
V. Atlassi Pak
Department of Agronomy and Plant Breeding, Shahid Chamran University, Ahvaz, Iran
M. Nabipour
Department of Agronomy and Plant Breeding, Shahid Chamran University, Ahvaz, Iran
M. Meskarbashee
Department of Agronomy and Plant Breeding, Shahid Chamran University, Ahvaz, Iran
Asian Journal of Agricultural Research
Year: 2009 | Volume: 3 | Issue: 2 | Page No.: 28-37







ABSTRACT
In order to investigate the effect of salt stress on chlorophyll content and fluorescence, sodium (Na+) and potassium (K+) ions content of rape (Brassica napus L.) plants, ten genotypes were subjected to salinity levels (control [2.5], 6, 10, 14 and 18 dS m-1) for 30 days in hydroponics. Salt treatments were imposed to genotypes in root establishment stage (4 leaves). Results showed that quantum yield of photosystem II from light adapted (ΦPSII) and dark-adapted leaf (Fv/Fm), photochemical quenching (qP) and minimal fluorescence from dark-adapted leaf (Fo) were affected by salinity. Genotypes MHA4921 and Hyola 401 had highest shoot dry weight at the two higher salt treatments (14 and 18 dS m-1) and resulted the most tolerant to salinity among the tested genotypes. Chlorophyll (chl) fluorescence attributes was generally affected by salinity stress, except in the two salt tolerant genotypes and thus could be used as a tool for screening for salinity tolerance. Chlorophyll content (SPAD units) changed significantly in all genotypes, except in salt tolerant ones. Shoots Na+ content increased, by increasing salinity levels, but in MHA4921 this increase was higher than the other genotypes and may be relation to decline in the osmotic potential of cellular contents. Rape ability to accumulate sodium in response to salinity is one of the major criteria of salt tolerance. K+ content in shoots, at the different levels of salinity in MHA4921 and Hyola 401 were higher than the other genotypes.
PDF Abstract XML References Citation
How to cite this article
V. Atlassi Pak, M. Nabipour and M. Meskarbashee, 2009. Effect of Salt Stress on Chlorophyll Content, Fluorescence, Na+
and K+ Ions Content in Rape Plants (Brassica napus L.). Asian Journal of Agricultural Research, 3: 28-37.
DOI: 10.3923/ajar.2009.28.37
URL: https://scialert.net/abstract/?doi=ajar.2009.28.37
DOI: 10.3923/ajar.2009.28.37
URL: https://scialert.net/abstract/?doi=ajar.2009.28.37
INTRODUCTION
Salt stress is a major abiotic stress problem in arid and semi-arid regions (Sudhir and Murthy, 2004) and agriculture productivity in these areas of the world is very low (Ashraf, 2004). Hence, considerable improvement in salinity tolerance is essential in crop species through conventional selection and breeding techniques (Ashraf and Harris, 2004). Many biochemical and physiological criteria or traits have been proposed for screening (Ashraf and Harris, 2004). Classical methods of screening for salt tolerance are based on yield response (Netondo et al., 2004) but the underlying genetic mechanisms for yield are complex with considerable environmental influence (Ashraf, 2004). Chlorophyll (chl) fluorescence could be used for screening for salt tolerance varieties and modified by salinity stress (Baker and Rosenqvist, 2004). Chl fluorescence provides non-invasive and rapid method for estimates of photosynthetic performance of plants (Kao et al., 2003; Baker and Rosenqvist, 2004; Zlatev and Yordanov, 2004). Effect of sodium chloride (NaCl) stress on chl fluorescence has been studied in different plants. Application of chl fluorescence for salinity tolerance was investigated in barely (Belkhodja et al., 1994), sorghum (Netondo et al., 2004), naked oat (Zhao et al., 2007), arabidopsis and thellungiella (Stepien and Johnson, 2009), rice (Moradi and Ismail, 2007) and wheat (Zair et al., 2003). A significant decline in quantum yield of photosystem II from dark-adapted leaf (Fv/Fm) accompanied by increase of non photochemical quenching (NPQ) occurred in sorghum varieties with 250 mM NaCl (Netondo et al., 2004). Moradi and Ismail (2007) reported that no significant differences in quantum yield of PSII (ΦPSII) were observed with increasing salinity level at vegetative stages in rice, but concluded that NPQ increased significantly. They suggested that no reduction in ΦPSII were observed in the tolerate lines but in the sensitive one it did. Zair et al. (2003) noted that the Fv/Fm ratio decreased significantly in salt sensitive line and remained unchanged in tolerant one in wheat. In wild soybean species, as increasing salinity, no significant differences were found in Fv/Fm and ΦPSII (Kao et al., 2003). Salinity stress significantly reduced chlorophyll content, photochemical quenching (qP) and Fv/Fm in naked oat (Zhao et al., 2007). The effect of salt stress on chl fluorescence attributes was examined in arabidopsis (Arabidopsis thaliana L.) and thellungiella (Thellungiella halophila L.) by Stepien and Johnson (2009). They concluded that increasing salinity resulted in a substantial increase in NPQ in arabidopsis (salt sensitive) while in thellungiella (salt tolerance) this parameter remained close to control levels at all salt concentrations. A considerable decrease was observed in Fv/Fm and ΦPSII in arabidopsis while in thellungiella no change occurred in Fv/Fm. In thellungiella ΦPSII did not changes in intermediate salinity.
