
M.N. Ndjiondjop
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
P.A. Seck
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
M. Lorieux
IRD/CIAT, Agrobiodiversity and Biotechnology Unit, International Center for Tropical Agriculture, AA6713, Cali, Colombia
K. Futakuchi
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
K.N. Yao
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
G. Djedatin
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
M.E. Sow
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
R. Bocco
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
F. Cisse
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
B. Fatondji
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
Asian Journal of Agricultural Research
Year: 2012 | Volume: 6 | Issue: 4 | Page No.: 144-157







ABSTRACT
Rice varieties response to drought has been extensively studied and many lines have been released, but identifying new tolerant lines is still a challenge for scientists due to the complexity and the specificity of this constraint over environments. Three sets of field experiments were conducted between 2006 and 2008 at Africa Rice Center research station, Togoudo, Benin to evaluate the effect of drought on some traits of rice (Oryza sp.). Three genotype types including 202 interspecific lines, from a cross between WAB56-104 (O. sativa subsp. japonica) and CG14 (O. glaberrima), adapted to upland conditions, 60 chromosome segment substitution lines made for lowland conditions and 211 accessions of O. glaberrima Steud., were evaluated using a split plot design replicated twice or thrice and an alpha lattice design with four blocks. There was a consistent negative effect of drought on plant height and grain yield across genotypes’ drought-tolerance levels and across genotype/types. Plant height and grain yield were more reduced for sensitive genotypes than for moderately tolerant and tolerant genotypes. Flowering and maturity were consistently delayed across genotype types and tolerance levels. Mean delays of 6.5, 21.8 and 9.4 days were observed for start, 50 and 100% flowering, respectively. Maturity was also delayed, with consistency across genotype types. However, no clear picture of drought effect on flowering and maturity was observed in terms of differences among drought-tolerance levels. The effects of drought on the number of tillers and on leaf temperature were not consistent. Plant height and grain yield showed the clearest differences between genotype-tolerance levels. Genotypes 151-3-8, 104-3-5, 116-2-4, 117-2-6, MPL-15-3, MPL-202-3, SENL-21-2, SENL-10-1, SENL-17-2, SENL-26-3, TOG5691, TOG6679 and TOG5591 yielded higher than the parents and checks.
PDF Abstract XML References Citation
Received: February 06, 2012;
Accepted: June 25, 2012;
Published: September 03, 2012
How to cite this article
M.N. Ndjiondjop, P.A. Seck, M. Lorieux, K. Futakuchi, K.N. Yao, G. Djedatin, M.E. Sow, R. Bocco, F. Cisse and B. Fatondji, 2012. Effect of Drought on Oryza glaberrima Rice Accessions and Oryza glaberrima Derived-lines. Asian Journal of Agricultural Research, 6: 144-157.
DOI: 10.3923/ajar.2012.144.157
URL: https://scialert.net/abstract/?doi=ajar.2012.144.157
DOI: 10.3923/ajar.2012.144.157
URL: https://scialert.net/abstract/?doi=ajar.2012.144.157
INTRODUCTION
Drought is a worldwide constraints affecting rice production by seriously influencing grain production and quality (Passioura, 1996, 2007). It is unevenly predictable over years and environments (Lafitte et al., 2002a). Fukai et al. (1999) provided four justifications for this inconsistency: (1) interaction between photosynthesis and transpiration processes in low water regime which limit increase in dry matter production and yield; (2) lack or weak availability of nutrient under water stress; (3) large genotype-by-environment interaction (GxE) for yield, causing yield gap performance in different environments, which is fortunately overcome by more testing in different locations and years and (4) different types of drought coupled with different traits that may therefore, be required for each drought type. However, the combination of new drought-screening facilities and methods (O’Toole, 2004), coupled with the existence of linkage map in rice (McCouch et al., 1988) and the breakthrough in map saturation (Causse et al., 1994; McCouch et al., 2002; Wu et al., 2002) are given more insight in the management of drought-resistance traits while enhancing the selection of drought-resistant genotypes (Kamoshita et al., 2008).
The drought problem is also becoming more severe as the human population increases and global climate changes. In addition, because of shrinking area of arable rice lands, change in employment pattern and its ease of cooking, rice (Oryza sativa L.) has become a staple crop for about half of the world’s population. Among the major constraints to rice cultivation, water shortage significantly increases the vegetative growth rate and reduces grain yield (Tao et al., 2006). It affects more than half of the rice production area worldwide (Bouman et al., 2005). Drought stress during each of the rice growth stage (vegetative growth, flowering and terminal stage) causes spikelet sterility that lead to unfilled grains (Kamoshita et al., 2004; Shahryari et al., 2008). Usually, drought reduces the grain-filling period and induces early senescence by redirecting remobilization of assimilates from the straw to the grains (Plaut et al., 2004). Similarly to plant response to drought and the manner in which drought reduces grain yield, drought itself is a complex physical, biochemical and physiological process (Passioura, 2007; Xoconostle-Cazares et al., 2010) which need different mechanisms of avoidance by plants when they encounter drought (Levitt, 1980). The understanding of these mechanisms to cope with water deficit as well as their occurrence would offer a great insight in designing accurate methods for drought management using molecular tools (Gorji et al., 2010). Moreover, identifying or developing drought-tolerant rice varieties and breeding lines for use as donors in breeding (Manickavelu et al., 2006) and gene discovery could provide a practical solution for drought alleviation (Serraj and Atlin, 2008).
