link - Campos, H., M. Cooper, J.E. Habben, G.O. Edmeades and J.R. Schussler, 2004. Improving drought tolerance in maize: A view from industry. Field Crops Res., 90: 19-34.
Direct Link - Cassman, K.G., 1999. Ecological intensification of cereal production systems: Yield potential, soil quality and precision agriculture. Proc. Nat. Acad. Sci., 96: 5952-5959.
CrossRefDirect Link - Dreher, K., M. Morris, M. Khairallah, J.M. Ribaut, S. Pandey and G. Srinivasan, 2000. Is marker-assisted selection cost-effective compared to conventional plant breeding methods? The case of quality protein maize. Proceedings of the 4th Annual Conference of the International Consortium on Agricultural Biotechnology Research, August 24-28, 2000, Ravello, Italy, pp: 203-236.
- Edmeades, G.O., J. Bolanos, A. Elings, J.M. Ribaut, M. Banziger and M.E. Westgate, 2000. The Role and Regulation of the Anthesis-Silking Interval in Maize. In: Physiology and Modeling Kernel Set in Maize, Westgate, M.A. and K.J. Boote (Eds.). Crop Science Society of America, USA., ISBN-13: 9780891185512, pp: 43-73.
- George, M.L.C., E. Regalado, W. Li, M. Cao and M. Dahlan et al., 2004. Molecular characterization of Asian maize inbred lines by multiple laboratories. Theor. Applied Genet., 109: 80-91.
CrossRef - Goldstein, D.B., A.R. Linares, L.L. Cavalli-Sforza and M.W. Feldman, 1995. An evaluation of genetic distances for use with microsatellite loci. Genetics, 139: 463-471.
Direct Link - He, C., V. Poysa and K. Yu, 2003. Development and characterization of Simple Sequence Repeat (SSR) markers and their use in determining relationships among Lycopersicon esculentum cultivars. Theor. Applied Genet., 106: 363-373.
CrossRef - Hillel, D. and C. Rosenzweig, 2002. Desertification in relation to climate variability and change. Adv. Agron., 77: 1-38.
CrossRefDirect Link - Iqbal, M.J., N. Aziz, N.A. Saeed, Y. Zafar and K.A. Malik, 1997. Genetic diversity evaluation of some elite cotton varieties by RAPD analysis. Theoret. Applied Genet., 94: 139-144.
CrossRefDirect Link - Ludlow, M.M. and R.C. Muchow, 1990. A critical evaluation of traits for improving crop yields in water-limited environments. Adv. Agron., 43: 107-153.
CrossRefDirect Link - Messmer, M.M., A.E. Melchinger, J. Boppenmaier, R.G. Herrmann and E. Brunklaus-Jung, 1992. RFLP analyses of early-maturing European maize germ plasm. Theor. Applied Genet., 83: 1003-1012.
CrossRef - Mukhtar, M.S., M.U. Rahmanw and Y. Zafar, 2002. Assessment of genetic diversity among wheat (Triticum aestivum L.) cultivars from a range of localities across Pakistan using Random Amplified Polymorphic DNA (RAPD) analysis. Euphytica, 128: 417-425.
CrossRefDirect Link - Perera, L., J.R. Russell, J. Provan and W. Powell, 2000. Use of microsatellite DNA markers to investigate the level of genetic diversity and population genetic structure of coconut (Cocos nucifera L.). Genome, 43: 15-21.
CrossRefDirect Link - Ruswandi, D., A.L. Carpena, R.M. Lantican, D.M. Hautea, A.O. Canama and A.D. Raymundo, 2014. Genetic analysis of components of resistance and quantitative trait loci mapping of Philippine downy mildew resistance gene in maize (Zea mays L.). Asian J. Agric. Res., 8: 139-149.
CrossRefDirect Link - Vega, C.R.C., F.H. Andrade, V.O. Sadras, S.A. Uhart and O.R. Valentinuz, 2001. Seed number as a function of growth. A comparative study in soybean, sunflower and maize. Crop Sci., 41: 748-754.
CrossRefDirect Link - Wu, Y. and D.J. Cosgrove, 2000. Adaptation of roots to low water potentials by changes in cell wall extensibility and cell wall proteins. J. Exp. Bot., 51: 1543-1553.
PubMedDirect Link - Zhang, X., C. Li, L.F.H.M. Wang, H. Wang, G. You and Y. Dong, 2002. An estimation of the minimum number of SSR alleles needed to reveal genetic relationships in wheat varieties. I. Information from large-scale planted varieties and cornerstone breeding parents in Chinese wheat improvement and production. Theor. Applied Genet., 106: 112-117.
CrossRef