In conclusion, results show that in some experiments (Misra et al., 2001; Zair et al., 2003; Stepien and Johnson, 2009) quantum yield of PSII was an early indicator of salt stress and provide important information on photosynthetic activity, but in studies of Belkhodja et al. (1994) and Kao et al. (2003) this parameter was not useful indicator for salt stress. The present study was conducted to investigate the effect of salt stress on chl fluorescence attributes in rapes (Brassica napus L.) genotypes known to differ in their salinity tolerance.
MATERIALS AND METHODS
This study was conducted in 2008-09 years in the growth chamber of Department of Agronomy and Plant Breeding, College of Agriculture, Shahid Chamran University, Ahvaz, Iran. The environmental conditions in the growth chamber were: Photosynthetic Active Radiation (PAR) 450 μmol/m/sec, day/night temperature 24/18°C, relative air humidity 55-70%. The tested rape genotypes were Sarigol, Ahatrol, Hyola401, Hyola308, RGS003, MHA8725, MHA4921, MHA4026, MHA9716 and MHA8716.
All seeds samples were surface sterilized with 1% sodium hypochlorite solution for 20 min and washed with distilled water. Plants were germinated in germinator for a week. After a week, seedlings were transplanted in aerated Hoagland’s solution. After the establishment of seedlings, genotypes were subjected to five level of salinity for 30 days. The experiment was designed as split plots on the basis of randomized complete design (RCD) with three replications. Salinity as main plot factor had five levels (control [2.5], 6, 10, 14 and 18 dS m-1). The genotypes were used as sub plot. NaCl (Merck) was used as a source of salt.
Chlorophyll fluorescence was measured with intact plants in the growth chamber. Measurements of chlorophyll fluorescence (with chlorophyll fluorometer, PAM-2000, Walz Germany) and chlorophyll content (SPAD units with a chlorophyll meters SPAD-502, Minolta Japan) were made on fully expanded youngest leaf (Ashraf, 2001). Potassium and sodium were determined with flame photometer (Ashraf and Ali, 2008). Analysis of variance was performed by SAS (version 6.12) (Moradi and Ismail, 2007) and MSTATC (Atlassi et al., 2008) programs. The mean values were compared by Duncan’s test.
RESULTS
Salinity and genotypes had significant effect on shoot dry matter. Shoot dry weight of ten genotypes decreased significantly with increasing NaCl concentrations in Hoagland’s solution. MHA4921 and Hyola401 had highest shoot dry weight among all genotypes at two higher salt concentrations (14 and 18 dS m-1) (Fig. 1). Comparison of different genotypes shows that MHA4026 and MHA9716 had lowest shoot dry weight (Fig. 1). Chlorophyll content (SPAD units) did not change significantly in MHA4921 and Hyola401 (Fig. 1). Quantum yield of PSII (ΦPSII and Fv/Fm) was not affected dramatically by salinity in two salt tolerant genotypes (Fig. 1, 2).