During the last two decades, AfricaRice scientists were therefore embarked, in a breeding program that ended up to a new plant type namely New Rice for Africa (NERICA), developed for upland and lowland ecologies, well adapted to rice major constraints including drought tolerance. The rustic characters of CG14, an O. glaberrima from Casamance in Senegal, that confer resistance to several abiotic (drought, salt and high and low temperatures) and biotic stresses (Rice yellow mottle virus, Heterodera and Meloidogyne nematodes, African gall midge insect, rice blast fungus, Xanthomonas bacteria and the Striga parasitic weed), were successfully combined to the high yield of three O. sativa to develop 18 released upland NERICAs. Molecular studies have been conducted by Ndjiondjop et al. (2006) and Semagn et al. (2007) to, respectively assess the genetic relationship among breeding lines, to know the contribution of each parent to the NERICAs lines and to assess the genetic relationship among all the released 18 and 48 upland and lowland NERICAs. However, no fine gene study has been implemented up to date and the nature of the genes responsible of the NERICAs’ performances is still unknown, even though these genes are proven to come from their glaberrima parent. In order to elucidate the genetic mechanism of drought tolerance of the NERICAs, development of a set of Chromosome Segment Substitution Lines (CSSL) derived from the rice cross IR64/TOG5681 and upland NERICAs’ sisters progenies derived from WAB56-104 and CG14 were performed.
The chromosome segment substitution lines, carrying overlapping chromosome segments of the drought tolerant variety TOG5681 in a genetic background of the elite cultivars IR64 from the International Rice Research Institute (IRRI), are also the most powerful tool to identify and accurately map quantitative traits including drought (Xu et al., 2010). Assessing these introgression lines and the glaberrima accessions according to their drought tolerant ability appears as a prerequisite to further monitor drought tolerant fine mapping genes based on CSSL (Xu et al., 2010; Zhu et al., 2009) and to widen the genetic pool of the resistance to drought, respectively. In this study, attention is focused on introgression lines, O. glaberrima accessions and NERICAs’ sisters progenies derived from cross between CG14 as donor and WAB56-104 as recurrent parent to assess some agronomic performances of these lines under drought stress.
The present study was also carried out to identify and to evaluate the drought tolerance of various rice genotypes bearing O. glaberrima alleles with a wide range of genetic variation through three sets of field experiments.
MATERIALS AND METHODS
Genetic materials: A population of 202 backcross inbred lines developed from a cross between CG14 (O. glaberrima) as donor and WAB56-104 (O. sativa subsp. japonica) as recurrent parent, was used in the first experiment. CG14 is adapted to drought conditions and is characterized by low yield potential but several useful traits, including high weed-competitiveness (Dingkuhn et al., 1997) as a result of early vigor and high tiller number. WAB56-104 is moderately drought tolerant (Hounkpatin, 2007) and an improved upland line developed by AfricaRice, with important agronomic traits such as high yield and short growth duration. The two parents were included as checks in the experiments.
In the second experiment, 211 O. glaberrima accessions of rice were evaluated. They originated from seven West African countries: Cote d’Ivoire, Ghana, Liberia, Mali, Nigeria, Senegal and Sierra Leone. The following checks were used in the experiment: (a) three local landraces: PADCKONO, DCKONO and Salikatato, (b) two O. glaberrima (RAM1 and RAM55) as susceptible and tolerant checks, respectively, based on results of several field experiments (Hounkpatin, 2007; Africa Rice Center (AfricaRice) 2010, unpublished) and (c) CG14 and WAB56-104 as additional checks.
The third experiment evaluated 60 rice genotypes made up of introgression lines (90%), their parents (3.3%): female IR64 (O. sativa) and male TOG5681 (O. glaberrima) used as checks and four of NERICA varieties (i.e., 6.7%) adapted to lowland ecology (NERICA-L varieties): NERICA-L 19, NERICA-L 20, NERICA-L 41 and NERICA-L 42. TOG 5681 is characterized by drought tolerant capacity. The four NERICA-L varieties were developed from the same parents: NERICA-L 19 and NERICA-L 20 from TOG5681/3xIR 64 and NERICA-L 41 and NERICA-L 42 from TOG5681/4xIR 64 and were also used in the study as additional checks. The introgression lines or a set of overlapping chromosome segment substitution lines were developed by the Institut de la Recherche pour le Développement (IRD). These lines are different from those used in the first experiment in that the introgression lines were developed using markers. The genetic materials used in the first and third experiments were obtained from the gene bank of AfricaRice, Togoudo, Benin and those used in the second experiment from IRD.
Experimental site: The first and second experiments were conducted in an upland environment and the third in a lowland environment at the AfricaRice research station, Togoudo, southern Benin (25 m above sea level; 6°25'N, 2°20'E). The station is located in the coastal savannah zone, with a sub-equatorial climate. Southern Benin is characterized by a long dry season (December-March), during which rice cannot be grown without supplemental irrigation, as the available soil moisture during this period is below the total evapotranspiration needs of the rice crop. There is also a water shortage, one-month dry season in southern Benin, usually experienced between mid-July and mid-August.
