
Surafel Shibru
Plant Biotechnology Research Program, Melkassa Agricultural Research Center, Ethiopian Institute of Agricultural Research, P.O. Box 436 Nazareth, Ethiopia
Ampaiwan Paradornuwat
Department of Plant Pathology, Faculty of Agriculture, Kasetsart University, Bangkhen Campus, Bangkok, 10900, Thailand
Srimek Chowpongpang
Center for Agricultural Biotechnology, Kasetsart University, Kamphaeng Saen Campus/Center of Excellence on Agricultural Biotechnology (AG-BIO) PERDO/CHE
Asian Journal of Agricultural Research
Year: 2014 | Volume: 8 | Issue: 2 | Page No.: 114-121







ABSTRACT
Citrus tristerza virus (CTV) is one of the most destructive diseases in citrus growing areas of Thailand. CTV coat protein gene (CTV-CP) of Thailand MK-50 isolate which is cloned in pQE30 vector, cloned into pET160/GW/D-TOPO vector (INVITROGEN) allowing fusion of the protein with N-terminal 6XHis tag for easier protein purification and transformed into BL21 starTM (DE3) expression E. coli strain. The expression was induced by 1 mM Isopropyl-beta-D-thiogalactopyranoside (IPTG), followed by further growth for 4 h. A rabbit was immunized intradermally with a mix of recombinant CTV Coat Protein (CTV-CP) and complete Freund’s adjuvant as primary immunization and the remaining six immunization was done with mix of incomplete Freund's adjuvants within two weeks interval. The animal was bled prior to each immunization and the titer level was examined for binding recombinant CTV-CP by the indirect ELISA (Enzyme-Linked Immunosorbent assay). The collected serum was used to determine the titer as primary antibody. The result indicated that the titer level was gradually increase and reach to pick after third to sixth immunizations then lowered. The collected sera can able to detect an antigen of 20 ng at 31, 250 fold dilution and the purified polyclonal antibodies also can able to detect positive plant samples.
PDF Abstract XML References Citation
Received: January 03, 2014;
Accepted: January 25, 2014;
Published: April 11, 2014
How to cite this article
Surafel Shibru, Ampaiwan Paradornuwat and Srimek Chowpongpang, 2014. Expression of Recombinant Coat Protein (CP) of Citrus tristeza virus
for Polyclonal Antibodies Production. Asian Journal of Agricultural Research, 8: 114-121.
DOI: 10.3923/ajar.2014.114.121
URL: https://scialert.net/abstract/?doi=ajar.2014.114.121
DOI: 10.3923/ajar.2014.114.121
URL: https://scialert.net/abstract/?doi=ajar.2014.114.121
INTRODUCTION
Tristeza, the most important viral disease of citrus, is caused by Citrus Tristeza Virus (CTV) a closterovirus that is disseminated in the field by aphids. Since, October 1998, CTV is recognized as one of the most destructive disease of citrus in Thailand and causes serious damage in many production areas and results of severe economic losses (Pradornuwat et al., 1984; Paradornuwat, 2004).
The virus is phloem-limited and the particles are the longest known form of plant virus in size of 2000 nm long, consisting of a single-stranded positive-sense RNA of about 20 kb (Sekiya et al., 1991; Pappu et al., 1993). The virons contain two Capsid Proteins (CPs), a 25 kDa coat protein (Sekiya et al., 1991; Pappu et al., 1994) covering about 95% of the particle length and a small amount of a diverged 27 kDa coat protein (p27) (Pappu et al., 1994; Karasev et al., 1995). The genome encodes 12 ORFs which potentially codes for at least 17 protein products (Karasev et al., 1995; Mawassi et al., 1996).
Biological disease diagnosis using Mexican lime (Citrus aurantifolia) as an indicator plant is the common method to detect CTV infected plants which usually induce typical vein clearing and cupping on leaves. Vein clearing for most isolates of CTV can be detected within 8 weeks in healthy vigorous plants of Mexican limes grown under proper temperature conditions (Roistacher, 2004); but this method is expensive and results are obtained only after several months.
Successfully developed enzyme linked immunosorbent assay (ELISA) procedure for rapid detection of CTV was a momentous breakthrough (Bar-Joseph et al., 1979). Detection of CTV by ELISA opened the door for large scale indexing to determine the distribution of CTV within a grove, area, region or country (Garnsey et al., 1993).
