
Y.T. Gidey
Humera Agricultural Research Center, P.O. Box 62, Humera, Ethiopia
S.A. Kebede
Jimma University, P.O. Box 307, Jimma, Ethiopia
G.T. Gashawbeza
Holetta Agricultural Research Center, P.O. Box 2003, Addis Ababa, Ethiopia
Asian Journal of Agricultural Research
Year: 2012 | Volume: 6 | Issue: 3 | Page No.: 118-128







ABSTRACT
The aim of this study was to characterize and to estimate the extent of genetic variation of morphological attributes of Ethiopian sesame landraces (Sesamum indicum L.). An experiment was conducted in 2010 at Humera and Dansha, Ethiopia. Eighty one germplasm accessions were grown in a lattice design with 2 replications. Data on 17 traits were collected and analyzed. Differences among the accessions were significant for all of the traits at each location and pooled over location. The accessions were grouped into six clusters of different sizes. Cumulative effects of a number of characters dictated differentiation of the accessions into clusters. There was no definite relationship between geographic and genetic diversity as overlapping was encountered in clustering pattern among accessions from different parts of the country. Accessions from different regions might have similar genetic background and those from the same origin might also have different genetic diversity. Therefore, geographic diversity should not necessarily be used as an index of genetic diversity and parental selection should be based on a systematic study of genetic diversity in a specific population. Genetic distances among most of the clusters were significant that crosses between parents selected out of them are expected to generate desirable genetic recombination. Selection should also consider the special advantages of each cluster and each accession within a cluster. Future germplasm collection, conservation and breeding efforts should focus not only on inter-regional diversity but also on intra-regional diversity.
PDF Abstract XML References Citation
Received: December 23, 2011;
Accepted: May 08, 2012;
Published: July 04, 2012
How to cite this article
Y.T. Gidey, S.A. Kebede and G.T. Gashawbeza, 2012. Extent and Pattern of Genetic Diversity for Morpho-agronomic Traits in Ethiopian Sesame Landraces (Sesamum indicum L.). Asian Journal of Agricultural Research, 6: 118-128.
DOI: 10.3923/ajar.2012.118.128
URL: https://scialert.net/abstract/?doi=ajar.2012.118.128
DOI: 10.3923/ajar.2012.118.128
URL: https://scialert.net/abstract/?doi=ajar.2012.118.128
INTRODUCTION
Sesame (Sesamum indicum L.) is diploid specie with 2n = 26 chromosomes. It is self pollinated crop which belongs to order Tubiflorae, family Pedaliaceae. About thirty six species have been described into the genus Sesamum, but only Sesamum indicum has been recognized as a cultivated species (Alemawu et al., 1998). The ancestry, origin and evolution of sesame have been reviewed by Bedigian (1981), Weiss (1983) and Bedigian and Harlan (1986), a variety of cytogenetic, botanical, archaeological, biochemical and historical linguistic criteria have been used.
Two alternative centers of origin have been proposed namely east Africa (Ethiopia) or Asia (Indian sub continent or further east upper central Asia). Sesame could have been distributed either eastward from Africa or westward along the ancient trade routes that are known to have existed (Bedigian, 2004). Sesame is one of the oldest oil seed crops from which oil was extracted by the ancient Hindus which was used for certain ritual purposes (Weiss, 1983). According to Seegeler (1983), it is the first crop recorded in Babylon and Assyria before 732 BC.
Knowledge of the extent and pattern of genetic diversity is very crucial to develop effective genetic resources and in crop evolution, breeding, germplasm collection and conservation strategies (Bartual et al., 1985; Dale et al., 1985; Rezai and Frey, 1990; Jaradat, 1991; Demissie and Bhornstad, 1997). Crosses between parents with maximum genetic divergence would be more responsive to improvement since they are likely to produce higher heterosis and desirable genetic recombination and segregation in their progenies (Arunachalam and Bandyopadhyay, 1984; Reddy, 1988; Singh, 1990; Wallace and Yan, 1998; Chahal and Gosal, 2002). Crosses between parents with high inter-parental diversity may also help to develop varieties with broad genetic base (Russell, 1978; Chandel and Joshi, 1983; Singh, 1990; Keneni et al., 1997) and raises the yield ceilings imposed by a narrow genetic base (Chandel and Joshi, 1983). Landraces have considerable breeding values as they contain valuable adaptive genes to different circumstances (Nechit et al., 1988; Ceccarelli, 1994; Bunder et al., 1996; Chahal and Gosal, 2002).
