
Hatem Mohamed El-Zefzaf
Department of Seed Pathology Research, Institute of Plant Pathology Research, Agriculture Research Center, Giza, Egypt
LiveDNA: 20.29424
Asian Journal of Applied Sciences
Year: 2020 | Volume: 13 | Issue: 4 | Page No.: 144-151







ABSTRACT
Background and Objective: Peanut is one of the most important crops all over the world. Global annual production of peeled peanuts reached 44 million t in 2016 and China produces 38% of total production. Peanut attacked by many of fungi which causes lot of quality or quantity losses The present investigation concerned with study of peanut seed-borne fungi and their extracellular enzyme activities under Egyptian condition. Materials and Methods: Commercial seed samples were collected from different four governorates with different conditions such as; Beheira, Ismailia, Giza and Sohag. All samples were subjected to seed healthy test using blotter method. Therefore, the most frequent fungi subjected to study their extra cellular enzyme activity. Results: Seventeen species of fungi belonging to 14 genera were isolated. Six fungi were recorded in all locations such as; Aspergillus flavus, Fusarium moniliforme, Fusarium oxysporum, Penicillium sp., Rhizoctonia solani and Aspergillus niger. The best activity for most enzymes was at 30°C. Conclusion: Tested fungi were varied in their enzyme activities between three evaluated enzymes such as; amylase, cellulase and lipase. Aspergillus flavus showed a remarkable activity of amylase and lipase while Rhizoctonia solani distinctive in cellulase.
PDF Abstract XML References Citation
Received: December 26, 2019;
Accepted: January 24, 2020;
Published: September 15, 2020
How to cite this article
Hatem Mohamed El-Zefzaf, 2020. Fungi Associated with Peanut Seeds and Their Extracellular Enzyme Activities under the Influence of Physical Factors. Asian Journal of Applied Sciences, 13: 144-151.
DOI: 10.3923/ajaps.2020.144.151
URL: https://scialert.net/abstract/?doi=ajaps.2020.144.151
DOI: 10.3923/ajaps.2020.144.151
URL: https://scialert.net/abstract/?doi=ajaps.2020.144.151
INTRODUCTION
Peanut (Arachis hypogaea L.) is an important crop grown in the worldwide especially in sandy or yellow soil. Peanut originated in South American and spreads beyond China, Africa, Indian, Japan and United States of America, etc. This crop achieves cash yield more meaningful when the farmers follows bilateral or trilateral agricultural with planting early maturing varieties such as; a Giza 5 and 6. Peanut is one of legume crop which plays an important role of soil fertilization. Treating cultivated legume seeds with co-inoculation with rhizobacteria (PGPR) and rhizobia lead to enhancement and stabilize nitrogen which releases from the remaining nodules on plants roots. This amount of nitrogen improves soil fertility for next crop1,2. Peanut is also one of the important crops either as an economic crop as a result of export or because it is an important oil crop.
Peanut attacked by many economically important pathogenic fungi. These fungi invade all parts of the plant (root, stem, leaf and pod) directly or through wounds. Several researchers isolated many fungi such as; Aspergillus niger, Aspergillus flavus, Alternaria dianthicola, Curvularia lunata, Curvularia pellescens, Fusarium oxysporum, Fusarium equiseti, Macrophominapha seolina, Rhizopus stolonifer, Penicillium digitatum and Penicillium chrysogenum, etc3. These fungi cause different symptoms such as; discoloration, rotting, shrinking, seed necrosis, loss in germination capacity and toxification or change properties to oil seeds3,4. Janardhan et al.5 noted that Aspergillus flavus is a common mold in tropical and subtropical environments and secret aflatoxin as a result of molding after badly stored of peanut seeds.
Amylases are a group of enzymes which plays an important role in degrading starch links and glycogen molecules. These enzymes refer to glycoside hydrolase enzymes group which have enzyme commission number (EC: 3.2.1). These enzymes are widely distributed in the microbial plant (bacteria and fungi) and animal kingdoms and varying in action patterns according to the source. Amylases have significant applications in food fermentation, textile, paper and pharmaceutical industries6. Generally, amylase group represents about 30% of the world enzyme production7.
Bio-degradation of cellulose is a very important process which reduces the environmental pollution by converting the cellulosic biomass to fermentable sugars through biocatalyst cellulases obtained from cellulolytic organisms. Also, there is much importance of the cellulase in the industry that used for many years in textile production, food processing, waste-water treatment and detergent8. Other practical uses of cellulase are for beer production of wine, production of juice and also for production of ethanol and biofuels.
Microbial lipases are biocatalysts with distinct properties, as action under reasonable conditions, stability in organic solvents and specific substrate9. Lipases have achieved a notable position in the world market of the enzymes, clearly demonstrated by the amount of information reported in the literature, which comprises over 1,000 publications per year10. The present study was aimed to isolate and detect seed-borne fungi of peanut seeds and focus on enzymes produced by the most important frequently fungi as well as study the effect of temperature on these enzymes activities.
