
Amel Kerkeni
Higher Agronomic Institute, Chott Mariem 4042-Sousse, Tunisia
Mejda Daami-Remadi
Regional Center of Research in Horticulture and Organic Agriculture, Chott Mariem 4042-Sousse, Tunisia
Neji Tarchoun
Higher Agronomic Institute, Chott Mariem 4042-Sousse, Tunisia
Mohamed Ben Khedher
Technical Center of Organic Agriculture, Chott Mariem 4042-Sousse, Tunisia
Asian Journal of Agricultural Research
Year: 2007 | Volume: 1 | Issue: 2 | Page No.: 50-58







ABSTRACT
Seven fungi isolated from an animal manure compost are tested for their in vitro and in vivo antagonistic activity against Pythium ultimum, the causal agent of the cucumber damping-off. In vitro, dual culture experiments, observed after incubation at 25°C on PDA during 4 days, showed that six fungi inhibited by 18 to 61% the mycelial growth of P. ultimum, whereas one fungus showed no activity. Competition for media was the most remarkable mechanism of action noted on PDA. In vivo experiments, sterilized peat individually treated with Aspergillus sp., Trichoderma viride (strain 1 and strain 2), at a rate of 0.5 g L-1 and infested with P. ultimum was evaluated for its suppressive effect of the cucumber (Cucumis sativus L. Mornegui) damping-off. Results showed the tested fungi, incorporated to the culture substrates, decreased the in vivo development of P. ultimum, but at variable degrees, comparatively to the untreated control. Damping-off was lower for cucumber plants treated with T. viride’ (strain 2) than those treated with Aspergillus sp. and with T. viride (strain 1). Aspergillus sp. isolated from compost, which was inactive in vitro, suppressed the cucumber damping-off by 69.4%.
PDF Abstract XML References Citation
How to cite this article
Amel Kerkeni, Mejda Daami-Remadi, Neji Tarchoun and Mohamed Ben Khedher, 2007. In vitro and in vivo Suppression of Pythium ultimum the Causal Agent of the Cucumber Damping-Off by Some Compost Fungi. Asian Journal of Agricultural Research, 1: 50-58.
DOI: 10.3923/ajar.2007.50.58
URL: https://scialert.net/abstract/?doi=ajar.2007.50.58
DOI: 10.3923/ajar.2007.50.58
URL: https://scialert.net/abstract/?doi=ajar.2007.50.58
INTRODUCTION
Pythium ultimum is a phytopathogenic fungus of a recognised importance in nurseries production. Once introduced, infection can reach a higher level because of pathogen development and spreading to the whole cultural system. Some methods are now available to prevent infection such as the use of resistant cultivars and fungicides application. However, resistant cultivars against Pythium sp. do not exist. In the other side, few fungicides are registered against this pathogen (Le Floch et al., 2003). Consequently, other alternatives for this pathogen control are required (Rankin and Paulitz, 1994).
Composts prepared from heterogeneous organic wastes showed the potential of biological control for several plant diseases, caused by soilborne pathogens (Hoitink et al., 1991, 1997). Phytopathogenic fungi such as Phytophthora sp. (Hoitink and Boehm, 1999; Aryantha et al., 2000), Rhizoctonia solani (Nelson and Hoitink, 1983; Chung and Hoitink, 1990) and Fusarium sp. (Chef et al., 1983; Cotxarrera et al., 2002) were successfully suppressed by composts. Numerous reports of the suppressive effects of composts against damping-off, caused by Pythium sp., were also published (Chen et al., 1987; Chen et al., 1988; Mandelbaum and Hadar, 1990; Zhang et al., 1996; Diab et al., 2003).