Even though, it is generally believed that sesame in Ethiopia in general have tremendous genetic diversity, the extent and pattern of genetic diversity in Ethiopian landraces is not yet systematically studied. Geographical separation with physical barriers and genetic barriers to crossability is believed to give rise to genetic diversity among genetic materials (Singh, 1990). However, whether differences in geographic origin (source) necessarily imply genetic distance in parental selection for hybridization is still a matter of some controversy. Joshi and Dhawan (1966) suggested the concept that geographic diversity may serve as an index of genetic diversity in parental selection. Others argue that genetic divergence was not apparently related to geographic diversity in some crops (Prasad et al., 1985; Sindhu, 1985; Nadaf et al., 1986; Rezai and Frey, 1990; Katule et al., 1992). If the former holds true, it is logical to expect that the physical barriers might have resulted in distinct genetic diversity of sesame accessions growing in different parts of Ethiopia as the country of great geographical diversity with high and rugged mountains (EMA, 1988). The objectives of this experiment were, therefore, to estimate the extent and pattern of genetic diversity among the Ethiopian sesame landraces, the relative contribution of various morpho agronomic traits to the total diversity in the accessions and to study the association of geographic origin with genetic diversity.
MATERIALS AND METHODS
Description of the experimental site: The experiment was conducted at two locations, namely, Humera Agricultural Research Center and Dansha, Ethiopia. Humera is located 600 km west of Mekelle, at an altitude of 604 masl and at 14°06'N latitudes and 38°31'E longitudes. Agro-ecology of the region is described as hot to warm semiarid plain sub agro-ecology. The maximum temperature varies from 33°C in May to 42°C in April while minimum temperature varies from 17.5°C in August to 22.2°C in July. Crop production is exclusively dependent on summer rainfall (400-650 mm year-1) which last from June to September. The dominant soil type of Humera Agricultural Research Center is chromic vertisol black in color characterized with very deep (>150 cm) clay textured. Dansha is located at geographical coordinates 25°12'16"N latitude and 15°10'23"E longitudes and at altitude of 948 masl. The mean annual temperature is 28.7°C and it has vertisol soil type. Average annual rainfall varies from 850-1400 mm.
Experimental materials: Eighty one sesame accessions were used for this study. All of the accessions represent the national collections from different major sesame growing regions of Ethiopia and that are maintained at the Institute of Biodiversity Conservation of Ethiopia (IBC). The details of the accessions are given in Table 1.
Experimental material, design and field management: Eighty one random samples of sesame accessions were obtained from Institute of Biodiversity Conservation and Research (IBCR) along with their geographical origins (Table 1). The materials were collected from different agro-ecologies varying in altitude, rainfall and temperature and soil type. The trial was laid out in 9x9 simple lattice designs. Accessions were planted in a plot size of 6.4 m2 (4 rows of 4 m length, 40 cm between rows and 10 cm between plants) in 2010/11 cropping season.
Data collected: Data was collected either on plot basis or from randomly selected 10 plants on days to 50% flowering, days to maturity, capsule filling period, biomass per plot (kg), biomass yield per hectare (kg), seed yield per plot (g), seed yield per hectare (kg), 1000 seed weight (g), harvest index, Oil Content (OC) (%), plant height (cm), number of primary branches per plant, internodes length (cm), height to first capsule (cm), number of capsules per plant, capsule length (cm) and number of seeds per capsule.
Data analysis: The SAS computer package (SAS Institute, 2008) was used to test for presence of outliers and normality of residuals. The total variability for each trait was quantified using pooled analyses of variance over locations using the following model:
Pitjk = μ+lt+ri(t)+bj(i)(t)+gk+(gl)kt+eitjk |
where, pitjk is the phenotypic observation in the ith replication, tth location, jth incomplete block within replication i and location t and from the kth accession, μ is the grand mean, lt is the effect of location t, ri(t) is the effect of replicate i within location t, bj(i)(t) is the effect of the incomplete block j within replication i and location t, gk is the effect of the kth accession, (gl)kt is the effect of interaction of the kth accession and the tth location and eitjk is the random error.