MATERIALS AND METHODS
Seed health test: Commercial samples of peanut seeds were collected from different four locations Beheira, Ismailia, Giza and Sohag governorates during season of 2017. Four hundred seed of each sample were used to seed a healthy test according to ISTA11. Seeds were plated in pre-sterilized Petri dishes containing 3 layers of wet blotters with rate 10 seeds in each Petri dish. All Petri dishes were incubated at 25°C for 7 days. Seeds were examined under stereomicroscope and microscope. The purification of fungi were made by transferring hyphal tips to Petri dishes containing PDA by using fine glass capillary tubes. All cultures were identified according to description sheets of Common Wealth Mycological Institute Kew12-19.
Inoculum preparation: The most frequently and most prevalent fungi which recorded in all location were selected for the further experiments. Selected fungi were grown on PDA medium for 7 days at 25°C. Then equal disks 0.5 mm were made by Cork Borer.
Preparation of media: The medium used in these experiments was mineral media containing (KH2PO4 0.14 g, NH4NO3 1 g, KCl 0.5 g, MgSO4.7H2O 0.01 g and FeSO4.7H2O 0.001 g) supplemented by the tested source of carbon as a sole source20. All these components dissolved in 1000 mL distilled water at pH 6.5 and autoclaved at 121°C for 15 min.
Amylase activity: In the present study, previous mentioned mineral media was supplemented by polysaccharides substrate (soluble starch) with rate 2 g/1000 mL of the medium. Erlenmeyer flasks 250 mL were filled with 100 mL of medium and autoclaved at 121°C for 15 min. Flasks were inoculated by one disk of each fungus separately with 3 replicates. All flasks were incubated in a shaking incubator for 48 h at 120 rpm and 30°C.
Extraction of crude amylase: The broth cultures were centrifuged at 7000 rpm for 30 min. Supernatants cell-free were used for estimation of the amylase activity.
Amylase assay: One milliliter of each extracted supernatant was transferred by pipette into the test tube and 1.0 mL of 1% soluble starch in citrate phosphate buffer pH 6.5 was added. All test tubes incubated at 30°C for 30 min. About 2 mL Dinitrosalicylic Acid (DNS) was added as a reagent to the reaction and the reaction was stopped by boiling the mixture for 5 min in water bath and cooled to room temperature and 10 mL of distilled water was added and color intensity was measured by using spectrophotometer20 at 540 nm.
Effect of temperature on amylase activity: Sterilized flasks filled with 100 mL medium, mentioned in the previous experiment were divided into 6 groups each 1 including all tested fungi with 3 replicates. All treatments incubated at different temperature 5, 10, 20, 30, 40 and 50°C for 48 h. Activity of amylase enzyme was evaluated in each treatment.
Cellulase activity: In this experiment, mineral medium mentioned before supplemented by cellulose carboxymethyl cellulose (CMC) with rate 2 g/1000 mL of medium as a solo source of carbon. Flasks (250 mL) were filled with 100 mL of medium and autoclaved at 121°C for 15 min. Flasks were inoculated by 1 fungal disk (0.5 cm) of each one of the tested fungi with three replicates. All treatments were incubated in a shaking incubator for 48 h at 120 rpm and 30°C temperatures.
All cultures were centrifuged at 7000 rpm for 30 min. Supernatants cell-free were used for the cellulase estimation.
Cellulase assay: One milliliter of each filtrated supernatants was pipetted into a test tube and 1.0 mL of 1% CMC in citrate phosphate buffer pH 6.5 was added. All test tubes incubated at 30°C for 30 min. Then treated with 2 mL DNS and the reaction was stopped by boiling for 5 min, in water bath and cooled to room temperature then 10 mL of distilled water was added and color intensity21 was measured using spectrophotometer at 540 nm.
Effect of temperature on cellulase activity: Sterilized flasks with medium mentioned in the previous experiment were inoculated with tested fungi each one separately. All treatments incubated at different temperature 5, 10, 20, 30, 40 and 50°C with 3 replicates for 2 days. Activity of cellulase enzyme was determined by using spectrophotometer at 540 nm as mentioned before21.
Lipase activity: In the present work, medium mentioned before was supplemented by olive oil with rate 10 mL L–1 as a solo source of carbon. About 250 mL flasks filled with medium sterilized and inoculated by one of the equal disks of tested fungi. All flasks incubated at 30°C for 48 h.
Lipase assay: The activity of lipase was determined by using the titrimetric method by using pipette, 3.5 mL of tris buffer and 3 mL of olive oil were transferred into 50 mL Erlenmeyer flasks. All flasks placed in checker water bath at 60 rpm and 37.5°C for 10 min. About 1 mL of crude enzyme was added to previous reaction substrate then incubated at 30°C for 30 min at 60 rpm with keeping the blank tube at -4°C. The reaction was inhibited by adding 3 mL of 95% ethanol solution buffer and 4 drops of phenolphthalein solution were then added.