Polyclonal antibodies can be produced either using CTV-Coat Protein (CTV-CP) or purified viral particles from CTV infected midrib as antigen to detect the virus. In comparisons, the virus purification from infected midrib are a laborious procedure, yields low amount and usually contaminates with plant proteins. The recombinant CTV-CP is antigenic when purified virus is injected into animal, resulted in highly specific polyclonal antibodies without reaction with healthy plant (Nurhadi and Sulaiman, 2003). In 2004 CTV-cp of 8 of Thailand isolates was successfully purified by one step RT-PCR from CTV infected plants midribs, the nucleotide and amino acid sequences of these isolates were compared to Florida T-36 (NC_001661) isolates, showed no deletion, insertion and/or frame shift mutation (Paradornuwat et al., 2004). The expression of CTV-cp gene were done in many research works but the expression and purification techniques of Thailand isolates were not studied before. Therefore, in this experiment Thai MK-50 isolate was used for the study purpose.
MATERIALS AND METHODS
Cloning of CTV-CP gene: Oligonucleotide primers were designed based on the 5’ and 3’ ends of CTV-CP gene and pET160/GW/D-TOPO cloning vector. A forward PCR primers CTV-CP1 (5’ CAC CGA CGA AAC AAA GAA ATT GAA GAA CA 3’) containing TOPO directional cloning site (the underline) and reverse PCR primers CTV-CP2 (5’ TCA ACG TGT GTT AAA TTT CCC AAG C 3’) were used to amplify a CTV-CP using pQE30 harboring cp gene as template accomplished by high fidelity thermostable DNA polymerase Phusion® (Finnzyme), to generate a blunt ends product from Thailand MK-50 isolate (Paradornuwat et al., 2004).
A 666 bp PCR product of CTV-CP gene was recovered from TAE agarose gel by silica spin column (INVITROGEN) and ligated into pET160/GW/D-TOPO cloning vector. The recombinant plasmid was chemically transformed into TOP10 Escherichia coli competent cell. The transformant cells were recovered by culturing on 2xYT broth for 1 h at 37°C on shaker then the cells were selected on prewarmed selective plate containing 2xYT agar media with 100 μg mL-1 ampicillin and incubated overnight at 37°C. Six grown colonies were selected and cultured in test tube containing 2xYT broth media with 100 μg mL-1 ampicillin at 37°C for overnight on shaker; in the following day plasmid DNA were extracted by alkaline lysis, the correct orientation of insertion was analyzed by PCR using forward primer T7F:- 5’ TAA TAC GAC TCA CTA TAG GG 3’ and reverse primer CTV-CP2:- 5’ TCA ACG TGT GTT AAA TTT CCC AAG C 3’ and sent for sequencing. The plasmid with correct orientation was subsequently introduced into E. coli BL21 StarTM (DE3) and selected on the same media as mention above.
Optimization and large scale production of recombinant protein: Two selected colonies were cultured on 2xYT broth containing 100 μg mL-1 ampicillin overnight at 37°C on shaker, on the following day new 2xYT media was inoculated with 10% of overnight grown culture and incubated at 37°C on shaker, to monitor the optimum length of time and production capacity of recombinant protein after induction with 1 mM IPTG.
The large scale production of recombinant protein was done using four one liter flasks containing 125 mL of 2xYT broth media containing 100 μg mL-1 ampicillin and protein induction was done by addition of 1 mM IPTG followed by induction for optimum period. Cells were collected by centrifugation, lysed by lysozyme and frozen at -45°C for 1 h and additional lysed by sonicator to shear DNA and made the solution runny. The produced recombinant protein was purified using QIAGEN Ni-NTA resin using denatured protein purification procedure, using 8M urea buffer in order to fully exposed the 6xHis tag protein will and improve binding to the Ni-NTA matrix ultimately to maximize the efficiency of the purification and reduced potential binding of nonspecific protein (QIAGEN, 2003). The concentration of purified recombinant protein was determined using SDS-PAGE using Bovine Serum Albumin (BSA) as a standard solution.
Immunization and titer determination: Five hundred microgram of purified CTV-CP was mixed with equal volume of complete Freund’s adjuvant and injected intradermally to a rabbit as a primary immunization. Two weeks after primary immunization the same amount of purified CTV-CP was mixed with equal amount of incomplete Freund’s adjuvant and injected to the same rabbit intradermally as booster immunization up to six times with two weeks interval. Prior to primary immunization and subsequent immunizations the rabbit was bled in order to determine the titer level. The titer was analyzed by indirect ELISA using 100 ng the purified CTV-CP as coating antigen. The intensity of the color change in each well was read at 405 nm and reading greater than 0.5 considered as positive. The collected serum was purified using ammonium sulfate.