Clustering of accessions was performed by average linkage method of SAS software using seventeen traits that were found to be significantly different among the accessions at least at one of the locations. The pseudo F statistic and the pseudo t2 statistic were examined to decide the numbers of clusters.
| Table 1: | Origin of the eighty one sesame accessions used in the study |
 | |
Genetic distances between clusters as standardized Mahalanobis D2 statistics were calculated as:
D2ij = (xi-xj) S-1(xi-xj) |
where, D2ij is the distance between cases i and j, xi and xj is the vectors of the values of the variables for cases i and j and S-1 is the inverse of pooled variance covariance matrix. Principal components based on correlation matrix were calculated using the same software as in clustering. The D2 values obtained for pairs of clusters were considered as the calculated values of Chi-square (χ2) and were tested for significance both at 1 and 5% probability levels against the tabulated value of χ2 for ‘P’ degree of freedom, where P is the number of characters considered (Singh and Chaudhary, 1985).
RESULTS AND DISCUSSION
Analysis of variance revealed highly significant differences (p<0.01) among the accessions for all characters (Table 2). This indicates that there was adequate variability among the accessions for all of the traits at both locations. Similarly, Alsemaan et al. (2011) reported the existence of genetic diversity within Rosa damascena cultivated in Syria and recommended Almarahl and Bab Alnayrab accessions to be u sed to broaden the production of rose oil. Furthermore, Gichimu and Omondi (2010) reported diversity on five developed Arabica coffee lines as compared to commercial cultivars. The genetic by location interaction for number of capsules per plant, biomass yield, harvest index, number of branches per plant and seed yield showed highly significant difference where as the rest showed non significant (Table 2). This shows there is an interaction effect. The accessions were grouped into six diversity classes (Table 3), different members within a cluster being assumed to be more closely related in terms of the traits under consideration with each other than those members in different clusters.
| Table 2: | Combined analysis of variance (mean squares) for 17 traits of eighty one sesame genotypes |
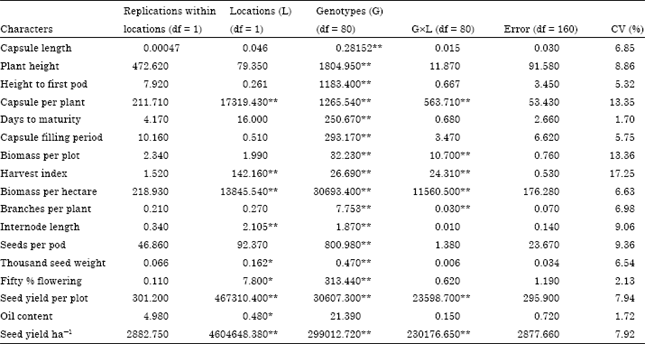 | |
| df: Degree of freedom, *p<0.05 and **p<0.01 are significance level | |
| Table 3: | Grouping of eighty one sesame accessions into different diversity classes |
 | |
Similarly, members in clusters with none significant distance were assumed to have more close relationships with each other than they are with those in significantly distant clusters. Likewise, Rohman et al. (2004) reported that high divergence between the clusters for 25 sorghum varieties which were grouped into four clusters. Cluster C2 was the largest with 42 accessions (51.9%), followed by cluster C1 with 15 accessions (18.5%) of the total population. These clusters constituted intermediate accessions for yielding potential and most of the traits. In addition to this C1 was characterized by shortest plant height, long capsule filling period and smallest biomass. Cluster IV contained 11 accessions (13.6%), while clusters III, V and VI had 8 (9.9%), 4 (4.9%) and 1 (1.2%) accessions, respectively. Cluster VI contained the largest plant height, height to first pod and number of capsules. The average inter cluster D2 values are presented in Table 5. The χ2- test for the six clusters indicated that there was a statistically very highly significant difference in all characters except cluster I with II (17.05), II with III (23.88) and I with IV (10.17). The highest average inter cluster D2 was recorded between cluster IV and cluster V (D2 = 213.83) followed by cluster V and cluster VI (D2 = 188.25) and cluster I and cluster V (D2 = 141.45) which had shown largest inter cluster distance. These clusters were genetically more divergent from each other than any other clusters.