The amount of fatty acids released during the reaction is determined by direct titration with NaOH 0.05 Mm and the pink color indicate the end point22:
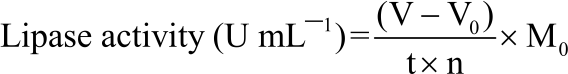 |
where, V is the volume of NaOH solution consumed in the sample solution (mL), V0 is the volume of NaOH solution consumed in blank solution (mL), t is the reaction time (min), n is the volume of enzyme solution (mL) and M is the concentration of NaOH solution.
Effect of temperature on lipase activity: Flasks (250 mL) containing 100 mL medium mentioned in the previous experiment were inoculated with tested fungi with three replicates for each tested fungus. All treatments incubated at different temperature 5, 10, 20, 30, 40 and 50oC for 2 days. Activity of lipase enzyme was evaluated in each treatment.
RESULTS AND DISCUSSION
Seed health test: Seventeen fungal species belonging to 14 genera were isolated from samples of peanut seeds collected from different four locations i.e., Beheira, Ismailia, Giza and Sohag governorates by using blotter method. In Table 1, Fusarium moniliforme and Aspergillus flavus were the most incidence at Beheira governorate followed by Rhizoctonia solani, Aspergillus niger and Penicillium sp.
| Table 1: Incidence and frequency of seed-borne fungi of peanut seeds collected from different 4 Egyptian governorates | ||||||
| Incidence at each location (%) | ||||||
| Isolates | Beheira | Ismailia | Giza | Sohag | Total Incidence | Frequency (%) |
| Alternaria alternate | 6 | 4 | 4 | 9 | 23 | 5.3 |
| Aspergillus flavus | 14 | 22 | 13 | 11 | 60 | 13.7 |
| Aspergillus niger | 11 | 9 | 11 | 15 | 46 | 10.5 |
| Curvularia sp. | 4 | - | 3 | - | 7 | 1.6 |
| Chaetomium sp. | 6 | 5 | - | - | 11 | 2.5 |
| Cephalosporium sp. | 2 | - | 3 | 2 | 7 | 1.6 |
| Cladosporium sp. | 4 | 2 | 4 | 1 | 11 | 2.5 |
| Fusarium solani | 5 | 7 | 2 | 2 | 16 | 3.7 |
| Fusarium moniliforme | 15 | 10 | 11 | 14 | 50 | 11.4 |
| Fusarium oxysporum | 11 | 16 | 12 | 9 | 48 | 11 |
| Mucor sp. | - | - | 2 | - | 2 | 0.5 |
| Penicillium sp. | 11 | 17 | 7 | 13 | 48 | 11 |
| Rhizoctonia solani | 13 | 10 | 9 | 14 | 46 | 10.5 |
| Rhizopus nigricans | 9 | 5 | 3 | 12 | 29 | 6.6 |
| Sclerotium rolifsii | 4 | 5 | - | 5 | 14 | 3.2 |
| Stemphylium sp. | 3 | - | - | 1 | 4 | 0.9 |
| Trichoderma spp. | 5 | 8 | 3 | - | 16 | 3.7 |
At Ismailia Aspergillus flavus recorded the highest incidence followed by Penicillium sp. and Fusarium oxysporum. Aspergillus flavus, Fusarium oxysporum, Fusarium moniliforme and Aspergillus niger appeared as the most incidence at Giza governorate. Aspergillus niger, Fusarium moniliforme and Rhizoctonia solani recorded the highest incidence at Sohag governorate.
From Table 1, it can be noticed that 10 isolates were recorded in all tested locations such as; Alternaria alternate, Aspergillus flavus, Aspergillus niger cladosporium sp., Fusarium solani, Fusarium moniliforme, Fusarium oxysporum, Penicillium sp., Rhizoctonia solani and Rhizopus nigricance. While, Cephalosporium sp., Sclerotium rolifsii and Trichoderma spp. recorded in 3 locations. Three fungal isolates were isolated from 2 locations Curvularia sp., Chaetomium sp. and Stemphylium sp., while Moucor sp. was recorded only in Giza. Generally, Aspergillus flavus, Fusarium Moniliforme, Fusarium oxysporum, Penicillium sp., Aspergillus niger and Rhizoctonia solani were the most frequently and most prevalent in all tested locations. Elwakil and El-Metwally3 isolated 27 fungal species from the 50 commercial markets samples of Egypt.
Amylase activity: In the present study mineral media supplemented by polysaccharides substrate (soluble starch) with rate 2 g/1000 mL of medium was used to determine the ability of most prevalent and the most frequent fungi for secretion amylase enzyme. The 6 tested isolates such as; Aspergillus flavus, Fusarium moniliforme, Fusarium oxysporum, Penicillium sp., Aspergillus niger and Rhizoctonia solani showed different activity of amylase.
