
J.P. Gweyi-Onyango
Department of Agricultural Sciences, Kenyatta University, P.O. Box 43844-00100 Nairobi, Kenya
LiveDNA: 254.233
T. Tesfamariam
Institute of Plant nutrition (330), University of Hohenheim, 70593 Stuttgart, Germany
G. Neumann
Institute of Plant nutrition (330), University of Hohenheim, 70593 Stuttgart, Germany
Asian Journal of Agricultural Research
Year: 2011 | Volume: 5 | Issue: 1 | Page No.: 45-55







ABSTRACT
Tropical soils are characterized by low pH with high aluminium saturation and phosphorus (p) fixation. Pinto Peanut (Arachis pintoi (Krapov and W.C. Gregory)) grows in such soils with no P deficiency symptoms. The current experiment dissected an array of mechanisms envisaged to explain growth advantages of Arachis pintoi (AP) in such P-deficient environments. Split root experiment was conducted with three ecotypes (CIAT 17434, CIAT 18744 and CIAT 22172) cultured in hydroponics with Hoagland solutions and P was later withheld from one root compartment. Nutrient solution pH was determined from 20-49 Days after sowing using pH meter, while exudates were collected by overlaying filter papers on root surfaces. Carboxylates were extracted from filter papers and later determined by HPLC (Bio-Rad, Richmond, CA, USA). CIAT 18744 produced significantly higher (p≤0.05) shoot and root (biomass), whereas CIAT 22172 had lowest. CIAT 18744 registered minimal acidification under P-deficiency compared to other two ecotypes and produced highest TCA cycle carboxylates (1.72 and 2.2 nmol/h/cm root length) malate and citrate, respectively whereas CIAT 22172, exuded only 0.50 and 0.52 nmol/h/cm root length] malate and citrate, respectively. There was no correlation between growth medium acidification and carboxylate exudation; probably implying carboxylates lacked contribution to growth medium acidification. Selecting for carboxylate exudation and internal P reallocation would be a better strategy in acid soils (e.g., Kenyan Alfisols) since carboxylates can complex aluminium without acidification of rhizosphere, while protonation that leads to rhizosphere acidification would be a better strategy in high pH soils, favoring availability of precipitated P (e.g., Calcareous Kenyan soils).
PDF Abstract XML References Citation
Received: July 07, 2010;
Accepted: August 20, 2010;
Published: October 11, 2010
How to cite this article
J.P. Gweyi-Onyango, T. Tesfamariam and G. Neumann, 2011. Contrasting Responses to Phosphorus Status by Arachis pintoi (Krapov and W.C. Gregory): A Lesson for Selecting Vegetables for Cultivation in Kenyan Ecozones. Asian Journal of Agricultural Research, 5: 45-55.
DOI: 10.3923/ajar.2011.45.55
URL: https://scialert.net/abstract/?doi=ajar.2011.45.55
DOI: 10.3923/ajar.2011.45.55
URL: https://scialert.net/abstract/?doi=ajar.2011.45.55
INTRODUCTION
Major factors that constrain fertility of tropical soils and sustainable agriculture are low nutrient capital, moisture stress, erosion, high acidity with aluminium toxicity and high P fixation (Cardoso and Kuyper, 2006). In these soils, P is one of the least available nutrients, often at concentrations similar to concentrations of micronutrients (Marschner, 1995; Raghothama, 1999). Most of the P supplied to the soils to meet P demand of plants is converted into unavailable forms that can not be easily used and taken up by plant roots. According to Vance et al. (2003), plants have evolved two broad strategies for P acquisition and use in nutrient-limiting environments: (1) Those aimed at conservation of use and (2) those directed towards enhanced acquisition or uptake. A diverse array of plant mechanisms have evolved to obtain adequate phosphorus under limiting conditions, including mycorrhizal symbiosis, rhizosphere modification with organic acids, protons, enzymes and chelators (Lynch and Ho, 2005; Raghothama, 1999). Other factors increase efficient metabolic use of phosphorus within the plant, such as increased growth rate per unit of phosphorus uptake and alternative respiratory pathways.