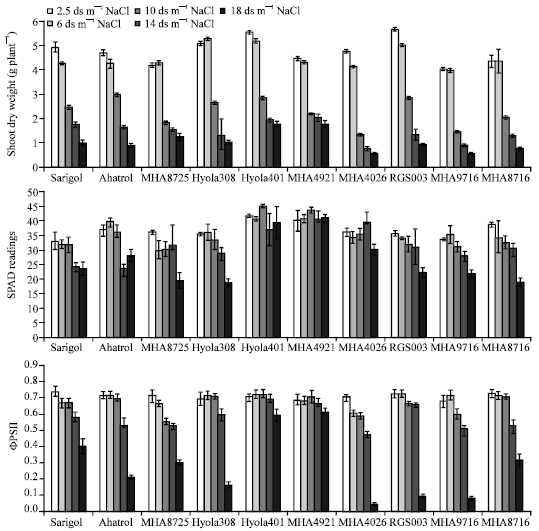 | |
| Fig. 1: | Shoot dry weight, Chlorophyll content (SPAD unites) and Quantum yield of PSII from light adapted leaf (ΦPSII) of ten genotypes of rape (B. napus). Three-week-old plants were exposed to salt for 30 days |
A trend of decrease in the SPAD values of genotypes can be observed by increasing NaCl concentration, but it was not in MHA4921 and Hyola401 (Fig. 1). Fv/Fm had a significant decrease at 14 and 18 dS m-1 NaCl concentrations (Fig. 2). Photochemical quenching (qP) showed a similar trend as Fv/Fm and at the two higher NaCl concentrations dramatically impaired (Fig. 2). ΦPSII and qP in MHA 4921 and Hyola 401 genotypes had a little decline at 18 dS m-1 NaCl (Fig. 1, 2).
In all genotypes increasing salinity resulted in increased non-photochemical quenching (NPQ) (Fig. 2). Minimal chlorophyll fluorescence (Fo) was increased by increasing salt concentration, but MHA4921 and Hyola401 were not affected (Fig. 3).
NaCl treatment resulted in accumulation of Na+ in leaves of genotypes. Salt treatmentxgenotypes interaction was not significant for Na+ content (Table 1). The data shown in Fig. 3 suggest that, by increasing salinity, in all genotypes Na+ content increased similarly. The Na+ content was the same for all genotypes at the highest NaCl concentration but in MHA4921 genotype, Na+ content was higher at the two highest salt concentrations (Fig. 3).
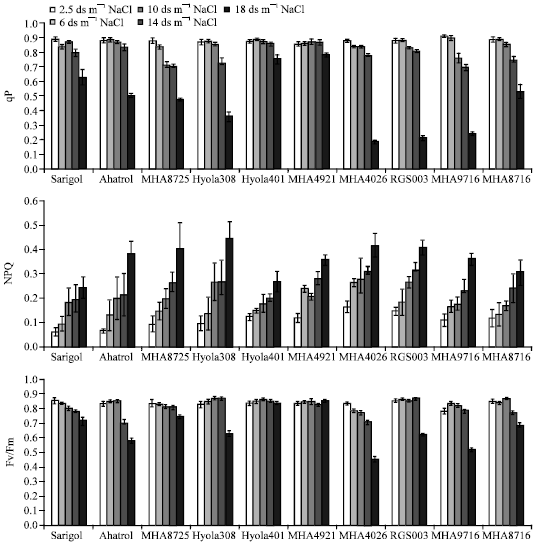 | |
| Fig. 2: | Photochemical quenching (qP), Non-photochemical quenching (NPQ) and Quantum yield of PSII from dark-adapted leaf (Fv/Fm) of ten genotypes of rape (B. napus). Three-week-old plants were exposed to salt for 30 days |
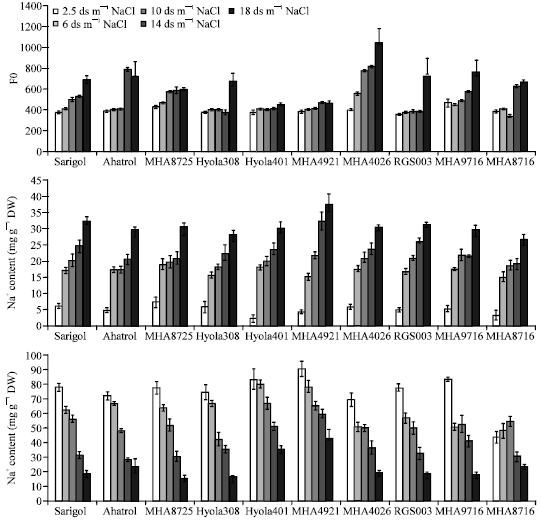 | |
| Fig. 3: | Minimal fluorescence from dark-adapted leaf (Fo), K+ and Na+ content of ten genotypes of rape (B. napus). Three-week-old plants were exposed to salt for 30 days |
| Table 1: | Mean squares from analysis of variance of data for shoot dry weight, quantum yield of photosystem II from light adapted (ΦPSII) and dark-adapted leaf (Fv/Fm), SPAD unites, Na+ and K+ ions content, photochemical quenching (qP), non-photochemical quenching (NPQ) and minimal fluorescence from dark adapted leaf (Fo) |
 | |
| *,**: Significant at 0.05 and 0.01 levels, respectively, Duncan’s test, ns: Non significant | |
Shoot K+ concentration significantly reduced in all genotypes by increasing salt treatment. As shown in Fig. 3, in two salt tolerant genotypes, K+ content was the highest at all level of salt concentrations.