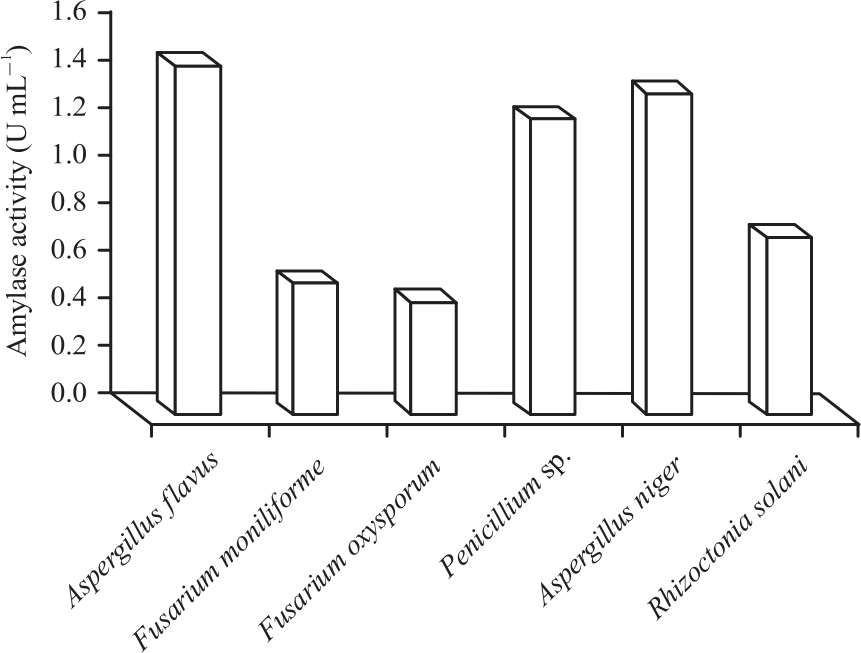 |
| Fig. 1: | Amylase activity of most prevalent fungal isolates from peanut seeds collected from different 4 governorates in Egypt |
Results presented in Fig. 1 which indicated that activity of amylase secreted by Aspergillus flavus, Aspergillus niger and Penicillium sp. was active far enough comparing with that secreted by the other three isolates. While, amylase secreted by Fusarium oxysporum was appeared with the less activity followed by Fusarium Moniliforme. Medium amylase activity was recorded by Rhizoctonia solani.
The optimum enzyme production by the all Aspergillus strain23 was found at 30°C.
Effect of temperature on amylase activity: Data present in Fig. 2 showed that all tested fungi recorded maximum activity at 30oC and this activity decrease as temperature decreased or increased. Both Aspergillus flavus, Aspergillus niger and Penicillium sp. are characterized by thermal stability in range of temperature between 30-40°C, while Rhizoctonia solani characterized by thermal stability between 20 and 40°C.
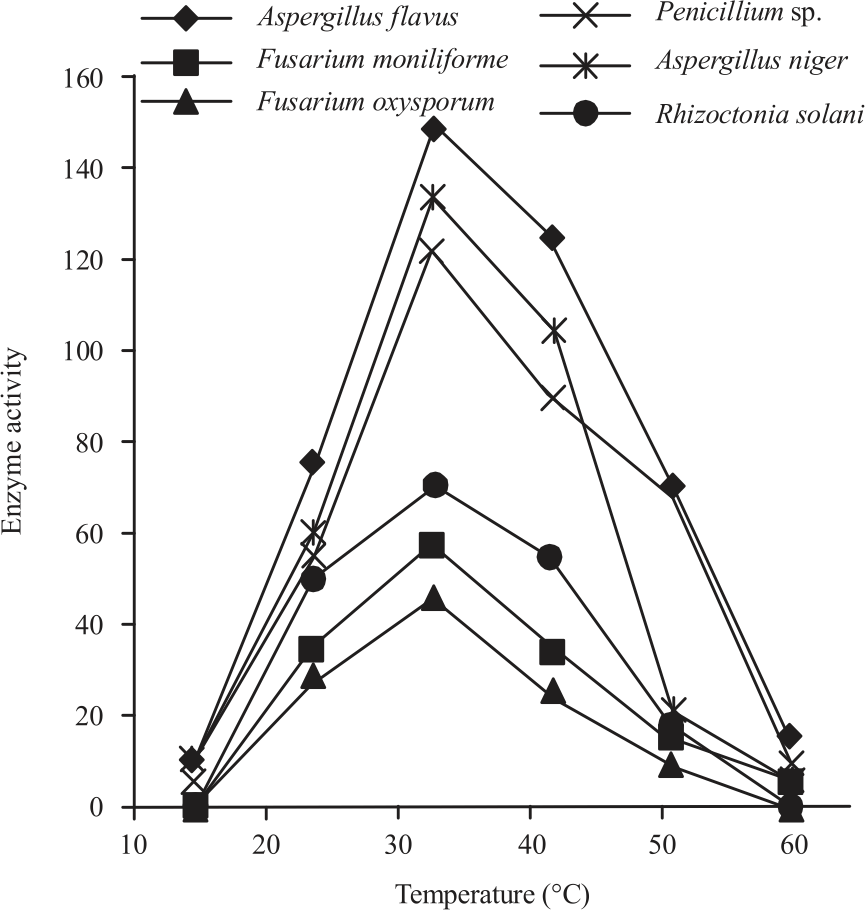 |
| Fig. 2: | Effect of different temperature on activity of amylase secreted by different 6 isolated fungi from the peanut seeds |