The roots of certain crop species, including white lupin (Lupinus albus L.) and chickpea (Cicer arietinum L.) exude large amounts of low-molecular weight organic anions (carboxylates) which enhance the availability of soil phosphorus to the plant (Ae et al., 1990; Dinkelaker et al., 1989; Gardner et al., 1983; Gerke et al., 1994; Hocking et al., 1997; Ohwaki and Hirata, 1992). Organic anions can complex metal cations, which bind phosphates (Al3+, Fe3+, Ca2+), or displace phosphate from the soil matrix by ligand exchange (Jones, 1998).
Where carboxylate exudation is associated with proton extrusion, the lower pH may itself contribute to greater P availability, if the soil pH is relatively high. However, cotransport of protons is certainly not associated with organic anion release in all species under all circumstances (Roelofs et al., 2001; Ryan et al., 2001). The effectiveness of carboxylates depends on their number of carboxyl groups and molecular structure. Tricarboxylates (e.g., citrate) are generally more effective than dicarboxylates (e.g., malate, malonate) due to stronger ligand binding. Soil properties also have large effects on the effectivity of carboxylates. For example, the stability of organic anion – metal complexes depends strongly on soil pH (Jones, 1998) and there is large variation in the mechanisms and strength with which phosphate is held in the soil. For this reason root exudation patterns may need to be different on different soils for optimal effects on P (mobilization). It is known that species differ in the carboxylates they exude, e.g., predominantly citrate in white lupin, malonate in chickpea, malate in wheat and a wide range of carboxylates in Proteaceae species (Roelofs et al., 2001; Ryan et al., 2001), but it is not clear if that affects the suitability of these species for certain soils or, more specifically, their ability to mobilize soil P. Knowledge about variation in carboxylate exudation in response to soil P status is virtually limited to observations that lupin root exudation rates are highest at low P availability (Keerthisinghe et al., 1998). However, Lambers et al. (2002) found that Bull Banksia (Banksia grandis Willd) an Australian Proteaceae species, modified its exudation pattern when grown on sand containing either poorly soluble Al-phosphate or Fe-phosphate as the only P source and grew equally well on either P source. Studies of Al3+ resistance have shown clearly that plants respond directly to the presence of Al3+ by releasing organic anions and those genotypes of wheat (Triticum aestivum L.), maize (Zea mays L.) and soybean (Glycine max L. Merr.) vary enormously in this trait (Ryan et al., 2001). If such genetic variation were associated with the ability to respond to P deficiency by producing the appropriate root exudates to enhance P availability, this could be exploited to improve the efficiency of P fertilizer use. Most tropical and subtropical soils generally have very low levels of P and much P fertilizer is ineffective due to high P sorption capacities (Bolland et al., 1994; Brennan et al., 1994). Although, iron oxides are often present in high concentrations, the sorption of P is most closely associated with aluminium (oxides and organic complexes) (Gilkes and Hughes, 1994). We herein report the results of hydroponics experiments, in which we determined plant growth and root exudations in terms of carboxylates (citrate, malate, fumarate and c-aconitate) and proton excretions (nutrient pH changes). The focus was on P deficiency-induced variation in root exudates among different accessions of Arachis pintio plants with known yield potentials. Present previous soil experiments failed to explain yield differences between CIAT 17434 and CIAT 18744 on basis of carboxylates excretion and we envisaged less interactions and interferences in nutrient solutions and hence examined array of carboxylic acid, including citrate from (tricarboxylic acid cycle) and Malate (from dicarboxylic acid cycle). Split root approach was aimed at testing the P tissue reallocation potential of the different AP accessions as a strategy against P stress.
MATERIALS AND METHODS
Plant cultivation in hydroponics: The experiment was conducted in nutrient solution in Growth chamber at Institute of Plant Nutrition; Universitaet Hohenheim, Stuttgart, Germany between September 2005 and February 2006. Obtained pinto peanut (Arachis pintoi (Krapov. and W.C. Gregory)) seeds were soaked for 30 min in aerated 10 mM CaSO4 solution and then geminated in rolls of filter papers soaked with 2.5 mM CaSO4 in the dark at 25°C. After 5 days, pre-germinated seeds were transferred to light for one day and then cultivated in pots containing 2.5 L of aerated half-strength nutrient solution (10 plants per pot) in a growth chamber with a 16/8 h light/dark regime at 25°C with light intensity of 150 μmol m-2 sec-1 and a relative humidity of 60% (Gweyi-Onyango, 2006). After 5 days the concentration of nutrients was doubled to full strength. (Composition: 2 mM Ca(NO3)2; 0.1-0.25 mM KH2PO4, 0.7 mM K2SO4; 0.1 mM KCl; 0.5 mM MgSO4; 30 μM Fe-EDTA; 10 μM H3BO3; 0.5 μM MnSO4; 0.5 μM ZnSO4; 0.2 μM CuSO4; 0.01 μM (NH4)6Mo7O24). After a pre-culture period of four weeks with complete nutrient solution, three Arachis pintoi ecotypes (CIAT 17434, CIAT 18744 and CIAT 22172) were grown for 47 days without P supply or with soluble P (0.25 mM KH2PO4) on different split root compartments.