Determination of antibody specificity using ELISA and Reverse Transcription Polymerase Chain Reaction (RT-PCR): To analyze the specificity of the purified polyclonal antibodies, citrus sample was collected and ground with liquid nitrogen and mixed with 50 mM carbonate coating buffer into 1 g to 9 mL ratio and used to coat on microtiterplate wells.
The half of ground tissue was used to extract RNA using QAIGEN RNeasy kit, about 50 mg of ground tissue from the same citrus leaves used for ELISA was mixed in 450 μL of RLT and then processed as described in the Qiagen kit protocol (QIAGEN, 2008). The quality of RNA was analyzed with 1% agarose gel. The isolated RNA was used for RT-PCR using CTV-CP1 and CTV-CP2 PCR primers. Moreover the specificity of the produced antibody against CTV was determined, by diagnosing leaves samples collected from pineapple infected by Pineapple mealybug wilt-associated virus form Closterovirus, papaya infected by Papaya ringspot virus from Potyvirus and Tomato yellow leaf curl virus from Geminivirus infected plant with the produced antibodies.
RESULTS AND DISCUSSION
CTV-CP gene from MK-50 isolate was successfully amplified with CTV-CP1 and CTV-CP2 primers. The amplified gene was run on 1% agarose gel along with 1 kb DNA marker and the gel photograph showed that the amplified PCR product is approximately 666 bp (Fig. 1). This amplified gene was cloned into pET160/GW/D-TOPO cloning vector and transformed into TOP10 E. coli for maintenance of the plasmid. From 6 selected transformed colonies, two of them miss correct orientation of insertion, when they were analyzed with PCR techniques (Fig. 1).
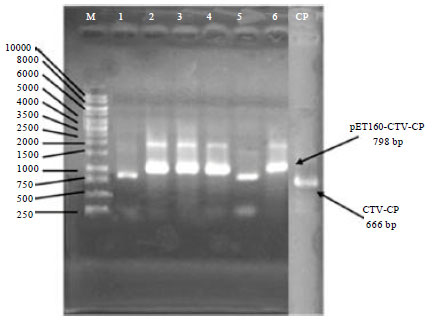 | |
| Fig. 1: | PCR analysis of six transformed colonies; M: 1 kb DNA ladder, 1-6: Transformed colonies CTV-CP into pET160/GW/D-TOPO vector amplified by T7F and CTV-CP2 primers, CP: CTV-CP gene amplified by CTV-CP1 and CTV-CP2 primers |
Two colonies with appropriate direction of insertion were chosen to study the expression capacity of CTV-CP gene without the expression cell and the result indicated that there is no any expression of CTV-coat protein, therefore, to obtain optimum expression of CTV-CP gene, the selected two colonies were transformed into BL21 StarTM (DE3) E. coli.
The sequence analysis result also showed that there is no deletion or insertion of bases comparing to the sequence of MK-50 isolates (Paradornuwat et al., 2004). Two transformed colonies from BL21 StarTM (DE3) E. coli were also selected to study the expression capacity of CTV-CP gene. The result revealed that inducing the cell for 4 h or more after the addition of 1 mM IPTG give optimum amount of recombinant protein (Fig. 2). Since, the difference between after 4, 6 h and overnight expression was negligible, 4 h incubation after addition of 1 mM IPTG was used for large-scale production.
Since, the recombinant coat protein contains 6xHistidine amino acids tag, it is suitable to purify the large-scale produced protein using Ni-NTA resin column to attain a highly purified CTV recombinant coat protein used as an antigen (Invitrogen, 2004). This purification protocol was very simple but highly sensitive to pH value of the buffers. Therefore, the pH of buffers used for the purification should be adjusted 30 min prior to use. The molecular weight of purified recombinant protein is approximately 30 kDa including the 44 amino acids sequences of pET160/GW/D-TOPO vector (Fig. 2) and when these amino acids were subtracted from the recombinant coat protein, the molecular weight will be 25 kDa which is similar to previous research work (Nurhadi and Sulaiman, 2003).
The purified protein was dialyzed and the yield of the purified recombinant protein was initially tried to estimate by Bradford’s reagent but due to the protein was purified in denaturing form, it hardly dissolve in the solution and this create difficulty to estimate the concentration of purified protein.