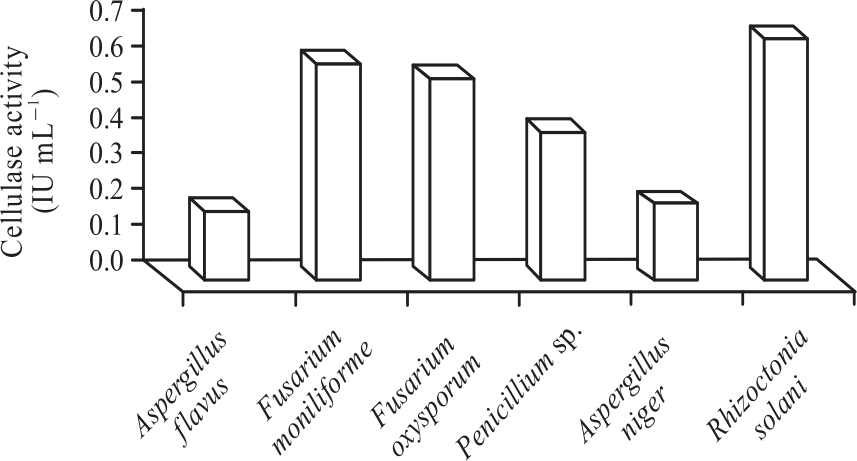 |
| Fig. 3: | Cellulase activity of most prevalent fungi isolated from peanut seeds collected from different 4 locations in Egypt |
Fusarium moniliforme and Fusarium oxysporum showed maximum activity at 30oC with a fast reduction in activity with increasing or decreasing temperatures. Enzymatic activity corresponds directly proportional to the proper temperature for growth24.
Cellulase activity: In this experiment mineral medium supplemented by sources of cellulose (CMC) with rate 2 g/1000 mL of medium as a solo source of carbon. Obtained data recorded in Fig. 3 showed that Rhizoctonia solani recorded maximum activity of cellulase enzyme, F. moniliforme and F. oxysporum occupied the second and third rank, respectively.
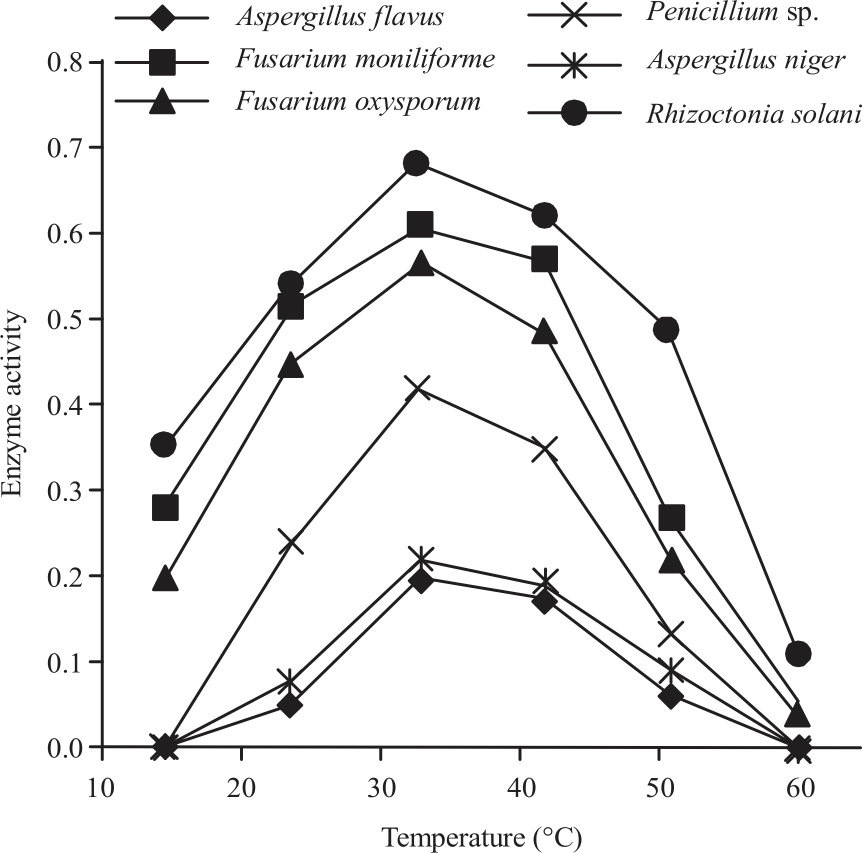 |
| Fig. 4: | Effect of different temperature on activity of cellulase secreted by different 6 fungi isolated from peanut |
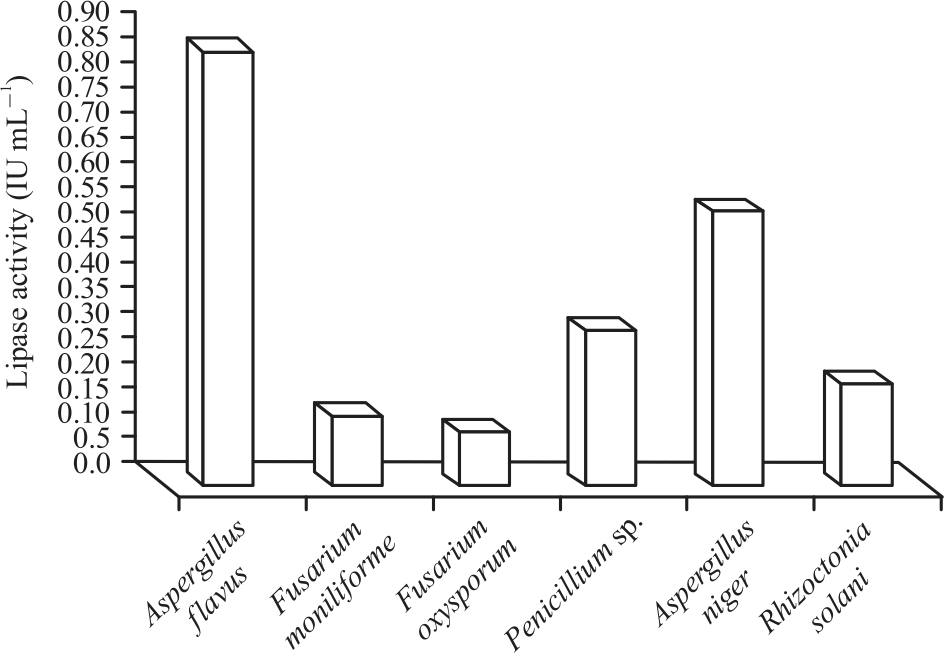 |
| Fig. 5: | Lipase activity of the most prevalent fungal isolates on peanut seeds collected from different four governorates in Egypt |