High-Performance Liquid Chromatography (HPLC) analysis: Localized collection of root exudates for different AP genotypes were done on fresh roots; small pieces of wet filter paper (Kania, 2005), previously washed with methanol and double distilled water were placed on the roots of plant samples and covered with moistened filter papers to prevent the plant roots from drying out. After 2 h, the filter papers used for collecting the samples were removed from the corresponding roots and put into Eppendorf vials and stored deep freezer. Extractions of root exudates from the filter papers were extracted from filter papers by centrifugation (5 min at 20,000x g) and the supernatant was used for HPLC analysis after appropriate dilution with the HPLC mobile phase (Neumann, 2006). Separation of organic acids was conducted on an Aminex 87H ion exclusion column (Bio-Rad, Richmond, CA, USA). A sample volume of 20 μL was injected into the isocratic flow (0.5 mL min-1) of the eluent (2.5 mM H2SO4, 40°C) and organic acids were detected spectrophotometrically at 210 nm (Neumann, 2006). Identification and quantification of carboxylates were conducted by comparing the retention times, absorption spectra and peak areas with known standards.
Plant harvest and pH determination: Nutrient solution pH was monitored using pH meter on consecutive days and plants were harvested at the end of culture period (47 days after sowing-DAS). Shoot and root parts were separated and gently pressed between sorption papers to remove adhering water and weight determined with weighing balance.
Statistical analysis: Data were analysed with a statistical software Sigma Stat® (Jandel Scientific). Differences between the treatments were performed with a one-way ANOVA and those showing differences further subjected Turkey test (p≤0.05) and significant differences were indicated with different letters.
RESULTS
Varietal differences in biomass production: The results showed clear variation in shoot and root fresh weights; with CIAT 18744 showing significantly higher shoot fresh weights while CIAT 22172 had the lowest shoot fresh weight (Table 1). There was an observed similar trend with root weights. The differences between the higher yielding ecotypes (CIAT 18744) and CIAT 17434 were however not statistically significant in terms of root fresh weights. The highest yielding ecotypes (CIAT 18744) showed no superiority in terms of root: shoot ratio (Table 1). The differences between root fresh weights were 26 and 54 % higher in case of CIAT 18744 as compared to CIAT 17434 and CIAT 22172, respectively, whereas the difference in shoot fresh weights for CIAT 17434 and 22172 were 18 and 40%, respectively lower as compared to CIAT 18744. However, root to shoot ratio of the higher yielding CIAT 18744 was lower than CIAT 17434 but higher than CIAT 22172.
Nutrient solution pH variation with progression of time of P deprivation: P supply elicited differential growth medium pH responses among different accessions studied. The results revealed differences between P-treated as compared with case where it was omitted (Fig. 1a-c). The differences between the nutrient solution pH values varied between 1-2.8 units between the two P treatments (minus or plus P).