Experimental details: Three experiments were conducted between July 2006 and April 2008. Each of the experiments was repeated once. The first experiment was direct-seeded with three seeds per hill. Plantlets were later thinned to one healthy plant per hill soon after emergence, at a spacing of 0.20 m between and within rows. Plots were 1.0x1.5 m and were arranged in a split-plot design replicated twice, with irrigation regime as the main-plot factor and rice lines as the sub-plot factor. Within each sub-plot, the lines were randomized using an alpha-lattice design. Two irrigation regimes were used: fully irrigated up to maturity and imposition of 21 days of drought (Efisue, 2006) from 45 Days After Sowing (DAS). Plants were sprinkler-irrigated using a pipe with water from a borehole to maintain the soil above field capacity until 45 DAS, when the treatments were applied according to the experimental design. The irrigation continued in fully irrigated plots until the end of the experiments. Compound fertilizer (NPK 15-15-15) was applied at the rate of 200 kg ha-1 two weeks after sowing, followed by 40 kg N ha-1 as urea (46% N) 40 DAS.
Similar protocol was used in the second experiment with the differences that the subplots were 1.5x1.5 m, the distance between plants was 25 cm and between subplots, 75 cm. The experimental plots were fertilized with 15 kg NPK (70-40-40) ha-1 at two and three weeks after sowing, followed by urea application at the rate of 40 kg ha-1 (18.4 kg N ha-1). The plots were arranged in a lattice design with four blocks.
In the third experiment, a split-plot design with three replicates in which the blocks were arranged in an Alpha lattice design, was used. Plots were 1x1 m with one genotype each. The distance between two consecutive lines and two consecutive plants on a line was 20 cm. Plots were fertilized with 40-40-40 kg ha-1 of NPK (15-15-15) and then with urea (30 kg ha-1 N), 20 DAS. Decis (Deltamethrin) and Mancozeb (Dithiocarbamate) were applied, respectively, as insecticide at 40 mL/15 L of water and as fungicide at 80 g/15 L of water. The three experiments were kept weed-free by regular hand weeding and bird damage was controlled using bird scares. Plants were harvested at maturity (four months after sowing).
Measurements: Data on rainfall, evapotranspiration, temperature and relative humidity during the experimental periods were recorded by the AfricaRice weather section. In all experiments, data were collected on agronomic traits of the plants following the Standard Evaluation System (SES) for rice (INGER and IRRI, 1996). Four plants were randomly selected and labeled in each experimental plot for the measurement of plant height, which was taken as the distance from the soil surface to the tip of the tallest panicle of each plant. Greenness of leaves (SPAD) was recorded using a SPAD meter on the last fully developed leaf. In all cases, the SPAD meter (SPAD-502, Konica Minolta) was placed in the middle of the widest part of the leaf. Leaf temperature was recorded using a handheld infrared thermometer (Model AG-42, Telatemp Corporation, Inc., Fullerton, CA, USA) placed in the middle (the widest part) of the last fully developed leaf. Leaf temperature and SPAD were recorded between 9 and 10 am. The following traits were also recorded: number of tillers per plant; number of days from sowing to exertion of panicles (flowering) in the first plants and in 50 and 100% of the plants in a plot; the grain yield per plant, determined after drying at 50°C for three days, with moisture content adjusted to 14%. Maturity was recorded as the number of days from sowing to grain ripening (defined as the point at which 85% of grains were mature). Leaf tip burning and leaf rolling were evaluated visually in all experiments at the end of the drought treatment both using scales of 0-9.
Statistical analyses: Analysis of variance (ANOVA) and Pearson correlation analysis were performed using SAS (version 9.1) statistical software (SAS Institute, 2003). The Fisher’s Least Significant Difference (LSD) test was used to test the difference between plant tolerance levels. Genotypes were classified into three groups (sensitive, moderately tolerant and tolerant) according to their leaf rolling and leaf tip burning potential using a Principal Component Analysis (PCA).
RESULTS
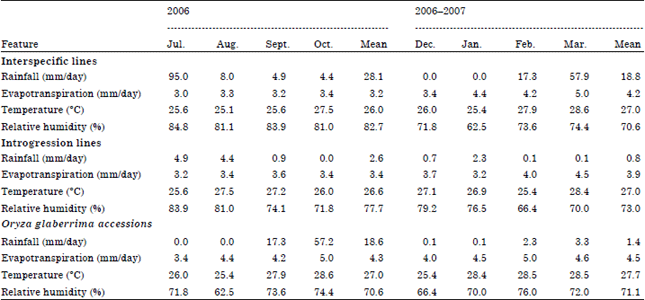
Rainfall, evapotranspiration, air temperature and relative humidity varied across seasons and therefore, varied across experiments and genotype groups (Table 1). Little rain (mean of 0.8-2.6 mm/day) was recorded during the experiments on introgression lines and second-season O. glaberrima accessions. More rain (mean of 18.6 to 28.1 mm day-1) was recorded during experiments on interspecific lines and during the first-year experiment on O. glaberrima accessions. The mean daily evapotranspiration during the experiments on the different genotype types ranged between 3.2 and 4.5 mm. Mean air temperature value ranged between 26.0 and 27.7°C and mean relative humidity between 70.6 and 82.7%.