Beneficial microorganisms present in composts are implicated in various suppressive activities (Hoitink et al., 1991) and strains of Trichoderma sp. (Cotxarrera et al., 2002) and strains of Bacillus subtilis (Phae et al., 1990) isolated from composts were effective antagonists against several pathogens. Strains of T. hamatum suppressed Fusarium oxysporum (Trialls-Gay et al., 1986). Isolates of Penicillium sp. and of Aspergillus sp. were suppressive to F. solani, F. graminearum, F. sambucinum and F. oxysporum f. sp. tuberosi (Daami-Remadi et al., 2006). Several microorganisms have been reported to be biocontrol agents for suppression of Pythium damping-off (Harman and Hadar, 1983). Daami-Remadi (2001) also showed an antagonistic activity of T. harzianum against P. aphanidermatum and P. ultimum. Antagonistic interactions with other fungi and mechanism of biological control were based on antibiosis, parasitism, induced resistance and competition for space and limited resources (Hoitink et al., 1993).
Preliminary dual culture of some compost extracts with Pythium ultimum, showed inhibition of the pathogen mycelial growth (Kerkeni et al., 2007). The aim of this study is to test individually, some fungi isolated from compost, against P. ultimum and those showing higher inhibitory effect, will be evaluated for their suppressive ability of the cucumber damping-off.
MATERIALS AND METHODS
Pathogen
P. ultimum, was isolated from potato tuber showing typical leak symptoms. This pathogen was cultured on PDA at 25°C for one week and stored at 4°C for long preservation.
Compost Fungi
A mature compost (>12 months), composed of 40% cattle manure, 40% sheep manure and 20% vegetable wastes and produced on 2006 at the composting-unit of the Technical Center of Organic Agriculture of Chott Mariem-Tunisia, was used for antagonistic fungi isolations.
Potato Dextrose Agar (PDA; Sigma) supplemented with 5 mg L-1 Penicillium-G was used for fungal isolation. A sample of 10 g of solid compost was suspended in 90 mL of sterilized distilled water in 250 mL bottle. The sample was stirred for 1 h at 200 rpm. A serial dilution up to 10-3 was carried out and then 100 μL aliquots of this dilution were spread onto PDA medium plates. After incubation at 25°C for 5 days, formed fungal colonies were individually transferred on PDA medium. The same procedure was repeated until having a purified fungal culture. A total of seven fungi were isolated.
Selected compost fungi were identified on the basis of their macroscopic and microscopic characteristics (El-Masry et al., 2002). They were sustained on PDA at 25°C.
In vitro Bioassay of the Antagonistic Activity of the Compost Fungi
The antifungal activity of each tested compost fungi against P. ultimum was studied via the dual culture technique in the Laboratory of Phytopathology of the Regional Center of Research in Horticulture and Organic Agriculture, Chott Mariem (Tunisia). The method consists of placing an active mycelial disc (6 mm in diameter) of the pathogen, 1 cm from the edge of a 9 cm Petri plate containing freshly prepared PDA medium. Another disc (6 mm) of the antagonist fungi was deposited in a diametrally opposed position 1 cm away from the other set of the plate. For untreated plates, an agar disc of P. ultimum was placed at the center of the Petri dish. All plates were then incubated at 25°C and evaluated for pathogen growth inhibition after 4 days of incubation. Three replicates were used per elementary treatment.
To determine the inhibition rate of P. ultimum by each of the tested compost fungi, the fungal growth of P. ultimum was recorded by measuring the colony diameters (average of the two perpendicular diameters). These diameters (control and treated) served for the calculation of the inhibition rate of the fungal growth. This rate is calculated according to the following formula used by Hibar et al. (2005):
In vivo Bioassay of the Antagonistic Activity of the Compost Fungi
Plant Material
Disease suppressiveness of substrates individually amended with the tested compost fungi, against P. ultimum was tested using cucumber (Cucumis sativus L. Mornegui) (Tunisian cultivar) as host plant. Seeds of cucumber were surface-disinfected in 1% sodium hypochlorite for 30 min and rinsed several times with sterile distilled water.