Cellulase enzyme secreted by A. flavus showed the lowest activity while Penicillium sp. and Aspergillus niger recorded medium level of cellulase enzyme activity. Cellulase activity produced by 5 isolated fungi was tested21. The result showed that the enzyme activity was superior of 2 isolates more than the other 3.
Effect of temperature on cellulase activity: Temperature is one of important factors which effect on enzyme activity. Results indicated that maximum cellulase activity of all tested fungi was recorded at 30°C with a difference in thermal stability between the tested fungi. Rhizoctonia solani characterized with a wide range of thermal stability ranged between 20-50°C.
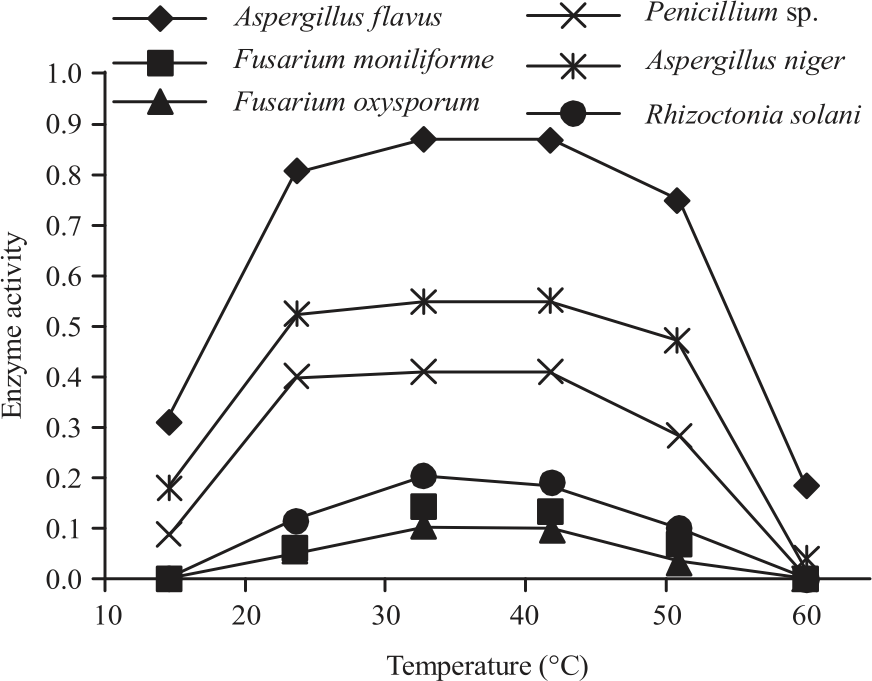 |
| Fig. 6: | Effect of different temperature on the lipase activity secreted by different 6 isolated fungi from peanut |
Also, thermal stability of Fusarium moniliforme and Fusarium oxysporum were between 20-40°C. Cellulase activity of Penicillium sp., Aspergillus niger and Aspergillus flavus recorded a noticeable reduction less than 30°C and more than 40°C. Effect of temperatures on enzymatic activity due to the impact of the effect of changes of temperature on the growth and cellular activity for each fungus. During the test of the effect of temperature on the cellulase activity, it was found that it reached maximum21 at a temperature of 30°C.