Variations were observed across plant traits and irrigation regimes for each tolerance level (Table 2). The rice genotype types evaluated in this study showed tremendous variation in response to water level. For introgression lines under fully irrigated conditions, values for plant height, flowering (start, 50 and 100%) and maturity were significantly higher (p≤0.05) for sensitive genotypes than for tolerant and moderately tolerant genotypes while values for number of tillers, leaf temperature, grain yield and SPAD were similar across tolerance levels. Under drought conditions, the same five mentioned-above traits, in addition to number of tillers were significantly different for sensitive genotypes compared with the other two types. For interspecific lines under fully irrigated conditions, values for plant height, number of tillers and time to flowering (50%) were significantly higher for sensitive lines than for moderately tolerant and tolerant lines. However, under drought, only plant height and number of tillers were significantly higher for sensitive lines.
| Table 1: | Weather data recorded during three experiments conducted at AfricaRice research station, Togoudo, Benin in 2006-2008 |
 | |
| Table 2: | Effect of drought on three rice genotype types according to their drought-tolerance levels evaluated under field conditions at AfricaRice, Togoudo, Benin, 2006-2008 |
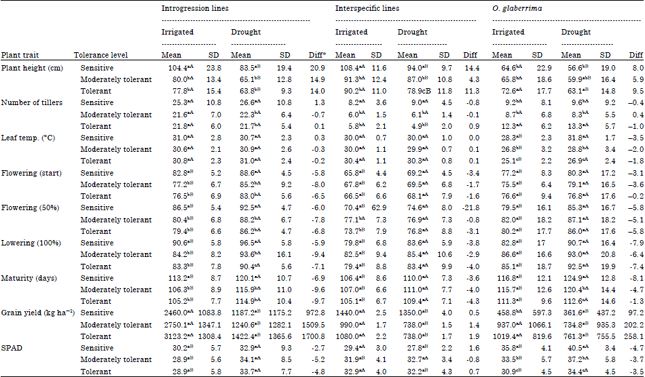 | |
| *Difference between values for fully irrigated and drought conditions, SD: Standard deviation. For a given plant trait, values with different lower-case letters are significantly different according to the Fishers’ LSD test, Values with different capital letters for the same trait for a given genotype type indicate significant differences between trait values under fully irrigated and drought conditions | |
In both irrigation regimes, values for leaf temperature, flowering (start and 100%), maturity, grain yield and SPAD were similar across tolerance levels. For O. glaberrima lines, leaf temperature and SPAD were significantly higher for sensitive genotypes than for other genotype types under fully irrigated and drought conditions. Under drought, sensitive lines also matured later. However, under both irrigation regimes, plant height and number of tillers were significantly higher for tolerant lines while only flowering was similar across the genotype’s drought-tolerance levels.
Plant height and grain yield were lower under drought than under fully irrigated condition during the three experiments for all plant tolerance types. The reduction of plant height under drought ranged between 4.3 and 20.9 cm according to the genotype type and the genotypes’ drought-tolerance level. Greatest grain yield reduction due to drought was observed for introgression lines (972.8-1700.8 kg ha-1) while the lowest yield reduction was noted for interspecific lines (0.5-1.9 kg ha-1). Flowering (start, 50 and 100%) and maturity were retarded under drought as compared to under full irrigation for introgression lines, interspecific lines and O. glaberrima accessions. This was observed for all drought-tolerance levels except for 50% flowering for interspecific lines with the moderately tolerant level. Mean delay in start of flowering ranged between 0.2 and 8.0 days. Fifty percent of plants in a plot flowered within a delay of 0.8-21.8 days due to drought. The mean difference between flowering (100%) under drought and fully irrigated conditions ranged from 2.9-9.4 days. Maturity was retarded by 1.3-9.7 days under drought as compared to fully irrigated condition. The number of tillers was not significantly affected by drought for any genotype type except for tolerant interspecific lines with the tolerant level. Leaf temperature of introgression and interspecific lines was not affected by drought; however, it was higher under drought than under full irrigation for O. glaberrima accessions, with an increase of 1.8-3.5° under drought as compared to fully irrigated condition.
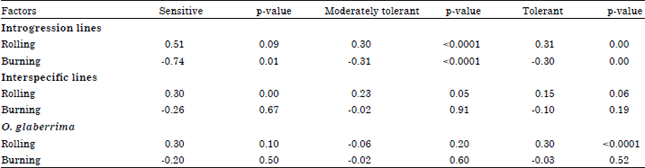
The relationship between leaf rolling and leaf tip burning and grain yield under drought was strongest for moderately tolerant introgression lines for leaf rolling and leaf tip burning and also for tolerant O. glaberrima for leaf rolling (Table 3). Positive relationship was observed between rolling and grain yield under drought for all genotype types, excluding the moderately tolerant O. glaberrima accessions. The relationship between leaf tip burning and grain yield under drought was negative for all genotype types even though not always significantly.
Some interspecific lines had better grain yield performance than the parents WAB56-104 and CG14 (Fig. 1a). For example, grain yields of lines 151-3-8 and 104-3-5 were higher under both drought and fully irrigated conditions than those of parent WAB56-104. Similarly, grain yields of 116-2-4 and 117-2-6 were higher than that of CG14 under drought and under both drought and fully irrigated conditions, respectively.