Preparation of Inoculum
P. ultimum was grown at 25°C on Potato Dextrose Agar (PDA). Three fungal plugs with active pathogen mycelium (0.6 cm) were used to inoculate rice grains (25 g). This substrate was already sterilized for two consecutive days (Diab et al., 2003). Inoculated rice was then incubated in the dark for 10 days at 20°C (Fuchs, 1993). After incubation, the pathogen-colonized rice was ground aseptically with a spatula and used as inoculum in this experiment.
Three fungal plugs with active mycelium of each of the tested compost fungi (0.6 cm) were also used to inoculate rice grains (25 g) and the same method was adopted for the preparation of the inoculum of the compost fungi. They were grown at 25°C.
Bioassay
Among the seven compost fungi tested in vitro, three fungi C2-4, C2-5 and C2-6, identified as Aspergillus sp., Trichoderma viride (strain 2) and T. viride (strain 1), respectively, were selected for the in vivo evaluation during 2006, in the greenhouse of the high Agronomic Institute of Chott Mariem, in Tunisia. The remaining fungi were not identified.
Sterilized peat was mixed with rice grains colonized at the rate of 0.5 g L-1 of P. ultimum and 0.5 g L-1 of each tested compost fungi. For an uniform distribution of the inoculum in the substrate, 0.5 g of rice cultures were mixed with 50 g of sterilized sand. The mixture was then homogenized with the substrate (peat) and was distributed equally into 10 cm-diameter plastic pots. Eight cucumber seeds (Cucumis sativus L. Mornegui) were sown at equal circular spacing in each pot.
Bioassay was conducted under greenhouse conditions at 25°C and under a photoperiod of 12 h dark/12 h light. The plants were watered regularly. Five replicate pots of each elementary were used and the whole bioassay was repeated twice.
Disease Assessment
Final evaluations of the damping-off suppressiveness were made after 14 days. The disease intensity was estimated on treated and inoculated plants comparatively to the untreated and non inoculated controls. The Disease Suppressiveness (DS) of the substrates amended with compost fungi, against P. ultimum was calculated as:
Where, the disease intensity in the substrates without compost fungi (DIncf) was calculated as:
And the disease intensity in the substrates supplemented with compost fungi (DIcf) was calculated as:
| Where, | ||
| HPncf,0 | = | No. of healthy plants for the control b. |
| HPncf,i | = | No. of healthy plants for the control a. |
| HPcf,0 | = | No. of healthy plants for the substrates with either of the three tested compost fungi and non inoculated with P. ultimum (0). |
| HPcf,i | = | No. of healthy plants for the substrates inoculated with P. ultimum (i) and with either of the three tested compost fungi. |
At the end of the bioassay, the mean shoot and root fresh weights of plants per elementary treatment were also determined.
Detection of Cellulase
The cellulase enzyme is a lysogenic enzyme that could have a possible role in fungal degradation. It was analysed in the substrates amended with either of the three tested fungi, according to the method of Alef and Nannipieri (1995). Cellulase activity was expressed as micrograms of hydrolyzed glucose per gram dry weight of substrate per 24 h.
RESULTS
In vitro Inhibition of the P. ultimum Growth by the Tested Compost Fungi
The results in Table 1 showed that, of the seven compost fungi tested, six fungi inhibited the mycelial growth of P. ultimum by more than 18%, comparatively to the untreated control. The most effective fungus was C2-5, where the pathogen development was of about 61%. Whereas, the fungal isolate C2-4 showed no inhibitory effect. The compost fungi C2-6 inhibited P. ultimum growth by 33% (Table 1).
Competition, traduced by overlapping of the pathogen colonies by tested fungal isolates, was the main mechanism, employed by the most effective antagonist to P. ultimum.