| Table 1: | Variations in shoot, root fresh weight and root to shoot ration of different Arachis pintoi ecotypes cultured in nutrient solutions. Plant were harvested 39 DAS; n = 4 |
 | |
| Values followed with similar letter(s) along the column are not statistically significant at p≤0.05 | |
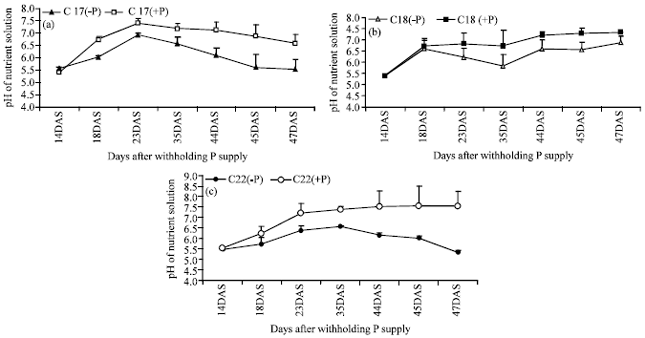 | |
| Fig. 1: | Nutrient solution pH changes of different Arachis pintoi ecotypes as affected by phosphorus supply-either plus P or minus P (C17 = CIAT 17434, C18 = CIAT 18744 and C22 = CIAT 22172) |
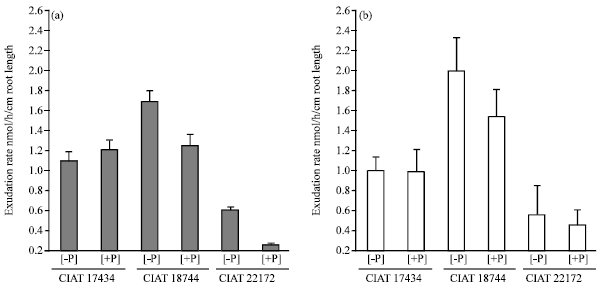 | |
| Fig. 2: | Variation in carboxylate exudation rates among Arachis pintoi ecotypes grown in hydroponic culture in a split root experiment with half part of roots without P (-P) and the other half compartment supplied with P (+P). Exudation rate was determined 39 DAS. n = 4. (a) malate and (b) citrate |
As the days progressed from time of withholding P supply from the nutrient solution, variations in pH values became more pronounced between the two P treatments and these observations were particularly evident in cases of CIAT 22172 and CIAT 17434, while the highest yielding accession (CIAT 18744) exhibited comparatively lower acidification of nutrient solution under P deficiency (suggesting lower proton excretion potential). The results revealed a distinct increase in nutrient solution pH during initial stages of growth (14-23 DAS) irrespective of P supply; after which trends changed (pH for minus P dropped in CIAT 17434 and CIAT 22172). At 47 DAS, CIAT 22172 had the highest pH differences between P treatments and this was followed by CIAT 17434, whereas the better performing accession (CIAT 18744) showed marginal differences between the P treatments (both P treatments had pH values of about 7.0 at 47 DAS) (Fig. 1).
P supply effect on carboxylates exudation by different Arachis pintoi ecotypes: Exudation of carboxylates showed variability among the studied three Arachis pintoi ecotypes. Both CIAT 18744 and CIAT 22172 showed significantly higher excretion of malate per unit root length under P starvation. When P was omitted from nutrient solution, malate exudation rate was (1.8 and 0.6 nmol/h/cm root length) for CIAT 18744 and CIAT 22172, respectively) whereas under sufficient P supply, it was only [1.3 and 0.2 (nmol h/cm/root length)] for CIAT 18744 and CIAT 22172, respectively (Fig. 2a, b).
On the contrary, CIAT 17434 showed higher malate exudation with the supply of P. The differences in malate exudation between plus and minus P-treatments were, however, statistically insignificant in case of CIAT 17434 (Fig. 2). The trends in citrate exudation patterns by Arachis pintoi ecotypes were similar to malate exudations, though the differences between P treatments for each ecotype were not statistically significant (p≤0.05).
It was interesting to note that fumarate exudation rate was enhanced by P supply as compared to case when phosphorus was omitted in the growth medium. The differences among the studied accessions in terms of fumarate exudation were not significant (p≤0.05).