| Table 3: | Relationship between leaf rolling and leaf tip burning ability and grain yield under drought of rice genotypes evaluated under field conditions at AfricaRice, Togoudo, Benin, 2006-2008 |
 | |
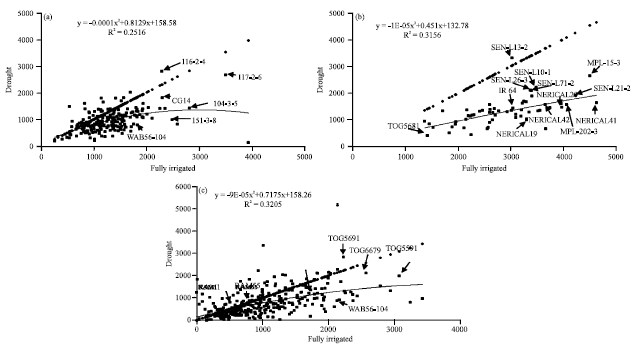 | |
| Fig. 1(a-c): | Grain yield of three genotype types, (a) interspecific lines (202) adapted to upland conditions, (b) introgression lines (60) made for lowland conditions and (c) O. glaberrima (211 accessions)-grown under fully irrigated and drought conditions at AfricaRice, Togoudo, Benin, 2006-2008 |
Grain yield of NERICA-L20, NERICA-L41, NERICA-L42, MPL-15-3, MPL-202-3, SEN-L21-2, SEN-L-10-1, SEN-L17-2 and SEN-26-3 were higher than those of the two checks TOG5681 and IR64 (Fig. 1b). The genotypes TOG5691, TOG6679 and TOG5591 gave grain yields higher under drought and irrigation conditions than the checks CG14, WAB56-104, RAM1 and RAM55 (Fig. 1c).
DISCUSSION
In this study, more weight was given to plant height, number of tillers, leaf temperature, flowering, maturity, grain yield and SPAD to evaluate drought tolerance. Although, some of the drought evaluation methods based on plant body symptoms such as plant wilting, leaf rolling and yield loss are not always appropriate for varietal screening because they are labor intensive (Hirayama et al., 2006), visible plant damage coupled with phenology are, however, believed to be a good indicator of the stress (Talebi et al., 2009; Gathu et al., 2012). Three field experiments were conducted to evaluate the drought effect on various rice genotypes. The results were consistent across experiments (with a few exceptions) for plant height, flowering (start, 50 and 100%), maturity, grain yield and SPAD. The effects on the number of tillers and leaf temperature were, however, not consistent across the experiments.
Plant height and grain yield were reduced under drought as extensively reported earlier by Lafitte et al. (2002b, 2004), Srinivasan (2005), Xu et al. (2005), Bernier et al. (2007), Ikeda et al. (2007), Ganapathy et al. (2007) and Liu et al. (2008). Under drought, plant development is reduced as a consequence of (a) poor root development; (b) reduced leaf-surface traits (form, shape, composition of cuticular and epicuticular wax, leaf pubescence and leaf color), which affect the radiation load on the leaf canopy; (c) delay in or reduced rate of normal plant senescence as it approaches maturity and (d) inhibition of stem reserves (Blum, 2002). The negative effect of drought on plant height and grain yield concurs with the results of previous studies (Efisue, 2006; Hounkpatin, 2007; Ndjiondjop et al., 2010). In this study, the observation was consistent across experiments, indicating that the traits are good indicators that could be used in drought screening tests as claimed by several authors (Talebi et al., 2009; Mitra, 2001; Sangtarash, 2010) that grain yield alone is not a good measure of drought tolerance even though it could apply to drought escape. This assertion broadly confirmed that the intensity of drought effect on both traits varied with the genetic materials (Loresto et al., 1976; Chang et al., 1982; Mackill et al., 1996; Ali, 2011).
The study also indicated strong relationship between grain yield under drought and leaf rolling and leaf tip burning for moderately tolerant introgression lines. Likewise, the same trend was observed between grain yield under drought and leaf rolling for tolerant O. glaberrima. Similar results have been reported by Ali et al. (2011) on maize, Acuna et al. (2008) on rice and Talebi et al. (2009) on wheat. This explains the role of yield increase of leaf rolling and leaf tip burning potential of genotype. Indeed, it has been demonstrated that photosynthesis rate and productivity is majorly critical in dry weight matter and subsequently in yield production (Poshtmasari et al., 2007; Cha-Um et al., 2007). Therefore, genotypes with enrolled or tip burned leaf forage less for light, close their stomata to maintain their inner moisture content and consequently their photosynthetic rate and productivity decrease (Gollan et al., 1986; Turner, 1986).
The delay in flowering under drought is a consequence of a reduction in plant dry-matter production and a delay in panicle exertion (Murty and Ramakrishnayya, 1982). Novero et al. (1985) reported that the delay in flowering depends on the intensity, time and period of drought. Wopereis et al. (1996) observed longer flowering delay when drought occurred during early tillering than when it occurred in mid-tillering stage. Pantuwan et al. (2002) made similar observations and concluded that under prolonged drought, flowering time is an important determinant of rice grain yield. The maturation stage, which is regarded as the period between anthesis and harvest, is also delayed as a result of delayed flowering or when drought appears after flowering. The delays in flowering and maturity could also be considered as good indicators in drought screening tests since the effect of drought on the traits was consistent. The differences observed among genotypes in the delays might be a result of differences in plant water status in the genotypes during the drought and consequently in the drought escape and avoidance potential of the genotypes. Audebert et al. (2002) stated that drought tolerance is complex, involving several interacting physiological, phenological and morphological mechanisms for escape, avoidance, resistance and recovery. They stated that, depending on the drought type, drought tolerance may be controlled by simple plant traits that can lead to simple and cheap breeding programs. To minimize the effect of drought on rice production, drought-tolerant genotypes will therefore be an appropriate farmer-friendly, seed-based technology that will be easy to disseminate.