The mean value of the 17 quantitative characters in each cluster is presented in Table 4. Cluster I had the characteristic of smallest plant height (97.91), smallest height to first capsule (29.24), capsule filling period (46.04 days), smallest biomass yield and smallest days to fifty percent flowering (48.21 days).The remaining characters are moderate in magnitude. Cluster II was characterized by smallest capsule length (2.48 cm) and lowest oil content (49.16%). Cluster III was characterized by the following features: largest harvest index (6.98%), fewest numbers of primary branches (3.06) and largest internodes length (4.82 cm). Cluster IV was characterized as the smallest seed yield and with shortest days to maturity (93.05 days). Cluster V was characterized by largest seed yield, largest capsule length (2.81 cm), largest oil content (51.4%) smallest number of capsules per plant (44.25) and smallest thousand seed weight (2.79 g).
| Table 4: | Cluster means for seventeen characters in sesame accessions |
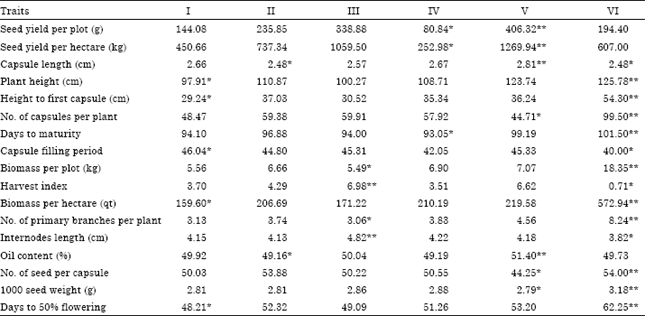 | |
| *,**Indicates for the lowest and the largest value | |
| Table 5: | Pairwise generalized squared distances (D2) among eighty one sesame accessions in six clusters |
 | |
| χ2: 26.30, 32.00 and 39.25 at 5%, 1% and 0.1% probability levels respectively, ***Indicates significant at 0.1% probability levels | |
Cluster VI had a characteristic feature of smallest capsule length (2.48 cm), largest plant height (125.78 cm), largest height to first capsule (54.3 cm), largest number of capsules per plant (99.5) largest days to maturity (101.5 days), smallest capsule filling period (40 days), largest biomass yield, smallest harvest index (0.71%), largest number of primary branches per plant (8.24), smallest internodes length (3.82 cm), largest number of seeds per capsule (54), largest thousand seed weight (3.18 g) and largest days to fifty percent flowering (62.25 days).
According to Ghaderi et al. (1984), increasing parental distance implies a great number of contrasting alleles at the desired loci and then to the extent that these loci recombine in the F2 and F3 generations following a cross of distantly related parents, the greater will be the opportunities for the effective selection for yield factors. Minimum inter cluster distance was observed between cluster I and cluster IV (D2 = 10.17) indicating that genotypes in these clusters were not genetically diverse or there was little genetic diversity between these clusters. This signifies that, crossing of genotypes from these two clusters might not give higher hetrotic value in F1 and narrow range of variability in the segregating F2 population. Maximum genetic recombination is expected from the hybridization of the parents selected from divergent cluster groups. Therefore, maximum recombination and segregation of the progenies is expected from crosses involving parents selected from cluster IV and cluster V, followed by cluster V and VI, despite of that the breeder has to specify his breeding objectives in order to make best use of the characters where they are divergent.