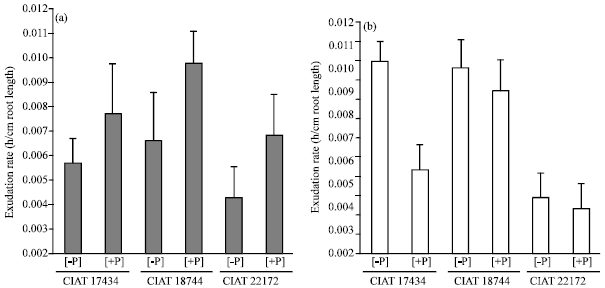 | |
| Fig. 3: | Differential carboxylate exudation rates among three Arachis pintoi ecotypes cultured in hydroponics supplied with P (+P) on one compartment of the split root while other compartment received no P (-P). Exudation rate was done on 39 DAS, n = 4. (a) fumarate and (b) c-aconitate |
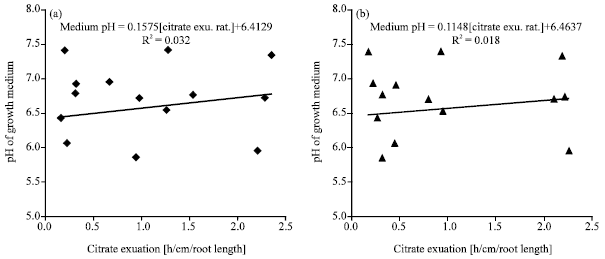 | |
| Fig. 4: | Relationships between Arachis pintoi ecotypes citrate (a) and malate (b) exudation rates and nutrient solution pH changes- at 35 DAS (Days after sowing); n = 4 ; p≤0.05 |
Similar trends were observed with C-aconitate exudation (Fig. 3a, b). However, there was an exception with CIAT 17434, where there was an enhanced C-aconitate exudation when P was omitted from growth medium as compared to sufficient P supply. The results revealed a similar level of C-aconitate exudation between CIAT 1734 and CIAT 18744 when compared under P starvation condition. However, under similar P-sufficient treatment, CIAT 18744 expressed more C-aconitate exudation. Notably, CIAT 22172 expressed the lowest expression of C- aconitate per unit root length in a given time irrespective of P supply status. This trend was similar for malate and citrate exudation with this accession (CIAT 22172) (Fig. 2, 3).
The correlations between citrate exudation rate and nutrient pH (acidification) were weak (R2 = 0.03) at 35 DAS. This lack of correlation between nutrient pH was also observed with malate (R2 = 0.02) at 35 DAS (Fig. 4a, b). The lack of correlation of nutrient pH and the TCA cycle (citrate) and dicarboxylic acid (malate) implies that the acidification under P stress (Fig. 1) may not be a result of carboxylic acid excreted by the roots into growth medium and probably solely due to protonation (H+) of the nutrient solution (Fig. 1, 2 and 4).
DISCUSSION
At optimum phosphorus supply, CIAT 17434 and CIAT 18744 were both observed to have similar shoot yields of 17 g plant-1 from previous experiments (Claasen et al., 2005). But at limited P supply, the P efficient genotype (CIAT 18744) out-yielded the P-inefficient genotype (CIAT 17434) by a factor of about four. The biomass yield superiority of P-efficient (CIAT 18744) in terms of shoot and root production as compared with two accessions (CIAT 17434 and CIAT 22172) (Table 1) are consistent with reports of other workers (Claassen et al., 2005; Marschner et al., 2007). The biomass production (shoot fresh weight) superiority was however only between 26 and 54% for CIAT 18744 as compared to CIAT 17434 and CIAT 22172, respectively. These observations (Claasen et al., 2005; Marschner et al., 2007) also showed marginal differences in shoot biomass between the genotypes during earlier stages of growth that became conspicuous with progression of growth. In the current experiment there was gradual P limitation with duration of culture period since one part of the root compartment was P-starved; though both roots (together as one) and the whole shoots were harvested (Table 1). Previous experiments failed to demonstrate higher P uptake by more root per unit shoot (Claasen et al., 2005) and other results (Fohse et al., 1988; Marschner et al., 2007) demonstrated low P content per plant for efficient genotypes. Current experiment also observed a lower root to shoot ratio for the efficient Arachis pintoi (Ap) accession (CIAT 18744) as compared to CIAT 17434 genotype (Table 1), a sign that P uptake may not be the main contributing factor to accession growth superiority. However, Claasen et al. (2005) attributed the yield differences to higher P influx in case of CIAT 18744 genotype as one of the factors contributing to its P efficiency.