Lipase activity: The results of evaluation for the activity of crude lipase enzyme extracted by the 6 tested fungi illustrated in Fig. 4. Data showed that lipase activity secreted by Aspergillus flavus was in highest activity followed by that secreted by Aspergillus niger. Medium activity was recorded by Penicillium sp. and R. solani. Very low activity of lipase enzyme was obtained by F. monilifofrme and F. oxysporum. Lipase activity of all isolated fungi from peanut seeds varied in lipase activity25.
Effect of temperature on lipase activity: Effects of different temperatures on lipase activity of the 6 isolated fungi from peanut were tested. Data obtained in Fig. 6 showed lipolytic activity of all tested fungi were almost at the maximum between 30 and 40°C and differed in its stability. Lipolytic activity of Aspergillus flavus and Aspergillus niger were ranged from 20-50°C with the optimum between 30-40°C. Maximum of lipolytic activity of Penicillium sp. was between 20-40°C with sharp decrease at more or less than this range. While, the stability of the other 3 fungi was between 30-40°C with gradually decreasing at above or under previous range. Penicillium notatum showed significant maximum lipase activity at 20°C and followed by Fusarium sp. while, Penicillium chrysogenum showed maximum lipase production at 30°C as compared to other fungi25.
Lower and higher temperatures appear falling in the activities values. Lower temperature decreases the activity because it is unsuitable for fungal growth. Consequently, decreased enzyme production while higher temperature lead to minimize media water content by vaporization, as a result, it effects on cells growth. In addition, the higher temperature limit oxygen concentration which effect on fungal activity26.
All obtained data helped to give us some information about how these fungi cause infection and also the value of quality losses. Data proved that Aspergelus flavus has great activity of lipase which effect on quality and quantity of lipid contents, but has a neglected activity of cellulase which indicated that it has no ability of causing infection by itself while Aspergillus flavus, Aspergillus niger and Penicillium sp. have high activity of amylase which cause loss in carbohydrates contain. Thakare et al.27 mentioned that amylase, protease and lipase are responsible for solubilizing reserve food material in form of starch, protein and lipid, respectively in the seed. In contrast, cellulose activity obtained from R. solani sufficient enough to penetrate the host plant and causing infection by itself while has very low lipase activity to change the properties of lipids in seeds. Heiler et al.28 found that cellulase enzyme activity increased during appressorium development and reached a maximum when infection hyphae and haustorial mother cells were formed. Seven cellulolytic enzymes were separated by chromate focusing. While, the fungus caused the infection, the enzymatic activity reached to 3.3% during appressorium formation and increased to 36.6% when infection hyphae were formed and to 45.4% in haustorial mother cells.
CONCLUSION
Seventeen fungal species belonging to 14 genera were isolated from tested peanut seeds samples collected from 4 locations i.e., Beheira, Ismailia, Giza and Sohag governorates. Aspergillus flavus, Fusarium moniliforme, Fusarium oxysporum, Penicillium sp., Aspergillus niger and Rhizoctonia solani were the most frequently and most prevalent in all tested locations. Rhizoctonia solani characterized with maximum activity of cellulase enzyme while Aspergillus flavus recorded high activity of lipase enzyme. Temperature has different effect on enzymes stability along the 6 tested isolates. All enzymes secreted from all tested and Aspergillus niger has stability along wide range of temperature 20-50°C.
SIGNIFICANCE STATEMENT
This study gives researchers an idea about geographical distribution of seed-borne fungi for peanut crop in 4 governorates represent to the various climatic conditions in Egypt. Also, this study found link between development of infection and ability to secrete more efficient enzymes for example the fungus that has the ability to penetrate cellular walls rich in cellulose has the ability to secret cellulase enzyme like Rhizoctonia solani, F. moniliforme and F. oxysporum this information can be used in the control programs. This study will help the researcher to concentrate their research to restrict these genera that responsible for secreting this enzyme.
REFERENCES
- Bai, Y., A. Souleimanov and D.L. Smith, 2002. An inducible activator produced by a Serratia proteamaculans strain and its soybean growth-promoting activity under greenhouse conditions. J. Exp. Bot., 53: 1495-1502.