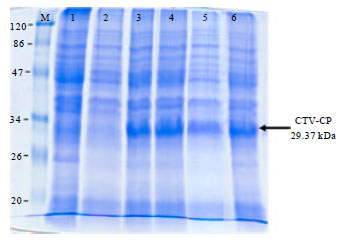 | |
| Fig. 2: | Expression of CTV coat protein cloned to pET160/GW/D-TOPO vector and transformed into BL21 StarTM (DE3) E. coli expression, M: Protein marker, 1: Overnight non-induced, 2: 2 h non-induced, 3: 2 h induced, 4: 4 h induced, 5: 6 h induced and 6: Overnight induced |
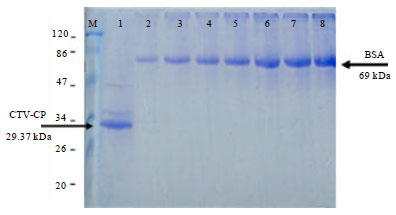 | |
| Fig. 3: | Concentration of purified recombinant protein using SDS-PAGE; M: Protein marker, 1: Purified recombinant protein; 2-8 BSA in mg mL-1 2: 0.2, 3: 0.4, 4: 0.6, 5: 0.8, 6: 1.0, 7: 1.2, 8: 1.4 |
Since, this troublesome, it was analyzed by SDS-PAGE using BSA as a standard solution. The concentration of purified recombinant protein is approximately 1 mg mL-1 and from 1 L culture (Fig. 3). A total 4.3 mg CTV recombinant coat protein was recovered from 1 L of bacterial culture which means the total amount of recombinant protein purified from a litter of culture is enough to immunize rabbit (1 primary injection and 6 booster injection in each 500 μg of antigen was used).
The ELISA result to determine the titer level indicated that the titer level increased gradually and reach to the pick 156, 250 fold dilution after the third immunization and this level remain constant up to the fifth immunization then lowered to 31, 250 fold dilution and remains constant up to the end of the experiment (Fig. 4). When we analyze the sensitivity of the antibody with serum of high titer level, it can able to detect an antigen of 20 ng well-1 concentration at 31, 250 fold dilution; when the antigen concentration level reduced to 4 ng well-1, it can be detected at 6250 dilution (Fig. 5). Moreover after purifying the antibodies with 60% ammonium sulfate; leaves samples from two mandarin plants from Chiang Mai province which showed typical CTV symptom and 1 sample from pummelo plants of Kasetsart University which didn’t show symptoms of CTV, were also analyzed by ELISA using purified antibody at 5000 fold dilution and RT-PCR procedure.
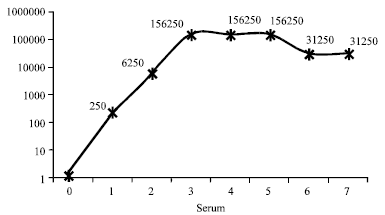 | |
| Fig. 4: | Rabbit antibody titer level using purified 100 ng well-1 CTV-CP as coating antigen based on five folds dilution at OD405 |
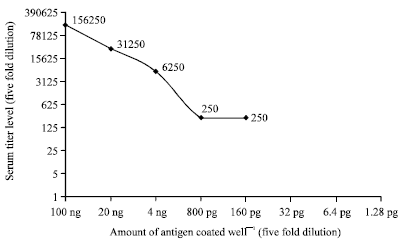 | |
| Fig. 5: | Polyclonal antibodies sensitivity test using CTV-CP as coating antigen at different concentration and serum collected on 10th week as primary antibody at OD405 |
The OD405 reading of samples indicated that plant from Kasetsart University is negative (0.127), whereas samples from Chiang Mai province are positive (0.764 and 1.036) on which the negative control (distilled water) is 0.086. The RT-PCR result also correlated with the ELISA result in which two samples collected from Chiang Mai province were positive and the other sample from Kasetsart University was negative.
Additional 21 leaves samples from lime plant which showed decline, leaf cupping and minor vein clearing symptoms, were also collected from Samut Sakhon province and analyzed by ELISA. The ELISA result revealed that 20 out of 21 collected samples were positive to CTV infection with OD405 reading greater than 0.245 in which the negative control (distilled water) is 0.098. The negative plant sample by ELISA diagnosis and other three plant samples were randomly selected and analyzed by RT-PCR. All plant sample including the sample which was negative by ELISA, were positive to CTV infection when analyzed by RT-PCR (Fig. 6).
Even though the sensitivity of the produced antibody is low, it is highly specific to CTV since both the results obtained from ELISA using infected and non infected plant materials correlated with RT-PCR.