Differences in proton excretions have previously been reported between genotypes of tomatoes (Gweyi-Onyango, 2006). The P efficiency amongst the tomato genotypes could not, however, be attributed to protonation potential of the genotypes. In the current experiment, the efficient genotype showed less variation in nutrient solution pH between P treatments (Fig. 1). This implies that in such genotype, rhizosphere pH acidification is not a strategy for P acquisition. A possible explanation of lack of intensive root proton excretion may be due to enhanced retranslocation potential of CIAT 18744 from the P sufficient plant organs and/or tissues to roots cultured in minus P compartment. Such remobilization of P has been previously reported in Caribbean stylo (Stylosanthes hamata L. Taub.) and tomatoes (Gweyi-Onyango 2006; Smith et al., 1990). However, CIAT 18744’s resistance to acidification could also be a protective strategy against root injury, especially to avoid aluminum toxicity, which is always associated with P deficiency. This ability to resist acidification (CIAT 18744) is in line with reports of Cowpea (Vigna unguiculata L. Walp.) genotypes exposed to low soil pH (Akinrinde et al., 2006). On the contrary, rhizosphere acidification, as depicted by CIAT 17434 and CIAT 22172 may be a better P acquisition strategy, particularly under high soil pH. These observed enhanced acidifications under P stress are in agreement with results of Gahoonia et al. (2002).
Previous experiment (Claasen et al., 2005) examined carboxylate exudation patterns between two genotypes of Ap and the only measurable carboxylates determined were Oxaloacetic Acid (OA), lactic acid and acetic acids. There were however no correlation between lactic acid and acetic acid with soil P content at the rhizosphere for CIAT 18744 and CIAT 17434. Indeed the P-inefficient genotype had very high lactic acid exudation as compared to P-efficient genotype and the lactic acid was negatively correlated with soil P, which was more pronounced with age of the plants. The above exudates could therefore not explain the varietal yield differences of the studied pinto peanut. Related work (Otani et al., 1996) also found for the groundnut or peanut (Arachis hypogea L.) that OA was not responsible for P-efficiency and in a later paper (Ae and Otani, 1997); it was proposed that cell wall component was related to the P efficiency of plants. The results herein, also found higher OA as reported previously (Ae and Otani, 1997; Otani et al., 1996) (results not shown) and also lactic acid (results not shown) as reported by Claasen et al. (2005). Interestingly, current experiment revealed higher exudation for TCA cycle carboxylate (citrate) and dicarboxylic acid cycle (malate) in case of CIAT 18744 as compared to CIAT 17434 and CIAT 22172 under P deficiency conditions (Fig. 3). There was however no clear root exudation patterns in terms of less efficient carboxylates (fumerate and C-aconitate) amongst the three studied Ap accessions (Fig. 4). The enhanced exudation of TCA carboxylate would therefore partly explain the P efficiency of CIAT 18744 genotype. Mobilisation of P is greatest for citrate (tricarboxylic acid) followed by oxalate (dicarboxylic acid), with malate, malonate and tartarate being moderately effective and succinate, fumarate, acetate and lactate being the least effective (Nagarajah et al., 1970; Richarson et al., 2009). Indeed it is unlikely that acetate and lactate were direct exudates from the roots, but rather, products of microbial decomposition of exudates (Saarnio et al., 2004) and hence not efficient in P mobilization from soil.
The acidification of the nutrient solution (protonation) had poor correlation with carboxylic acid (Citrate and Malate) (Fig. 4). This probably implies that acidification was not related to carboxylic acids exudation into nutrient solutions and most likely proton extrusion and carboxylate exudation are regulated by independent biochemical pathways. The current results are at variance with previous conclusions that decrease in rhizosphere pH was due to the accompanying H+ extrusion (Dinkelaker et al., 1989). However, present results are in support of findings that demonstrated that the release of organic acid across plasma membrane involved two separate transport processes because of high cytosolic pH (Yan et al., 2002). Firstly, citrate transport is mediated by anion channel; and this is also supported by findings of Neumann et al. (1999) in which the release of citrate from proteoid roots of white lupin was mediated by anion channels. And secondly, the export of H+ (CIAT 17434 and CIAT 22172 scenario) from the plant cell is mediated by the plasma membrane H+-ATPase (Serrano, 1989; Yan et al., 2002). Therefore, these apparent differences in pathways employed by different AP accessions in response to P stress can be a big lesson in selecting for different plant strategies for P adaptation in various ecosystems, for example in the growth of vegetables in Kenya.