DISCUSSION
Two genotypes (MHA4921 and Hyola401) had highest shoot dry weight at maximum NaCl concentration while other two (MHA4026 and MHA9716) had the lowest shoot dry weight: therefore MHA4921 and Hyola401 were ranked as salt tolerant and MHA4026 and MHA9716 were salt sensitive. By increasing of salinity, sodium ions changes the ratio of K:Na, which seems to affect the bioenergetic processes of photosynthesis (Sudhir and Murthy, 2004). Effect of salinity on plant growth may result from impairment of supply of photosynthetic assimilates (Ashraf, 2004) and cell expansion in leaves can be inhibited by salt stress (Chaves et al., 2009). Each of these parameters may results in a decline in shoot dry weight. Conservation of shoot dry weight is one of the selection criteria for salinity tolerance (Shannon, 1998). In many plant species, screening methods and the physiological works for salinity are based on the young plant stages (Dasgan et al., 2002).
Some researchers suggest that decreasing dry mass production in shoots at early vegetative growth stages is associated with decreased seed yield in rapes (Qasim et al., 2003). Shoot biomass measuring at early vegetative growth stages was introduced as a scale for show salt tolerant and sensitive variety of rapes and a positive association between shoot biomass and seed yield was found (Ashraf, 2001; Ashraf and Ali, 2008). Other authors concluded that early vegetative growth responses could be used as a trait for rapid selection of salt-resistant varieties in rice (Aslam et al., 1993). Ashraf (2001) suggested that shoot dry weight at the vegetative growth in Brassica spp. was related to the ultimate of tolerance of these species.
Chlorophyll fluorescence and analyses for Fv/Fm and ΦPSII were useful for the monitoring of salt stress in our experiment. The decrease of quantum yield of PSII (ΦPSII) at increasing NaCl concentrations, except in salt tolerant genotypes is in line with results reported by some authors on different species and cultivars (Misra et al., 2001; Zair et al., 2003). In sorghum (Netondo et al., 2004) and rice (Moradi and Ismail, 2007) no changes in this parameter under salt stress were observed. These authors noted that these plant species PSII is highly resistant to salinity stress. In the present study Fv/Fm and ΦPSII decreased significantly by increasing NaCl concentration in salt sensitive genotypes. Salinity stress is though to cause lesions in the reaction center of PSII (Yang et al., 1996; Baker and Rosenqvist, 2004), either directly or via an accelerated senescence (Netondo et al., 2004), which plays a critical role in the response of photosynthesis (Yang et al., 1996); hence ΦPSII has been widely used for the measurement of stress condition of crops (Khan et al., 2006) to detected stress-induced perturbations in photosynthetic apparatus (Baker and Rosenqvist, 2004).
Reduction in ΦPSII is on of the major factor responsible for the drastic reduction in photosynthetic rate under salt stress and differences in Fv/Fm is often used as characteristics of cultivars or species differences in soybean (Kao et al., 2003).
Photochemical quenching (qP) is a measure of the proportion of PSII reaction centers capable of photochemistry (Thioyapong et al., 2004) and changes in ΦPSII are similar to the changes in qP (Baker and Rosenqvist, 2004). These changes have been seen in our experiment where qP showed a significant decline at the highest salt concentration level, also in two salt tolerant genotypes.