In vivo Inhibition of the P. ultimum Growth by Some Compost Fungi
The two isolates C2-5 and C2-6, showing an inhibition rate of more than 30% in vitro, were identified as T. viride strain 2 and strain 1, respectively. The isolate C2-4 showing no in vitro inhibitory effect against P. ultimum was identified as Aspergillus sp. Those three fungi were selected for their in vivo antagonistic activity evaluation. Results showed that those compost fungi were suppressive to the damping-off caused by P. ultimum (Table 2).
| Table 1: | Inhibition of the P. ultimum mycelial growth in presence of some compost fungi isolates (PDA, after four days of incubation at 25°C) |
 | |
| Compost isolates C2-1, C2-2, C2-3 and C2-7 were not identified; Isolates C2-4, C2-5 and C2-6 were identified as Aspergillus sp., Trichoderma viride (strain 2) and T. viride (strain 1), respectively. They were selected for in vivo evaluation. Each value represents the mean of 3 values. Different letter(s) within columns represent values that are significantly different at p = 0.05 based on ANOVA and Duncan test | |
| Table 2: | Suppression percentage of cucumber damping-off, caused by P. ultimum, by some compost fungi |
| Each value represents the mean of 5 values, Different letters within columns represent values that are significantly different at p = 0.05 based on ANOVA and Duncan test | |
| Table 3: | Cellulase activity noted in the three substrates amended with tested compost fungi and inoculated with Pythium ultimum |
| Each value represents the mean of 3 values. Different letter(s) within columns represent values that are significantly different at p = 0.05 based on ANOVA and Duncan test | |
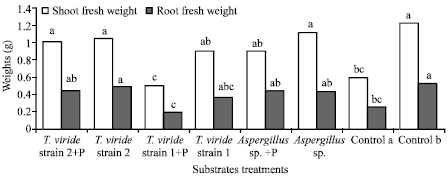 | |
| Fig. 1: | Shoot and root fresh weights of 14 days old cucumber seedlings. Control a: Substrates with P. ultimum alone; Control b: Substrates non inoculated. Each value represents the mean of 5 values. Different letter(s) within columns represent values that are significantly different at p = 0.05 based on ANOVA and Duncan test |
Suppression percentage of P. ultimum was significantly higher in substrate amended individually with T. viride strain 2 (C2-5) and Aspergillus sp. (C2-4). Contrarily to the in vitro results, Aspergillus sp. which found to be inefficient, showed an important suppression rate (69.4%)) in vivo. Suppressive activity induced by T. viride strain 1 (C2-6) was lower than that noted in in vitro tests (10.4%).
Shoot and Root Fresh Weights
Figure 1 and 2 showed that the pathogen presence in the substrates resulted in a pronounced decrease in the cucumber shoot and root fresh weights, compared to the non inoculated control (control b) and to substrates individually amended with the tested fungi. Presence of P. ultimum alone in the substrate decreased significantly the fresh weight of cucumber plants in comparison to substrates with the two most effective compost fungi Aspergillus sp. and T. viride strain 2. The reduction in shoot and root fresh weights amounted to 51.67 and 50%, respectively in the control a, inoculated with P. ultimum alone. No shoot fresh weight reduction was noted in the presence of T. viride strain 2 (Fig. 1). Whereas, root fresh weight reduction for this treatment, was not very important compared to the non inoculated control (control b).
Cellulase Activity
The results showed that cellulase enzyme was present in the tested substrates. Table 3 indicates that the highest cellulase activity was detected in the culture substrate amended with T. viride (strain 2) (136 μg Glucose/g/24 h) and in that supplemented with Aspergillus sp. (131.3 μg Glucose/g/24 h). Whereas, the least cellulase activity (94.58 μg Glucose/g/24 h) was obtained in the substrate amended with T. viride (strain 1).
 | |
| Fig. 2: | In vivo bioassay illustrating the efficacy of the three selected compost fungi in the suppression of damping-off disease by P. ultimum in 14 days old cucumber seedlings. Control a: Substrate with P. ultimum alone; Control b: Substrates non inoculated |
DISCUSSION
These results showed for the first time in Tunisia that fungi isolated from an animal manure compost are suppressive against the phytopathogenic fungus Pythium ultimum. In a previous study, Daami-Remadi (2001) reported a suppressive effect against P. ultimum induced by a local T. harzianum isolated from soil.