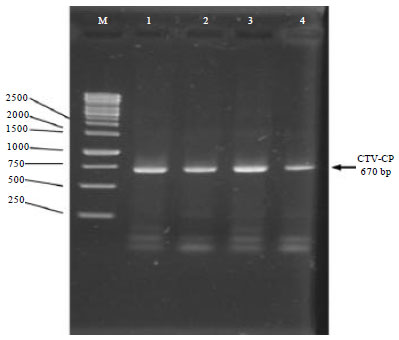 | |
| Fig. 6: | RT-PCR result for plant samples collected from Samut Sakhon province (M: 1 kb DNA marker, 1, 2 and 4: Positive sample with ELISA, 3: Negative sample with OD405 = 0.201) |
Moreover, the CTV-CP antisera didn't react with plant samples collected from infected plants from pineapple with Pineapple mealybug wilt-associated virus (PMWaV) form Closterovirus, Papaya ringspot virus (PRSV) from Potyvirus and Tomato yellow leaf curl virus from Geminivirus.
CONCLUSION
Production of polyclonal antibodies using recombinant coat protein of CTV is uncomplicated method which gives a highly specific polyclonal antibodies that don’t cross react with plant protein as compared to using purified viral particles from CTV infected midrib. The sensitivity of the produced polyclonal antibodies against CTV-CP of Thai MK-50 isolate was assessed by coating (100, 20 and 4 ng, 800, 160, 32, 6.4 and 1.28 pg) of antigen per well. The ELISA result indicated that only 100 ng antigen coated wells was detected with maximum of 156, 250 fold dilution; when the amount of the coated antigen was reduced by five fold dilution (20 ng well-1), the antibody dilution level able to detect the antigen also reduced by five fold dilution and become 31, 250 fold dilution. When we further diluted to the antigen by five fold (4 ng well-1), the titer level of antibody to detect the antigen lowered to 6250 fold dilution, for antigen concentration 800 and 160 pg was detected by 250 fold dilution. This result revealed that, the sensitivity of the produced polyclonal antibodies is low and it could be difficult to detect the pathogen in the season when the titer concentration of the pathogen in plant tissue is reduced. ELISA result for samples collected from infected and uninfected plant also correlate with the RT-PCR result. The produced antibody can able detect the pathogen during a season when the pathogen titer level in plant is higher. Therefore, in order to enhance the sensitivity of polyclonal antibodies produced against CTV-CP, the antigenicity property of other Thailand CTV isolate should be studied, in parallel with the antigenicity level of other viral protein including CTV minor coat protein, in order to get a highly sensitive antibodies that can detect the pathogen at lower titer level.
ACKNOWLEDGMENTS
The authors would like to thank, Ethiopian Ministry of Agriculture and Rural Development, Rural Capacity Building Project for their financial support to cover costs of the research expense.
REFERENCES
- Bar-Joseph, M., S.M. Garnsey, D. Gonsalves, M. Moscovitz, D.E. Purcifull, M.F. Clark and G. Loebenstein, 1979. The use of enzyme-linked immunosorbent assay for detection of citrus tristeza virus. Phytopathology, 69: 190-194.
Direct Link - Karasev, A.V., V.P. Boyko, S. Gowda, O.V. Nikolaeva and M.E. Hilf et al., 1995. Complete sequence of the citrus tristeza virus RNA genome. Virology, 208: 511-520.
CrossRef - Nurhadi, K.S. and I. Sulaiman, 2003. Production of polyclonal antibody to the coat protein of citrus tristeza virus in chicken eggs. Indonesian J. Agri. Sci., 4: 18-26.
Direct Link - Pappu, H.R., S.S. Pappu, R.F. Lee, M. Cambra, P. Moreno, S.M. Garnsey and C.L. Niblett, 1994. The molecular basis for the antigenic diversity of CTV Implications for virus detection. Proc. Fla. State Hort., 107: 8-12.
Direct Link - Pappu, H., S. Pappu, C. Niblett, R. Lee and E. Civerolo, 1993. Comparative sequence analysis of the coat proteins of biologically distinct citrus tristeza closterovirus isolates. Virus Genes, 7: 255-264.
CrossRef - Sekiya, M.E., S.D. Lawrence, M. McCaffery and K. Cline, 1991. Molecular cloning and nucleotide sequencing of the coat protein gene of citrus tristeza virus. J. Gen. Virol, 72: 1013-1020.
PubMed