The negative impact of NaCl on photosynthesis rate in our experiment, results in an increase in NPQ in all genotypes. Sudhir and Murthy (2004) noted that salt stress enhances the oxigenase activity of RUBPco and can cause a decline in Co2 fixation. Increase in NPQ may represent the decreased demand for product of electron transport, which has been using for assimilation and thus results in heat dissipation of light energy (Netondo et al., 2004; Moradi and Ismail, 2007). Increase in NPQ, might result from changes in protective high-energy-state (Stepien and Johnson, 2009) and photoinhibition (Moradi and Ismail, 2007; Stepien and Johnson, 2009). NPQ and qP has been used for screening of salt tolerant wheat genotypes (Zair et al., 2003). In our experiment, NPQ was less informative for screening of salt tolerant. In sorghum, qP significantly decreased but NPQ increased under salinity conditions (Netondo et al., 2004). Shabala et al. (1998) concluded that fluorescence quenching in leaves are the most sensitive photosynthetic characteristics for measuring salinity tolerance in maize. In this study, the high rates of qP in salt tolerant genotypes represent the high efficiency of light use for electron transport by PSII and/or ability of Hyola 401 and MHA 4921 to maintain QA partially oxidized.
Any excess of absorbed light energy can sensitize the formation of reactive oxygen species (ROS) (Thioyapong et al., 2004).
Greater ΦPSII and therefore, qP in salt tolerant plants, in our experiment, compared to the other genotypes likely contributed to the greater leaf chlorophyll content. Some authors found that higher rates of ROS in salt sensitive rape genotypes can cause damage to photosynthetic pigments (Ashraf and Ali, 2008). Present results indicated that decrease in chlorophyll content, in salt sensitive genotypes, resulted in a decrease in Fv/Fm and ΦPSII. Leaf chlorophyll content was affected by salinity in tetraploid wheat (Munns and James, 2003), rice (Sultana et al., 1999), Brassica oleracea (Bhattacharya et al., 2004), Brassica juncea (Qasim, 1998). Salinity can affect chlorophyll content through inhibition of chlorophyll synthesis or an acceleration of its degradation (Zhao et al., 2007). Thipyapong et al. (2004) found that the chlorophyll losses due to salinity stress is consistent with possible differences in ROS production among the genotypes and suggested that, in salt sensitive genotypes ROS scavenging systems were unable to detoxify ROS generated.
The chlorophyll loss in leaf results in an increase in Fo (Minimal fluorescence from dark adapted leaf), in all genotypes studied. Fo ensures that the PSII reaction centers are in the open state (Baker and Rosenqvist, 2004). Salt stress did not have any significant effect on Fo, in salt tolerant genotypes. The increase in Fo at highest salt concentration is characteristic of PSII inactivation and occurred concomitantly to the decrease in Fv/Fm (Zlatev and Yordanov, 2004). The increased of Fo may be due to reduce in plastoquinon acceptor (QA) and its ability to maintain oxidized completely (Zlatev and Yordanov, 2004).
Other researchers suggested that more accumulation of sodium (Na+) ions in shoots of salt sensitive genotypes (compared to the salt tolerant ones), is one of the most important factors affecting chlorophyll losses (Dingkuhn et al., 1992).
In our experiment, Na+ ions of each genotype were significantly increased under salinity stress, with higher values recorded in the salt tolerant genotype at highest NaCl concentration with no dramatic differences genotypes. It seems that increase in Na+ content in this genotype results in a decline in the osmotic potential of cellular contents and higher level of water uptake (Zhang et al., 2001).
Mokhamed et al. (2006) noted that Na+ content in leaves of salt tolerant of rape genotype, was higher than salt sensitive one.
On the contrary, Ashraf and McNeilly (2004) suggested that the salt tolerant plants of rape was the lowest and the salt sensitive one was the highest in shoot Na+ concentration when plants were subjected to saline conditions. Some authors pointed out that salt tolerance in some plant species has negative correlation with Na+ in plant shoots (Dasgan et al., 2002). Other researchers suggested that the ability of plants to accumulate inorganic ions (Na+ and Cl¯), in response to salinity, may be one of the major criteria of salt tolerance, rape in particular (Zhang et al., 2001).