Populations from areas far separated geographically and having complex environment are normally expected to accumulate enormous genetic diversity (Chandel and Joshi, 1983). However, the distribution of strains in different clusters did not follow definite pattern with regard to geographical origins in the present case. Some accessions from different regions were found to be closely related regardless of their geographic origin and the rugged nature of the terrain which could have favored isolation among the accessions and hence, distinct lines of evolution in each region. This could be realized from the overlapping in clustering pattern among accessions from different parts of the country. In most of the cases, accessions from same place of origin fell into the different clusters and from different places of origin fell into same cluster. For instance accessions from around Kemissie area are distributed in different clusters. Results indicate that 8.33% of the accessions are in cluster I, 58.33% in cluster II, 16.67% in cluster III, 8.33% in cluster IV and 8.33% are under cluster V. Accessions from Kobo area are also distributed in different clusters. For example 14.29% of the accessions from Kobo are under cluster I, 57.14% are in cluster II, 14.29% are in cluster IV and 14.29% are in cluster IV. This agrees with the reports of Sudhakar (2003) in which he grouped 62 genotypes into 13 clusters and observed that there is no relation between geographical and genetic diversity. Similarly, Ujjainkar et al. (2002) reported absence of significant relationship between genetic and geographical diversity among 50 evaluated sesame lines on the contrary to Bhatt (1973) suggestion. The fifty sesame germplasm lines evaluated for genetic diversity using 12 characters were grouped into 11 clusters with no significant relationship between genetic and geographically diversity. Furthermore, Belete et al. (2011) reported similar result on 36 Ethiopian mustard accessions. On the other hand, Zhuang et al. (2011) reported that the clustering analysis done on Persian wheat (Triticum turgidum ssp. carthlicum) accessions using EST-SSR markers suggested that most of the accessions with adjacent geographic origins had the tendency to cluster together. Several possible reasons could be given for the genetic similarity among accessions from different regions. There could also be a tendency, particularly among resource-poor farmers in marginal areas, of selecting for the same traits of interest like yield stability, resistance to diseases, insects and abiotic calamities and low dependence on the external inputs (De Boef et al., 1996). Although the original sources might vary, the crop might have also been forced to evolve in the same direction by this kind of local breeding for the same targets which may emanate from similar economic, social, cultural and ecological reasons in the area. Some accessions from the same origin were also found to scatter over different clusters while others were limited to two or three clusters, indicating that genetic diversity in sesame is not uniformly distributed over the regions. Farmers could play important roles in the dynamics of genetic diversity by providing opportunities for hybridization by bringing together geographically and ecologically isolated landraces and selection for desirable agronomic traits (Teshome et al., 1997).
In this study, the results showed that there was high genetic diversity in sesame accessions. Genetic architecture of a population is generally believed to be the result of breeding system, gene flow within and between populations, isolation mechanisms and prolonged selection by various natural and artificial forces (Chandel and Joshi, 1983). Ecological environment is believed to be the major force in crop evolution (Spagnoletti-Zeuli and Qualset, 1987). Therefore, this diversity in sesame accessions could mainly be attributed to diverse agro-climatic conditions in the country as suggested by Harlan (1969). However, there was no definite relationship between geographic diversity and genetic diversity. The concept that geographic diversity could be used as an index of genetic diversity (Joshi and Dhawan, 1966) was also not supported by a number of workers in different crops. It is suggested that selection of parents for hybridization need not necessarily be based on geographic diversity but genetic diversity must form the base for parental selection. Not only interregional diversity but also intraregional diversity should be considered as a useful component in sesame hybridization programs in Ethiopia.