CrossRefDirect Link - Elwakil, M.A. and M.A. El-Metwally, 2001. Seed-borne fungi of peanut in Egypt: Pathogenicity and transmission. Pak. J. Biol. Sci., 4: 63-68.
CrossRefDirect Link - Janardhan, A., D. Subramanyam, A.P. Kumar, M.R. Pradeep and G. Narasimha, 2011. Aflatoxin impacts on germinating seeds. Ann. Biol. Res., 2: 180-188.
Direct Link - Pandey, A., P. Nigam, C.R. Soccol, V.T. Soccol, D. Singh and R. Mohan, 2000. Advances in microbial amylases. Biotechnol. Applied Biochem., 31: 135-152.
PubMedDirect Link - Van der Maarel, M.J.E.C., B. van der Veen, J.C.M. Uitdehaag, H. Leemhuis and L. Dijkhuizen, 2002. Properties and applications of starch-converting enzymes of the α-amylase family. J. Biotechnol., 94: 137-155.
CrossRefPubMedDirect Link - Cen, P. and L. Xia, 1999. Production of cellulase by solid-state fermentation. Adv. Biochem. Eng./Biotechnol., 65: 69-92.
CrossRefDirect Link - Mahadik, N.D., U.S. Puntambekar, K.B. Bastawde, J.M. Khire and D.V. Gokhale, 2002. Production of acidic lipase by Aspergillus niger in solid state fermentation. Process Biochem., 38: 715-721.
CrossRefDirect Link - Migheli, Q., V. Balmas, H. Harak, S. Sanna, B. Scherm, T. Aoki and K. O'Donnell, 2010. Molecular phylogenetic diversity of dermatologic and other human pathogenic fusarial isolates from hospitals in Northern and Central Italy. J. Clin. Microbiol., 48: 1076-1084.
CrossRefDirect Link - O'Donnell, K., E. Cigelnik and H.I. Nirenberg, 1998. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia, 90: 465-493.
CrossRefDirect Link - Visagie, C.M., J. Houbraken, J.C. Frisvad, S.B. Hong and C.H.W. Klaassen et al., 2014. Identification and nomenclature of the genus Penicillium. Stud. Mycol., 78: 343-371.
CrossRefDirect Link - McClenny, N., 2005. Laboratory detection and identification of Aspergillus species by microscopic observation and culture: The traditional approach. Med. Mycol., 43: S125-S128.
CrossRefDirect Link - McCabe, P.M., M.P. Gallacher and J.W. Deacon, 1999. Evidence for segregation of somatic incompatibility during hyphal tip subculture of Rhizoctonia solani AG 4. Mycol. Res., 103: 1323-1331.
CrossRefDirect Link - Ratnasri, P.V., B.K.M. Lakshmi, K. Ambika Devi and K.P.J. Hemalatha, 2014. Isolation, characterization of Aspergillus fumigatus and optimization of cultural conditions for amylase production. Int. J. Res. Eng. Technol., 3: 457-463.
CrossRefDirect Link - Kaur, H.P. and D. Joshi, 2015. Optimization of cellulase produced by fungus isolated from water. World J. Pharm. Pharmaceut. Sci., 4: 521-534.
Direct Link - Pualsa, J., D. Verma, R. Gavankar and R.D. Bhagat, 2013. Production of microbial lipases isolated from curd using waste oil as a substrate. Res. J. Pharmaceut. Biol. Chem. Sci., 4: 831-839.
Direct Link - Satyanagalakshmi, K., G. Sridhar, Renuka and D. Sunny, 2014. Screening and production optimisation of alpha amylase from Aspergillus strains by using solid state fermentation. Int. J. Curr. Microbiol. Applied Sci., 3: 623-631.
Direct Link - Krishnakumar, S., V.D.M. Bai and J. Premkumar, 2015. Production of alpha amylase by salt-tolerant actinomycete Streptomyces sp.-SBU3 isolated from marine sponge. Indian J. Geo-Mar. Sci., 44: 1-6.
Direct Link - Kakde, R.B. and A.M. Chavan, 2011. Extracellular lipase enzyme production by seed-borne fungi under the influence of physical factors. Int. J. Biol., 3: 94-100.
CrossRefDirect Link - Bedan, D.S., G.M. Aziz and A.J.R. Al-Sa'ady, 2014. Optimum conditions for α-amylase production by Aspergillus niger mutant isolate using solid state fermentation. Curr. Res. Microbiol. Biotechnol., 2: 450-456.
Direct Link - Thakare, H.S., V. Kumar and C. Singh, 2014. Effect of different hydrolytic enzymes on germination of inter and intra specific cotton hybrids and parents. Bioscan, 9: 943-946.
Direct Link - Heiler, S., K. Mendgen and H. Deising, 1993. Cellulolytic enzymes of the obligately biotrophic rust fungus Uromyces viciae-fabae are regulated differentiation-specifically. Mycol. Res., 97: 77-85.
CrossRefDirect Link