The current study showed that some of the tested compost fungi were able of inhibiting the mycelial growth of P. ultimum and suppressing the cucumber damping-off. This results joined that obtained by Phae et al. (1990), Zhang et al. (1998) and Bess (2000), showing that compost contain microorganisms suppressive to plant pathogens. The effectiveness of compost fungi against plant diseases as biocontrol agents was previously found by Daami-Remadi et al. (2006), where an antagonistic effect of some filamentous fungi, isolated from compost, was noted against the Fusarium sp. complex, causing dry rot of potato tubers. El Masry et al. (2002) and Muhammad and Amusa (2003) isolated from compost, several fungal microorganisms such as Aspergillus niger, Rhizopus sp., Drechslera sp. and Trichoderma harzianum, showing an inhibitory effect against Pythium aphanidermatum, Fusarium oxysporum and Rhizoctonia solani.
In vitro, all fungi tested induced P. ultimum growth reduction by more than 18%, excepting Aspergillus sp., seeming to be inactive via dual culture experiments. T. viride, (strain 2) was the most effective in reducing the mycelial growth of P. ultimum where the inhibition ratio was of about 61%. Previous work conducted in vitro by Kerkeni et al. (2007), showed that the whole compost extract used for fungi isolation, inhibited the growth of this same isolate of P. ultimum by only 14.5%. This suggests that composts and compost extracts contain biocontrol agents that are more efficient when used alone and other agents with lesser inhibitory effect when used individually, probably this is the case with Aspergillus sp.
Competition for nutrients present in the media, traduced by an overlapping of the pathogen colonies by that of the tested fungi, was the main mechanism, employed by the compost fungi during antagonism of P. ultimum in dual culture. According to Zhang et al. (1996), competition for carbon can partially explain the suppressive effect noted against Pythium sp. Whereas, for Mandelbaum and Hadar (1990), competition for nutrients was considered an important mechanism of suppression of the seedling damping off caused by Pythium aphanidermatum.
The variable efficiency of the tested fungi may be attributed to a variable mode of action and/or a variable type of antifungal metabolite produced by the antagonists (Williams and Asher, 1996).
The overlapping of the P. ultimum colonies by antagonists colonies is probably due to a physical contact between pathogen and the compost fungi and parasitism thus, mycelium destruction may occur. In fact, multiple mechanisms of action including mycoparasitism, lysis, induction of mycelial cords and early chlamydospores formation were observed in dual cultures of some compost fungi with four species (Ayed et al., 2006; Daami-Remadi et al., 2006).
It is known that the in vitro bioassays results may be different with those obtained under in vivo conditions (Inam-ul-Haq et al., 2003). In fact, in the present study, the selected potential boicontrol agents are also tested, on the basis of their high inhibition ratio, for their ability to suppress the cucumber Pythium damping-off under greenhouse conditions.
Contrarily to the in vitro assay, bioassay conducted in vivo, showed that Aspergillus sp. significantly suppressed the disease severity on cucumber seeds and showed higher antifungal activity comparatively to T. viride (strain 1). Tests based on in vitro mycelial inhibition do not always correlate with biocontrol efficacy under natural conditions. This finding proved that in vitro and in vivo results may be divergent as may be due to the variable physical and chemical properties within niches occupied by the biocontrol agents who may affect both root colonization and expression of biocontrol mechanisms.
Howell (2003) reported that biocontrol agents produce enzymes such as chitinase, protease and cellulase. These enzymes have been proved to be involved in the antagonistic activity, they act by breaking down and dissolving the polysaccharides, responsible for the rigidity of fungal cell wall. Cellulase enzyme having the ability to degrade and to hydrolyze the fungal cell wall, was detected in the culture substrates used in this current study. A higher enzyme activity of cellulase was noted in the culture substrate supplemented with T. viride (strain 2) and with Aspergillus sp. (136 and 131.3 μg Glucose/g/24 h, respectively). Whereas, lesser activity was noted with T. viride (strain 1) (94.58 μg Glucose/g/24 h) which was also correlated to a lesser suppressive effect in vivo against P. ultimum. However, in vivo suppression of P. ultimum could not be only attributed to the action of this enzyme, since an important cellulase activity was also noted in the substrate inoculated only with the pathogen. Probably this enzyme was produced by all the fungi, including the pathogen, for the decomposition of organic matter contained in the substrates.