CONCLUSION AND RECOMMENDATIONS
The current experiment points to a number of strategies employed by Arachis pintoi ecotypes in alleviating P deficiency. Internal P mobilization and remobilization as well as citrate exudation was key strategy by CIAT 18744 and this may be a good strategy for selecting field crops and vegetables for cultivation in acid Alfisols of Western Kenya. This is because the carboxylates can mobilize P and chelate Al without further decreasing rhizosphere pH. The proton release (Strategy employed by CIAT 17434 and CIAT 22172) under acidic situation can only aggravate the situation by increasing saturation of toxic aluminum species. On the contrary, protonation of rhizosphere as exhibited by these accessions (CIAT 17434 and CIAT 22172) would be an alternative strategy for selecting crops for soils in Semi- arid regions of Eastern Kenya that are characterized by high pH. This is because the protons would make the P available from Ca-P. Therefore, breeding for P efficiency should target genes responsible for carboxylic acid, particularly citrate exudations as well as TQL loci for H+-ATPase over-expression and proton excretion for crops targeting alkaline Luvisol in areas like Kibwezi in Eastern province of Kenya.
ACKNOWLEDGMENT
We wish to thank DAAD for financial support without which this experiment would not have been possible. We also thank Dr. I M Rao for Arachis pintoi germplasm. Our sincere thanks are also due for Dr A. Kania for helping with HPLC analysis. Thanks also go to Institute of Plant Nutrition, Universitaet Hohenheim, Stuttgart (Germany) for logistic support.
REFERENCES
- Ae, N. and T. Otani, 1997. The role of cell wall components from groundnut roots in solubilizing sparingly soluble phosphorus in low fertility soils. Plant. Soil., 196: 265-270.
CrossRef - Ae, N., J. Arihara, K. Okada, T. Yoshihara and C. Johansen, 1990. Phosphorus uptake by pigeon pea and its role in cropping systems of the Indian subcontinent. Science, 248: 477-480.
CrossRefDirect Link - Bolland, M.D.A., I.R. Wilson and D.G. Allen, 1994. Effect of P buffer capacity and P retention index of soils on soil test P, soil test P calibrations and yield response curvature. Aust. J. Soil Res., 32: 503-517.
CrossRef - Claasen, N., N. Castaneda-Ortiz, G. Neumann, J.P. Gweyi-Onyango and I.M. Rao, 2005. Root Exudation of Organic Acids and/or Acid Phosphatase Cannot Explain Phosphorus Mobilization by Arachis pintoi. In: Plant Nutrition for Food Security, Human Health and Environmental Protection, Li, C.J. (Ed.). Tsingua University Press, Hong Kong, China, pp: 460-461.
- Dinkelaker, B., V. Romheld and H. Marschner, 1989. Citric acid excretion and precipitation of calcium citrate in the rhizosphere of white lupin (Lupinus albus L.). Plant Cell Environ., 12: 285-292.
CrossRef - Gahoonia, T.S., N. Claassen and A. Jungk, 1992. Mobilization of phosphate in different soils by ryegrass supplied with ammonium or nitrate. Plant Soil, 140: 241-248.
CrossRefDirect Link - Gerke, J., W. Romer and A. Jungk, 1994. The excretion of citric and malic acid by proteoid roots of Lupinus albus L; effect on soil solution concentrations of phosphate, iron and aluminum in the proteoid rhizosphere in samples of an oxisol and a luvisol. Zeitschrift Pflanzenernahrung Bodenkunde, 157: 289-294.
CrossRef - Gilkes, R.J. and J.C. Hughes, 1994. Sodium fluoride pH of southwestern Australian soils as an indicator of P-sorption. Aust. J. Soil Res., 32: 755-766.
CrossRef - Hocking, P.J, G. Keerthisinghe, F.W. Smith and P.J. Randall, 1997. Comparison of the Ability of Different Crop Species to Access Poorly-Available Soil Phosphorus. In: Plant Nutrition for Sustainable Food Production and Environment, Ando, T., K. Fujita, T. Mae, H. Matsumoto, S. Mori and J. Sekiya (Eds.). Kluwer Academic Publishers, Dordrecht, pp: 305-308.
- Keerthisinghe, G., P.J. Hocking, P.R. Ryan and E. Delhaize, 1998. Effect of phosphorus supply on the formation of proteoid roots of white lupin (Lupinus albus L.). Plant Cell. Environ., 21: 467-478.