Present results indicated that sodium influx from plant cells results in increase in resistant to salt stress as reported by Mokhamed et al. (2006) and Siddiqui et al. (2008). Ashraf and Ali (2008) in their experiments did not observe remarkable differences in Na+ content of shoots in rape genotypes under saline conditions (150 mM). Dasgan et al. (2002) reported that some salt tolerant genotypes of tomato show an inclusion mechanism for Na+ while others show exclusion mechanism. Sodium transport into the vacuole can remove toxic effect of sodium from the cytosol; these ions act as an osmoprotectant within the vacuole (Zhu, 2003). The role of sodium compartmentation in plant salt tolerance has been demonstrated in transgenic Brassica napus (Zhang et al., 2001).
Another indicator of plant salt tolerance is their ability to maintain potassium (K+) ions at high level of salinity (Blumwald et al., 2000) and high preservation of K+ content as increasing NaCl concentration, as indicated in our experiment, results in ionic homeostasis in the salt tolerant genotypes (Mokhamed et al., 2006). Protein synthesis in plant cells depends on physiological K+ concentration (Blumwald et al., 2000) and its conservation in plant cells is essential for cell metabolism.
REFERENCES
- Ashraf, M., 2004. Some important physiological selection criteria for salt tolerance in plants. Flora-Morphol. Distrib. Funct. Ecol. Plants, 199: 361-376.
CrossRefDirect Link - Ashraf, M. and P.J.C. Harris, 2004. Potential biochemical indicators of salinity tolerance in plants. Plant Sci., 166: 3-16.
CrossRefDirect Link - Ashraf, M., 2001. Relationships between growth and gas exchange characteristics in some salt-tolerant amphidiploid Brassica species in relation to their diploid parents. Environ. Exp. Bot., 45: 155-163.
CrossRef - Ashraf, M. and Q. Ali, 2008. Relative membrane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (Brassica napus L.). Environ. Exp. Bot. 63: 266-273.
CrossRef - Ashraf, M. and T. McNeilly, 2004. Salinity tolerance in brassica oilseeds. Crit. Rev. Plant. Sci., 23: 157-174.
CrossRef - Aslam, M., R.H. Qureshi and N. Ahmed, 1993. A rapid screening technique for salt tolerance in rice (Oryza sativa L.). Plant Soil, 150: 99-107.
CrossRefDirect Link - Atlassi, V., M. Meskarbashee and R. Mamghani, 2008. Effect of planting pattern on morphology, yield and yield components of three spring canola varieties in Ahvaz climatic conditions. Res. Crops, 9: 39-44.
Direct Link - Baker, N.R. and E. Rosenqvist, 2004. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot., 55: 1607-1621.
CrossRefDirect Link - Belkhodja, R., F. Morales, A. Abadia, J. Gomes-Aparisi and J. Abadia, 1994. Chlorophyll fluorescence as a possible tool for salinity tolerance screening in barley (Hordeum vulgare L.). Plant Physiol., 104: 667-673.
Direct Link - Bhattacharya, R.C., M. Maheswari, V. Dineshkumar, P.B. Kirti, S.R. Bhat and V.L. Chopra, 2004. Transformation of Brassica oleracea var. capitata with bacterial betA gene enhances tolerance to salt stress. Sci. Hortic., 100: 215-227.
CrossRef - Blumwald, E., G.S. Aharon and M.P. Apse, 2000. Sodium transport in plant cells. Biochim. Biophys. Acta (BBA)-Biomembr., 1465: 140-151.
CrossRefDirect Link - Chaves, M.M., J. Flexas and C. Pinheiro, 2009. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot., 103: 551-560.
CrossRefDirect Link - Dasgan, H.Y., H. Aktas, K. Abak and I. Cakmak, 2002. Determination of screening techniques to salinity tolerance in tomatoes and investigation of genotype responses. Plant. Sci., 163: 695-703.
CrossRef - Dingkuhn, M., S.K. De Datta, R. Pamplona, C. Javellana and H.F. Schnier, 1992. Effect of late-season N fertilization on photosynthesis and yield of transplanted and direct-seeded tropical flooded rice. II. A canopy stratification study. Field Crops Res., 28: 235-249.
Direct Link - Kao, W.Y., T.T. Tsai and C.N. Shih, 2003. Photosynthetic gas exchange and chlorophyll a fluorescence of three wild soybean species in response to NaCl treatments. Photosynthetica, 41: 415-419.