The study identified genotypes 151-3-8, 104-3-5, 116-2-4, 117-2-6, MPL-15-3, MPL-202-3, SEN-L21-2, SENL-10-1, SENL-17-2, SENL-26-3, TOG5691, TOG6679 and TOG5591 as with grain yield higher than that of the parents and checks used in the experiments under the two irrigation conditions. These genotypes could serve for the aforementioned purpose and could also serve as potential donors and could be introduced into large scale evaluation programs aiming at designing drought tolerant lines. In addition, genotypes MPL-15-3 and MPL-202-3 could be used for further drought tolerant gene fine mapping and gene identification.
The results, obtained on the number of tillers and the leaf temperature, were not consistent across the experiments. With the exception of the tolerant interspecific lines, drought stress did not impact the number of tiller. However, this negative effect of drought, observed on the number of tillers, corroborates results of Efisue (2006), Mostajeran and Rahimi-Eichi (2009) and Ndjiondjop et al. (2010), who explained the phenomenon as a result of poor plant development. Apart from O. glaberrima accessions, which leaf temperature increased under drought condition, leaf temperature was surprisingly not significantly different for both interspecific and introgression lines regardless of the tolerance level and the water regime. The increase of leaf temperature under drought (in some genotype types) concurs with Ingram et al. (1990), who observed a significant correlation between leaf temperature and grain yield under drought. Leaf temperature has been proposed as a good indicator of plant stress level for its association with plant water status under drought (Serraj et al., 2009). Blum et al. (1978), Ishiahara and Saito (1983) and O’Toole et al. (1984) justified this assertion by the rise in the leaf temperature associated with the decrease of the transpiration rate due to the closure of stomata before the occurrence of macroscopical damage. As about 2470 Joules are lost by the plants following transpiration of every gram of water, transpiration tends to cool the plant leaf; the water losses due to transpiration increase with increase in leaf surface area (Arraudeau, 1998). Maurya and O’Toole (1986) reported leaf temperature as a good drought-avoidance indicator, based on the principle of the cooling effect of transpiration, as high transpiration rate occurs in the leaf at lower temperature in genotypes that avoid drought.
CONCLUSION
In this study, O. glaberrima accessions were confirmed to provide useful genes for drought tolerance as reported for most biotic and abiotic stresses of rice. Once these genes are introgressed into high yielding promising lines, these lines showed improved yield and better resistant to drought. Overall, drought stress impacted significantly the traits studied of some genotypes and some of them revealed tolerance to drought, suggesting the genetic variability for drought tolerance in the investigated material. Therefore, based on this limited sample and environments, testing and selection under non-stress and stress conditions alone may not be the most effective for increasing yield under drought stress. As drought is clearly not a single and homogeneous stress, the type of drought must be defined to avoid that drought levels and timing used in a stress nursery identify lines with specific advantageous traits that might not be valuable in other drought scenarios.
ACKNOWLEDGMENTS
The research was supported by the Generation Challenge Programme project lead by the first author and AfricaRice. The authors thank Dr. Hugues Baimey for the critical reading of the manuscript and IRD for providing part of the material.
REFERENCES
- Ali, M.A., 2011. Pedigree selection for grain yield in spring wheat (Triticum aestivum L.) under drought stress conditions. Asian J. Crop Sci., 3: 158-168.
CrossRefDirect Link - Ali, Z., S.M.A. Basra, H. Munir, A. Mahmood and S. Yousaf, 2011. Mitigation of drought stress in maize by natural and synthetic growth promoters. J. Agric. Social Sci., 7: 56-62.
Direct Link - Audebert, A., F. Asch and M. Dingkuhn, 2002. Morpho-Physiological Research on Drought Tolerance in Rice at WARDA. In: Field Screening for Drought Tolerance in Crop Plants with Emphasis on Rice, Saxena, N.P. and J.C. O'Toole (Eds.). International Crops Research Institute for the Semi-Arid Tropics, India, pp: 59-60.
- Xoconostle-Cazares, B., F.A. Ramirez-Ortega, L. Flores-Elenes and R. Ruiz-Medrano, 2010. Drought tolerance in crop plants Am. J. Plant Physiol., 5: 241-256.
CrossRef - Bernier, J., A. Kumar, V. Ramaiah, D. Spaner and G. Atlin, 2007. A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice. Crop Sci., 47: 507-518.
CrossRef - Blum, A., K.F. Scheriz. R. Toler, R.I. Welch, D.T. Rosenow, D.T. Johnson and L.E. Clark, 1978. Selection for drought avoidance in sorghum using infrared photography. Agron. J., 70: 474-477.
Direct Link - Acuña, T.L.B., H.R. Lafitte and L.J. Wade, 2008. Genotype × environment interactions for grain yield of upland rice backcross lines in diverse hydrological environments. Field Crops Res., 108: 117-125.
CrossRefDirect Link - Bouman, B.A.M., S. Peng, A.R. Castaneda and R.M. Visperas, 2005. Yield and water use of irrigated tropical aerobic rice systems. Agric. Water Manage., 74: 87-105.
CrossRefDirect Link - Causse, M.A., T.M. Fultion, Y.G. Cho, S.N. Ahn and J. Chunwongse et al., 1994. Saturated molecular map of the rice genome based on an interspecific backcross population. Genetics, 138: 1251-1274.