Aspergillus sp. was able to suppress P. ultimum only in vivo, probably by the induction of systemic resistance on cucumber plants, which seems to be not frequently detected in vitro (Schisler et al., 1997).
These results showed that compost fungi are suitable products to suppress plant pathogenic fungi. All tested fungi showed an in vivo antagonistic activity against P. ultimum. Thus, they could be a promising way for the biological control of plant diseases and could reduce the need of fungicides use and they are safe for the environment preservation.
REFERENCES
- Aryantha, I.P., R. Crossc and D.I. Guest, 2000. Suppression of Phytophthora cinnamomi in potting mixes amended with uncomposted and composted animal manures. Phytopathology, 90: 775-782.
Direct Link - Ayed, F., M. Daami-Remadi, F. Jabnoun-Khiareddine and M. El Mahjoub, 2006. Potato vascular Fusarium wilt in Tunisia: Incidence and biocontrol by Trichoderma sp. Plant Pathol. J., 5: 92-98.
CrossRefDirect Link - Chef, D.G., H.A.J. Hoitink and L.V. Madden, 1983. Effects of organic components in container media on suppression of Fusarium wilt of chrysanthemum and Flax. Phytopathology, 73: 279-281.
Direct Link - Chen, W., H.A.J. Hoitink and A.F. Schmitthenner, 1987. Factors affecting suppression of Pythium damping-off in container media amended with composts. Phytopathology, 77: 755-760.
Direct Link - Chen, W., H.A. Hoitink and L.V. Madden, 1988. Microbial activity and biomass in container media for predicting suppressiveness to damping-off caused by Pythium ultimum. Phytopathology, 78: 1447-1450.
Direct Link - Chung, Y.R. and H.A.J. Hoitink, 1990. Interactions between thermophilic fungi and Trichoderma hamatum in suppression of Rhizoctonia damping-off in a bark compost-amended container medium. Phytopathology, 80: 73-77.
Direct Link - Cotxarrera, L., M.I. Trillas-Gay, C. Steinberg and C. Alabouvette, 2002. Use of sewage sludge compost and Trichoderma asperellum isolates to suppress Fusarium wilt of tomato. Soil Biol. Biochem., 34: 467-476.
CrossRefDirect Link - Daami-Remadi, M., H. Jabnoun-Khiareddine, F. Ayed, K. Hibar, I.E.A. Znaidi and M. El Mahjoub, 2006. In vitro and in vivo evaluation of individually compost fungi for potato fusarium dry rot biocontrol. J. Boil. Sci., 6: 572-580.
CrossRefDirect Link - Diab, H.G., S. Hu and D.M. Benson, 2003. Suppression of Rhizoctonia solani on impatiens by enhanced microbial activity in composted swine waste-amended potting mixes. Am. Phytopathol. Soc., 93: 1115-1123.
CrossRefPubMedDirect Link - El-Masry, M.H., A.I. Khalil, M.S. Hassouna and H.A.H. Ibrahim, 2002. In situ and in vitro suppressive effect of agricultural composts and their water extracts on some phytopathogenic fungi. World J. Microbiol. Biotechnol., 18: 551-558.
CrossRef - Hibar, K., M. Daami-Remadi, H. Jabnoun-Khiareddine and M. El Mahjoub, 2005. Effet inhibiteur in vitro et in vivo du Trichoderma harzianum sur Fusarium oxysporum f. sp. Radicis-lycopersici. Biotecnnol. Agron. Soc. Eviron., 9: 163-171.
Direct Link - Hoitink, H.A.J., Y. Inbar and M.J. Boehm, 1991. Status of compost-amended potting mixes naturally suppressive to soilborne diseases of floricultural crops. Plant Dis., 75: 869-873.