CrossRefDirect Link - Khan, S., Y. Yoshida and S. Islam, 2006. Characteristics of photosynthetic behaviors and chlorophyll fluorescence in different vegetable species. Asian J. Plant Sci., 5: 266-270.
CrossRefDirect Link - Misra, A.N., A. Srivastava and R.J. Strasser, 2001. Utilization of fast chlorophyll fluorescence technique in assessing the salt/ion sensitivity of mung bean and Brassica seedling. J. Plant Physiol., 158: 1173-1181.
CrossRef - Mokhamed, A.M., G.N. Raldugina, V.P. Kholodova and V.l.V. Kuznetsov, 2006. Osmolyte accumulation in different rape genotypes under sodium chloride salinity. Russian J. Plant Physiol., 53: 649-655.
CrossRefDirect Link - Moradi, F. and A.M. Ismail, 2007. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot., 99: 1161-1173.
CrossRefDirect Link - Munns, R. and R.A. James, 2003. Screening method for salinity tolerance: A case study with tetraploid wheat. Plant Soil, 253: 201-218.
Direct Link - Netondo, G.W., J.C. Onyango and E. Beck, 2004. Sorghum and salinity: II. Gas exchange and chlorophyll fluorescence of sorghum under salt stress. Crop Sci., 44: 806-811.
Direct Link - Qasim, A.A., 1998. Effect of salinity on growth, proline accumulation and chlorophyll content during vegetative growth, flowering and seed formation of Brassica juncea L. J. King Saud Univ. Agric. Sci., 10: 145-152.
Direct Link - Qasim, M., M. Ashraf, M.A. Jamil, M.Y. Ashraf, S.U. Rehman and E.S. Rha, 2003. Water relations and leaf gas exchange properties in some elite canola (Brassica napus) lines under salt stress. Ann. Applied Biol., 142: 307-316.
Direct Link - Shabala, S.N., L. Shabala, A.I. Martynenko, O.K. Babourina and I.A. Newman, 1998. Salinity effect on bioelectric activity, growth, Na+ accumulation and chlorophyll fluorescence of maize leaves: A comparative survey and prospects for screening. Aust. J. Plant Physiol., 25: 609-616.
Direct Link - Stepien, P. and N.G. Johnson, 2009. Contrasting responses of photosynthesis to salt stress in the glycophyte arabidopsis and the halophyte thellungiella: Role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol., 149: 1154-1165.
CrossRefDirect Link - Sudhir, P. and S.D.S. Murthy, 2004. Effects of salt stress on basic processes of photosynthesis. Photosynthetica, 42: 481-486.
CrossRefDirect Link - Sultana, N., T. Ikeda and R. Itoh, 1999. Effect of NaCl salinity on photosynthesis and dry matter accumulation in developing rice grains. Environ. Exp. Bot., 42: 211-220.
CrossRef - Thioyapong, P., J. Melkonian, D.W. Wolf and J.C. Steffens, 2004. Suppression of polyphenol oxidases increases stress tolerance in tomato. Plant Sci., 167: 693-703.
CrossRef - Zair, I., A. Chlyah, K. Sabounji, M. Tittahsen and H. Chlyah, 2003. Salt tolerance improvement in some wheat cultivars after application of in vitro selection pressure. Plant Cell Tissue Org. Cult., 73: 237-244.
CrossRefDirect Link - Zhang, H.X., J.N. Hodson, J.P. Williams and E. Blumwald, 2001. Engineering salt tolerant Brassica plants: Characterization of yield and seed oil quality in transgenic plants with increased vacuolar sodium accumulation. Proc. Nat. Acad. Sci. USA., 48: 12832-12836.
CrossRefDirect Link - Zhao, G.Q., B.L. Ma and C.Z. Ren, 2007. Growth, gas exchange, chlorophyll fluorescence and ion content of naked oat in response to salinity. Crop Sci., 47: 123-131.
CrossRefDirect Link - Zhu, J.K., 2003. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol., 6: 441-445.
CrossRefDirect Link - Zlatev, Z.S. and I.T. Yordanov, 2004. Effect of soil drought on photosynthesis and chlorophyll fluorescence in bean plants. Bulg. J. Plant Physiol., 30: 3-18.
Direct Link