Direct Link - Dingkuhn, M., M. Jones, D. Johnson, B. Fofana and A. Sow, 1997. Oryza sativa and Oryza glaberrima Gene Pools for High Yielding Weed Competitive Rice Plant Types. In: Breeding Strategies for Rainfed Lowland Rice in Drought Prone Environments, Fukai, S., M. Cooper and J. Salisbury (Eds.). Australian Centre for International Agricultural Research, Canberra, Australia, pp: 144-155.
- Fukai, S., G. Pantuwan, B. Jongdee and M. Cooper, 1999. Screening for drought resistance in rainfed lowland rice. Field Crops Res., 64: 61-74.
CrossRef - Ganapathy, S., S.K. Ganesh and R.C. Babu, 2007. Evaluation of genetic potential of parents and their hybrids for drought tolerance in rice (Oryza sativa L.). Int. J. Plant Breed. Genet., 1: 82-88.
CrossRefDirect Link - Gathu, E.W., E.G. Karuri and P.M.K. Njage, 2012. Physical characterization of new advanced drought tolerant common bean (Phaseolus vulgaris) lines for canning quality. Am. J. Food Technol., 7: 22-28.
CrossRefDirect Link - Gollan, T., J.B. Passioura and R. Munns, 1986. Soil water status affects the stomatal conductance of fully turgid wheat and sunflower leaves. Aus. J. Plant Physiol., 13: 459-464.
CrossRef - Gorji, A.H., F. Darvish, M. Esmaeilzadehmoghadam and F. Azizi, 2010. Application RAPD technique for recognition genotypes tolerant to drought in some of bread wheat. Asian J. Biotechnol., 2: 159-168.
CrossRefDirect Link - Hirayama, M., Y. Wada and H. Nemoto, 2006. Estimation of drought tolerance based on leaf temperature in upland rice breeding. Breeding Sci., 56: 47-54.
CrossRefDirect Link - Poshtmasari, H.K., H. Pirdashti, M. Nasiri and M.A. Bahmanyar, 2007. Study the effect of nitrogen fertilizer management on dry matter remobilization of three cultivars of rice (Oryza sativa L.). Pak. J. Biol. Sci., 10: 3425-3429.
CrossRefPubMedDirect Link - Ikeda, H., A. Kamoshita and T. Manabe, 2007. Genetic analysis of rooting ability of transplanted rice (Oryza sativa L.) under different water conditions. J. Exp. Botany, 58: 309-318.
CrossRef - Ingram, K.T., J.G. Real, M.A. Maguling, M.A. Obien and G.C. Loresto, 1990. Comparison of selection indices to screen lowland rice for drought resistance. Euphytica, 48: 253-260.
CrossRef - Kamoshita, A., R. Rodriguez, A. Yamauchi and L. Wade, 2004. Genotypic variation in response of rainfed lowland rice to prolonged drought and rewatering. Plant Prod. Sci., 7: 406-420.
CrossRefDirect Link - Kamoshita, A., C.R. Babu, N.M. Boopathi and S. Fukai, 2008. Phenotypic and genotypic analysis of drought-resistance traits for development of rice cultivars adapted to rainfed environments. Field Crops Res., 109: 1-23.
CrossRefDirect Link - Lafitte, H.R., B. Courtois and G.N. Atlin, 2002. The International Rice Research Institute's Experience in Field Screening for Drought Tolerance and Implications for Breeding. In: Field Screening for Drought Tolerance in Crop Plants with Emphasis on Rice, Saxena, N.P. and J.C. O'Toole (Eds.). ICRISAT, Patancheru, India.
- Lafitte, R., B. Courtois and M. Arraudeau, 2002. Genetic improvement of rice in aerobic systems: Progress from yield to genes. Field Crops Res., 75: 171-190.
CrossRef - Liu, G., H.W. Mei, X.Q. Yu, G.H. Zou and H.Y. Liu et al., 2008. QTL analysis of panicle neck diameter, a trait highly correlated with panicle size, under well-watered and drought conditions in rice (Oryza sativa L.). Plant Sci., 174: 71-77.
CrossRef - Mackill, D.J., W.R. Coffman and D.P. Garrity, 1996. Rainfed Lowland Rice Improvement. International Rice Research Intitute, Manila, ISBN-971-22-0071-X,.
Direct Link - Manickavelu, A., R.P. Gnanamalar, N. Nadarajan and S.K. Ganesh, 2006. Identification of important traits in rice (Oryza sativa L.) For lowland drought situation by association analyses. Int. J. Agric. Res., 1: 509-521.
CrossRefDirect Link - McCouch, S.R., G. Kochert, Z. Yu, Z. Wang, G.S. Khush, W. Coffman and S.D. Tanksley, 1988. Molecular mapping of rice chromosomes. Theor. Applied Genet., 76: 815-829.
CrossRefDirect Link - Mitra, J., 2001. Genetics and genetic improvement of drought resistance in crop plants. Curr. Sci., 80: 758-763.
Direct Link - Mostajeran, A. and V. Rahimi-Eichi, 2009. Effects of drought stress on growth and yield of rice (Oryza sativa L.) cultivars and accumulation of proline and soluble sugars in sheath and blades of their different ages leaves. Am.-Eurasian J. Agric. Environ. Sci., 5: 264-272.
Direct Link - Ndjiondjop, M.N., B. Manneh, M. Cissoko, N.K. Drame and R.G. Kakai et al., 2010. Drought resistance in an interspecific backcross population of rice (Oryza sp.) derived from the cross WAB56-104 (O. sativa) x CG14 (O. glaberrima). Plant Sci., 179: 364-373.