CrossRefDirect Link - Hoitink, H.A.J., M.J. Boehm and Y. Hadar, 1993. Mechanisms of suppression of soilborne plant pathogens in compost-amended substrates. In: Science and Engineering of Composting: Design, Environmental, Microbiological and Utilization Aspects, Hoitink, H.A.J. and H.M. Keener (Eds.). Ohio State University.
- Hoitink, H.A.J., A.G. Stone and D.Y. Han, 1997. Suppression of plant diseases by composts. HortScience, 32: 184-187.
Direct Link - Hoitink, H.A.J. and M.J. Boehm, 1999. Biocontrol within the context of soil microbial communities: A substrate-dependent phenomenon. Annu. Rev. Phytopathol., 37: 427-446.
CrossRefDirect Link - Howell, C.R., 2003. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis., 87: 4-10.
CrossRefDirect Link - Inam-ul-Haq, M., N. Javed, R. Ahmad and A. Rehman, 2003. Evaluation of different strains of Pseudomonas fluorescens for the biocontrol of fusarium wilt of chickpea. Plant Pathol. J., 2: 65-74.
CrossRefDirect Link - Kerkeni, A., M. Daami-Remadi, N. Tarchoun and M. Ben Khedher, 2007. In vitro assessment of the antifungal activity of several compost extracts obtained from composted animal manure mixtures. Int. J. Agric. Res., 2: 786-794.
CrossRefDirect Link - Le Floch, G., P. Rey, F. Deniel, N. Benhamou, K. Picard and Y. Tirilly, 2003. Enhancement of development and induction of resistance in tomato plants by antagonist, Pythium oligandrum. Agronomie, 23: 455-460.
CrossRefDirect Link - Mandelbaum, R. and Y. Hadar, 1990. Effects of available carbon source on microbial activity and suppression of Pythium aphanidermatum in compost and peat container media. Phytopathology, 80: 794-803.
Direct Link - Muhammad, S. and N.A. Amusa, 2003. In vitro inhibition of growth of some seedling blight inducing pathogens by compost-inhabiting microbes. Afr. J. Biotechnol., 2: 161-164.
Direct Link - Nelson, E.B. and H.A.J. Hoitink, 1983. The role of microorganisms in the suppression of Rhizoctonia solani in container media amended with composted hardwood bark. Phytopathology, 73: 274-278.
Direct Link - Phae, C.G., M. Sasaki, M. Shoda and H. Kubota, 1990. Characteristics of Bacillus subtilis isolated from composts suppressing phytopathogenic microorganisms. Soil Sci. Plant Nut., 36: 575-586.
CrossRefDirect Link - Schisler, D.A., P.J. Slininger and R.J. Bothast, 1997. Effects of antagonist cell concentration and two strain mixtures on biological control of Fusarium dry rot of potatoes. Phytopathology, 87: 177-183.
CrossRefPubMedDirect Link - Trillas-Gay, M.I., H.A.J. Hoitink and L.V. Madden, 1986. Nature of suppression of Fusarium wilt of radish in a container medium amended with composted hardwood bark. Plant Dis., 70: 1023-1027.
Direct Link - Williams, G.E. and M.J.C. Asher, 1996. Selection of rhizobacteria for the control of Pythium ultimum and Aphanomyces cochlioides on sugar-beet seedlings. Crop Prot., 15: 479-486.
CrossRefDirect Link - Zhang, W., W.A. Dick and H.A.J. Hoitink, 1996. Compost-induced systemic acquired resistance in cucumber to Pythium root rot and Anthracnose. Phytopathology, 86: 1066-1070.
Direct Link - Zhang, W., D.Y. Han, W.A. Dick, K.R. Davis and H.A.J. Hoitink, 1998. Compost and compost water extract-induced systemic acquired resistance in cucumber and arabidopsis. Phytopathology, 88: 450-455.
CrossRefPubMedDirect Link