| Table 6: | Percentage and cumulative variances and eigenvectors on the first six principal components for seventeen characters in eighty one sesame accessions |
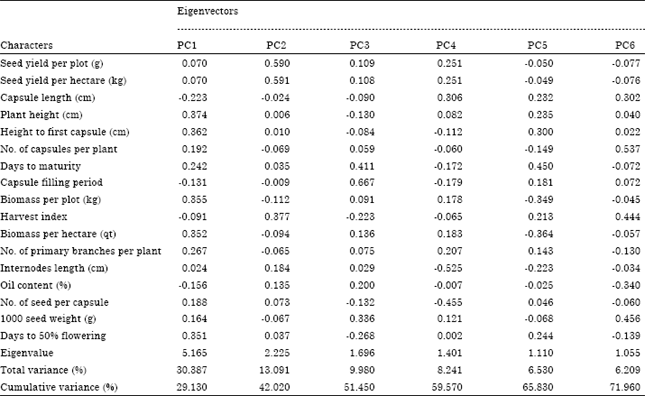 | |
Principal Component Analysis (PCA) is one of the multivariate statistical techniques which are a powerful tool for estimation and summarizing underlying trends in complex data structures (Legendre and Legendre, 1998). Principal component analysis reflects the importance of the largest contributor to the total variation at each axis for differentiation (Sharma, 1998). The principal component for the pooled analysis (Table 6) revealed that six principal components PC1, PC2, PC3, PC4, PC5 and PC6 with eigenvalues 5.16, 2.22, 1.69, 1.40, 1.11 and 1.05, respectively, have accounted for 74.44% of the total variation. The first two principal components PC1 and PC2 with values of 30.39 and 13.09%, respectively contributed more to the total variation. Similarly, Azeez and Morakinyo (2011) reported that the principal component analysis for seed physical dimensions of six sesame accessions revealed that the first two principal components accounted for approximately 88.96% of the total variations among the accessions. Furthermore, Kitila et al. (2011) reported the principal component analysis on 49 coffee accessions revealed the first ten principal components accounts approximately 85.74% of the total variation. On the other side Aragaw et al. (2011) reported that the principle component analysis on 36 ginger accessions revealed that the first six principal components explained 77.7% of the total variation. According to Chahal and Gosal (2002), characters with largest absolute values closer to unity within the first principal component influence the clustering more than those with lower absolute values closer to zero. Therefore, in this study, differentiation of the genotypes into different clusters was because of a cumulative effect of a number of characters rather than the contribution of specific few characters (±0.002-0.667). Agronomic characters having relatively higher value in the first principal component (PC1) were plant height, height to first pod, biomass per plot and fifty percent flowering had more contribution to the total diversity and they were responsible for the differentiation of the six clusters. Characters like seed yield, harvest index, internodes length and oil content had contributed a lot for principal component (PC2), capsule filling period, days to maturity and thousand seed weight had contributed in the third principal component (PC3), internodes length, number of seeds per capsule, capsule length and seed yield per plot and per hectare had contributed in the fourth principal component (PC4), days to maturity, biomass per plot and per hectare, height to first pod and plant height were the major contributors in the fifth principal component (PC5), number of capsules, thousand seed weight, harvest index and capsule length were the major contributor in the last principal component (PC6).
CONCLUSIONS
There is high genetic diversity in the Ethiopian sesame landraces even though it was not uniformly distributed across the regions. Accessions from different regions of origin might be closely related regardless of their geographic origin and accessions from the same regions of origin also might have different genetic background. There was no definite correspondence between geographic origin and genetic diversity. Parental selection should, therefore, not be based on geographic difference but it should rather be made based on systematic assessment of genetic distance in a specific population. Future collection and conservation missions and breeding programs should focus on effective and efficient exploitation of not only inter-regional diversity but also on intra-regional diversity in the species. Inter-cluster gene recombination of sample accessions drawn from the significantly distant clusters followed by selection should prove to generate agronomically desirable progenies as expected.
ACKNOWLEDGMENTS
The authors are thankful to the Institute of Biodiversity Conservation and Research (IBCR) for providing the germplasms used in this study. The Werer Agricultural Research Center is duly acknowledged for its assistance in seed multiplication. The financial, material and technical assistance of the Humera Agricultural Research Center is highly appreciated.
REFERENCES
- Arunachalam, V. and A. Bandyopadhyay, 1984. Limits of genetic divergence for occurrence of heterosis-experimental evidence from crop plants. Indian J. Genet. Pland Breeding, 44: 548-554.
Direct Link - Alsemaan, T., N. Albatal, H. Baydar and K. Almaarri, 2011. Genetic diversity and qualitative variation of Rosa damascene in Syria. Int. J. Agric. Res., 6: 429-436.
CrossRefDirect Link - Azeez, M.A. and J.A. Morakinyo, 2011. Genetic diversity of the seed physical dimensions in cultivated and wild relatives of sesame (Genera Sesamum and Ceratotheca). Int. J. Plant Breed Genet., 5: 369-378.
CrossRef - Bartual, R., E.A. Carbonell and D.E. Green, 1985. Multivariate analysis of a collection of soybean cultivars for southwestern Spain. Euphytica, 34: 113-123.
CrossRef - Bedigian, D., 2004. History and lore of sesame in Southwest Asia. Econ. Bot., 58: 329-353.