CrossRef - Novero, R.P., J.C. O'Toole, R.T. Cruz and D.P. Garrity, 1985. Leaf water potential, crop growth response and microclimate of dryland rice under line source sprinkler irrigation. Agric. Forest Meteorol., 35: 71-82.
CrossRef - Passioura, J., 2007. The drought environment: Physical, biological and agricultural perspectives. J. Exp. Bot., 58: 113-117.
CrossRefDirect Link - Plaut, Z., B.J. Butow, C.S. Blumenthal and C.W. Wrigley, 2004. Transport of dry matter into developing wheat kernels and its contribution to grain yield under post-anthesis water deficit and elevated temperature. Field Crops Res., 86: 185-198.
CrossRefDirect Link - Sangtarash, M.H., 2010. Responses of different wheat genotypes to drought stress applied at different growth stages. Pak. J. Biol. Sci., 13: 114-119.
CrossRefDirect Link - Serraj, R. and G. Atlin, 2008. Drought-Resistant Rice for Increased Rainfed Production and Poverty Alleviation: A Concept Note. In: Drought Frontiers in Rice: Crop Improvement for Increased Rainfed Production, Serraj, R., J. Bennett and B. Hardy (Eds.). International Rice Research Institute, Los Banos, Philippines, pp: 385-400.
- Serraj, R., A. Kumar, K.L. McNally, I. Slamet-Loedin and R. Bruskiewich et al., 2009. Improvement of drought resistance in rice. Adv. Agronomy, 103: 41-99.
CrossRef - Shahryari, R., E. Gurbanov, A. Gadimov and D. Hassanpanah, 2008. Tolerance of 42 bread wheat genotypes to drought stress after anthesis. Pak. J. Biol. Sci., 11: 1330-1335.
CrossRefPubMedDirect Link - Cha-Um, S., S. Roytakul, T. Sathung, A. Maijandang and C. Kirdmanee, 2007. Effect of exogenous glucose and abscisic acid on physiological and morphological performances of in vitro indica rice (Oryza sativa L. sp. indica). Am. J. Plant Physiol., 2: 155-166.
CrossRefDirect Link - Talebi, R., F. Fayaz and A.M. Naji, 2009. Effectiveselection criteria for assessing drought stress tolerancein durum wheat (Triticum durum Desf.). General Applied Plant Physiol., 35: 64-74.
Direct Link - Tao, H., H. Brueck, K. Dittert, C. Kreye, S. Lin and B. Sattelmacher, 2006. Growth and yield formation for rice (Oryza sativa L.) in the water-saving ground cover rice production system (GCRPS). Field Crops Res., 95: 1-12.
CrossRef - Wopereis, M.C.S., M.J. Kropff, A.R. Maligaya and T.P. Tuong, 1996. Drought stress responses of two lowland rice cultivars to soil water status. Field Crops Res., 46: 21-39.
CrossRef - Wu, J., T. Maehara, T. Shimokawa, S. Yamamoto and C. Harada et al., 2002. A comprehensive rice transcript map containing 6591 expressed sequence tag sites. Plant Cell, 14: 525-535.
Direct Link - Xu, J.L., H.R Lafitte, Y.M. Gao, B.Y. Fu, R. Torres and Z.K. Li, 2005. QTLs for drought escape and tolerance identified in a set of random introgression lines of rice. Theor. Applied Genet., 111: 1642-1650.
CrossRef - Xu, J., Q. Zhao, P. Du, C. Xu and B. Wang et al., 2010. Developing high throughput genotyped chromosome segment substitution lines based on population whole-genome re-sequencing in rice (Oryza sativa L.). BMC Genomics, 11: 656-656.
CrossRef - Zhu, W., J. Lin, D. Yang, L. Zhao and Y. Zhang et al., 2009. Development of chromosome segment substitution lines derived from backcross between two sequenced rice cultivars, Indica recipient 93-11 and Japonica donor Nipponbare. Plant Mol. Biol. Reporter, 27: 126-131.
CrossRefDirect Link - McCouch, S.R., L. Teytelman, Y. Xu, K. Lobos and K. Clare et al., 2002. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res., 9: 199-207.
CrossRefPubMedDirect Link - Ndjiondjop, M.N., K. Semagn, M. Cissoko, H. Tsunematsu and M. Jones, 2006. Genetic relationships among rice varieties based on expressed sequence tags and microsatellite markers. Asian J. Plant Sci., 5: 429-437.
CrossRefDirect Link - Semagn, K., M.N. Ndjiondjop, M. Loreiux, M. Cissoko, M. Jones and S. McCouch, 2007. Molecular profiling of an interspecific rice population derived from a cross between WAB56-104 (Oryza sativa) and CG 14 (Oryza glaberrima). Afr. J. Biotechnol., 6: 2014-2022.
Direct Link - Pantuwan, G., S. Fukai, M. Cooper, S. Rajatasereekul and J.C. O'Toole, 2002. Yield response of rice (Oryza sativa L.) genotypes to different types of drought under rainfed lowlands: Part 1. Grain yield and yield components. Field Crop Res., 73: 153-168.
CrossRef - Venuprasad, R., M.E. Bool, L. Quiatchon, M.T. Sta Cruz, M. Amante and G.N. Atlin, 2011. A large-effect QTL for rice grain yield under upland drought stress on chromosome 1. Mol. Breed., 30: 535-547.
CrossRef