Direct Link - Lambers, H., D. Juniper, G.R. Cawthray, E.J. Veneklaas and E. Martinez-Ferri, 2002. The pattern of carboxylate exudation in Banksia grandis (Proteaceae) is affected by the form of phosphate added to the soil. Plant Soil., 238: 111-122.
CrossRef - Lynch, J.P. and M.D. Ho, 2005. Rhizoeconomics: Carbon costs of phosphorus acquisition. Plant Soil., 269: 45-56.
CrossRefDirect Link - Marschner, H., 1995. Mineral Nutrition of Higher Plants. 2nd Edn., Elsevier, Amsterdam, Netherlands, ISBN-13: 9780080571874, Pages: 889.
Direct Link - Marschner, P., Z. Solaiman and Z. Rengel, 2007. Brassica genotypes differ in growth, phosphorus uptake and rhizosphere properties under P-limiting conditions. Soil Biol. Biochem., 39: 87-98.
CrossRef - Nagarajah, S., A.M. Posner and J.P. Quirk, 1970. Competitive adsorption of phosphate with polygalacturonate and other organic anions on kaolinite and oxide surfaces. Nature, 228: 83-85.
CrossRef - Ohwaki, Y. and H. Hirata, 1992. Differences in carboxylic acid exudation among P-starved leguminous crops in relation to carboxylic acid contents in plant tissues and phospholipid level in roots. Soil Sci. Plant Nutr., 38: 235-243.
CrossRef - Raghothama, K.G., 1999. Phosphate acquisition. Ann. Ver. Plant physiol. Plant Mol. Biol., 50: 665-693.
PubMed - Richarson, A.E., P.J. Hocking, R.J. Simpson and T.S. George, 2009. Plant mechanisms to optimize access to soil phosphorus. Crop Pasture Sci., 60: 124-143.
CrossRef - Roelofs, R.F.R., Z. Rengel, G.R. Cawthray, K.W. Dixon and H. Lambers, 2001. Exudation of carboxylates in Australian Proteaceae: Chemical composition. Plant Cell Environ., 24: 891-904.
Direct Link - Ryan, P.R. and E. Delhaize, 2001. Function and mechanism of organic anion exudation from plant roots. Annu. Rev. Plant Physiol. Plant Mol. Biol., 52: 527-560.
CrossRefDirect Link - Saarnio, S., L. Wittenmayer and W. Merbach, 2004. Rhizospheric exudation of Eriophorum viginatum L.-potential link to methanoganesis. Plant Soil, 267: 343-355.
CrossRef - Serrano, R., 1989. Structure and function of plasma membrane ATPase. Ann. Rev. Plant Pathol. Mol. Biol., 40: 61-94.
CrossRef - Smith, F.W., W.A. Jackson and P.J. Vanden Berg, 1990. Internal phosphorus flows during development of phosphorus stress in Stylosanthes hamata. Aust. J. Plant Physiol., 17: 451-464.
CrossRef - Vance, C.P., C. Uhde-Stone and D.L. Allan, 2003. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol., 157: 423-447.
CrossRefDirect Link - Yan, F., Y. Zhu, C. Muller, C. Zorb and S. Schubbert, 2002. Adaptation of H+-ATPase activity in proteoid roots of white lupin under phosphate deficiency. Plant Physiol., 129: 50-63.
PubMed - Fohse, D., N. Claassen and A. Jungk, 1988. Phosphorus efficiency in plants. I. External and internal P requirement and P uptake efficiency of different plant species. Plant Soil, 110: 101-109.
CrossRef - Cardoso, I.M. and T.W. Kuyper, 2006. Mycorrhizae and tropical soil fertility. Agric. Ecosyst. Environ., 116: 72-84.
CrossRef - Akinrinde, E.A., L. Iroh, G.O. Obigbesan, T. Hilger, G. Neumann and V. Romheld, 2006. Differential expression of aluminium tolerance mechanisms in cowpea genotypes under phosphorus limitation. J. Applied Sci., 6: 854-859.
CrossRefDirect Link - Jones, D.L., 1998. Organic acids in the rhizosphere: A critical review. Plant Soil, 205: 25-44.
CrossRef - Brennan, R.F., M.D.A. Bolland, R.C. Jeffery and D.G. Allen, 1994. Phosphorus adsorption by a range of Western-Australian soils related to soil properties. Commun. Soil Sci. Plant Anal., 25: 2785-2795.
CrossRefDirect Link