CrossRefDirect Link - Bedigian, D. and J.R. Harlan, 1986. Evidence for cultivation of sesame in the ancient world. Econ. Bot., 40: 137-154.
CrossRefDirect Link - Bhatt, G.M., 1973. Comparison of various methods of selecting parents from hybridization in bread wheat (Triticum aestivum L.). Aust. J. Agric. Res., 24: 457-464.
CrossRefDirect Link - Ceccarelli, S., 1994. Specific adaptation and breeding for marginal conditions. Euphytica, 77: 205-219.
CrossRef - Chandel K.P.S. and B.S. Joshi, 1983. Multivariate analysis in green-seeded pea. Indian J. Agric. Sci., 53: 198-200.
Direct Link - Dale, M.F.B., B.V. Ford-Lloyd and M.H. Arnold, 1985. Variation in some agronomically important characters in a germplasm collection of beet (Beta vulgaris L.). Euphytica, 34: 449-455.
CrossRef - Demissie, A. and A. Bhornstad, 1997. Geographical, altitude and agro-ecological differentiation of isozyme and hordein genotypes of landrace barleys from Ethiopia: Implications to germplasm conservation. Genet. Resour. Crop Evol., 44: 43-55.
CrossRefDirect Link - Prasad M.M.K.D., V. Arunachachalam and A. Bandyopadyay, 1985. Diversity pattern elucidating choice of parents for hybridization in varieties of groundnut, Arachis hypogaea L. Trop. Agric., 62: 237-242.
Direct Link - Ghaderi, A., M.W. Adams and A.M. Nassib, 1984. Relationship between genetic distance and heterosis for yield and morphological traits in dry edible bean and faba bean. Crop Sci., 24: 37-42.
CrossRefDirect Link - Gichimu, B.M. and C.O. Omondi, 2010. Morphological characterization of five newly developed lines of Arabica coffee as compared to commercial cultivars in Kenya. Int. J. Plant Breed. Genet., 4: 238-246.
CrossRefDirect Link - Jaradat, A.A., 1991. Phenotypic divergence for morphological and yield-related traits among landrace genotypes of durum wheat from Jordan. Euphytica, 52: 155-164.
CrossRefDirect Link - Aragaw, M., S. Alamerew, G.H. Michael and A. Tesfaye, 2011. Variability of ginger (Zingiber officinale Rosc.) accessions for morphological and some quality traits in Ethiopia. Int. J. Agric. Res., 6: 444-457.
CrossRefDirect Link - Kitila, O., S. Alamerew, T. Kufa and W. Garedew, 2011. Genetic diversity analysis of limmu coffee (Coffea arabica L.) collection using quantitative traits in Ethiopia. Int. J. Agric. Res., 6: 470-481.
CrossRef - Rezai, A. and K.J. Frey, 1990. Multivariate analysis of variation among wild oat accessions-seed traits. Euphytica, 49: 111-119.
CrossRef - Rohman, M.M., M.A. Hakim, N.A. Sultana, M.E. Kabir, M. Hasanuzzaman and M. Ali, 2004. Genetic divergence analysis in sorghum (Sorghum bicolor L.). Asian J. Plant Sci., 3: 211-214.
CrossRefDirect Link - Teshome, A., B.R. Baum, L. Fahrig, J.K. Torrance, T.J. Arnason and J.D. Lambert, 1997. Sorghum [ Sorghum bicolor ( L.) moench ] landrace variation and classification in North Shewa and South Welo, Ethiopia. Euphytica, 97: 255-263.
Direct Link - Belete, Y.S., S.A. Kebede and A.W. Gemelal, 2011. Multivariate analysis of genetic divergence among Ethiopian mustard (Brassica carinata A. Brun) genotypes in relation to seed oil quality traits. Int. J. Agric. Res., 6: 494-503.
CrossRef - Zhuang, P.P., Q.C. Ren, W. Li and G.Y. Chen, 2011. Genetic diversity of Persian wheat (Triticum turgidum ssp. carthlicum) accessions by EST-SSR markers. Am. J. Biochem. Mol. Biol., 1: 223-230.
CrossRefDirect Link