
Chukwuma Chukwuemeka Chukwuma
Department of Biochemistry, Faculty of Science, University of Port Harcourt, P.M.B. 5239, Choba, Rivers State, Nigeria
LiveDNA: 234.25603
Jude Chigozie Ikewuchi
Department of Biochemistry, Faculty of Science, University of Port Harcourt, P.M.B. 5239, Choba, Rivers State, Nigeria
Michael Okechukwu Monanu
Department of Biochemistry, Faculty of Science, University of Port Harcourt, P.M.B. 5239, Choba, Rivers State, Nigeria
Asian Journal of Applied Sciences
Year: 2019 | Volume: 12 | Issue: 4 | Page No.: 166-172







ABSTRACT
Background and Objective: Soil extracellular enzymes activities can reflect the metabolic need and nutrient availability of soil and as well possess the propensity to restore polluted soil towards normalcy. This study investigated the ability of some soil extracellular enzymes to recover crude oil polluted agricultural soil. Materials and Methods: Standard potentiometric method of pHKCl determination, loss of weight on ignition and culture methods were employed for all enzymatic and respiratory assays, organic matter and microbiological analyses, respectively. Results: Ninety days after planting, M. alternifolius and F. ferruginea treated soil groups recorded between 8.53 and 673.24% recovery as regards to dehydrogenase, protease, acid and alkaline phosphatase activities. Conclusion: Generally, the enzymes restored the polluted soil towards normalcy, probably necessitated by the treatment plants which aided in the abatement of the pollution and restoration of polluted soil.
PDF Abstract XML References Citation
Received: December 09, 2018;
Accepted: January 29, 2019;
Published: August 27, 2019
How to cite this article
Chukwuma Chukwuemeka Chukwuma, Jude Chigozie Ikewuchi and Michael Okechukwu Monanu, 2019. Enzymatic Recovery of Crude Oil Polluted Soil Enhanced by Treatment using Mariscus alternifolius Vahl. and Fimbristylis ferruginea. Asian Journal of Applied Sciences, 12: 166-172.
DOI: 10.3923/ajaps.2019.166.172
URL: https://scialert.net/abstract/?doi=ajaps.2019.166.172
DOI: 10.3923/ajaps.2019.166.172
URL: https://scialert.net/abstract/?doi=ajaps.2019.166.172
INTRODUCTION
Soil extracellular enzymes (SEES) are proximal drivers of decomposition1 and activities of these enzymes in soils can be altered by crude oil pollution. Soil enzyme activities are related to heavy metal contamination and reduced by 10-50 times with an increase in heavy metal concentration. This increase may lead to heavy metal toxicity thereby affecting the microbial density, diversity and activity2.
Soil micro-organisms produce extracellular enzymes to execute various biogeochemical processes involving several inorganic and redox reactions. Hydrolytic enzymes are needed for decomposition and mineralization of nutrients which are significant for the function of ecological unit3. In fertile soils, heterotrophic microbes are presented with detritus from plants and other biomasses that are rich in carbon4 and other nutrients essential for their cell growth and maintenance. However, micro-organisms cannot transport these large macro-molecules directly into their cytoplasm5. They require extracellular hydrolytic enzymes (EHES)4 to depolymerize them and generate soluble, low-number oligomers and monomers that are then recognized by cell-wall receptors and transported across the outer membrane and into the cell5.
Extracellular enzymes such as proteases, dehydrogenases and phosphatases are engrossed for the recycling of carbon, nitrogen and phosphorus6. The activities of these enzymes can unswervingly mirror metabolic need and the nutrients available to soil micro-organisms7.
Soil proteases (EC 3.4) are enzymes that perform proteolysis (protein catabolism) by hydrolyzing peptide bonds8. They are fundamental in nitrogen mineralization in a process which controls the quantity of plant available nitrogen9,10. Synthesis and activities of proteases in the soil are regulated by many factors such as climate, soil properties and the presence of organic compounds of plant and microbial basis11.
Soil dehydrogenases (EC 1.1.1) are the main representatives of the oxidoreductase enzymes class12. Amid all enzymes in the soil environment, dehydrogenases are one of the central and are used as a marker of overall soil microbial activity12,13. This is due to its intracellular occurrence in virtually all viable living microbial cells14,15 and function as a measurement of the the metabolic state of soil micro-organism16. It is one of the most adequate, vital and sensitive indicators relating to soil fertility17. The activities of dehydrogenases depend on the same factors, which influence the microbial abundance and activity18. They are closely related with microbial oxidoreduction processes14. Dehydrogenases play a momentous task in the biological oxidation of soil organic matter (SOM) by transferring hydrogen from organic substrates to inorganic acceptors19. However, the association between an individual biochemical property of soil dehydrogenases and the total microbial activity is not always apparent since microbial disintegration is highly diverse20.
The phosphatase enzyme (EC 3.1.1) is used to explain a wide group of enzymes that hydrolyze organic phosphorus compounds, pyrophosphates, inorganic polyphosphates and metaphosphates in soils. It is generally accepted that plants utilize only inorganic phosphorus and since a huge proportion of soil phosphorus is organically bound, the mineralization of this organic fraction can be vital in plant nutrition21. The C-O-P ester bonds present in soil organic phosphorus compounds are hydrolyzed by phosphatase. Phosphatase activity is, hence, a vital aspect in sustaining and controlling the rate at which phosphorus is recycled through soils22. This study was therefore carried out to determine the soil extracellular enzyme activities in crude oil polluted soils undergoing treatment using M. alternifolius and F. ferruginea.
MATERIALS AND METHODS
Experimental design: This study was carried out at the University of Port Harcourt Ecological Center for 10 months between May, 2017 and February, 2018.
Crude oil polluted agricultural soil obtained from a spill site located in Ogoniland, Nigeria was randomly collected using sterile plastic bags sealed with rubber bands. Likewise, unpolluted soil was collected from an agricultural farmland located in the University of Port Harcourt. The soil samples were transported to Ecological Centre of the University of Port Harcourt for pot experimental study. Viable and mature seeds of M. alternifolius and F. ferruginea, collected from the wild after identification of the species from the spill site were used for nursery and afterward transplanted into polluted soil samples for treatment. Soil enzymatic, respiratory and microbial activities were determined in the soil samples prior to the commencement of the treatment and subsequently every 30 days over a 90 day period. The soil extracellular enzymes and respiratory activities and microbial density in crude oil polluted soils undergoing treatment were ascertained.
Soil enzymatic and soil microbial respiratory activities: The Alef and Nannipieri23 potentiometric method of pHKCl determination as adapted by Chukwuma et al.2 was employed for all enzyme assays while substrate induced method was employed for respiratory assays.
Protease activities: Protease activities were determined based on amino acids released after incubation of soil with sodium caseinate for 2 h at 50°C. The supernatant containing 1.4 M NaSO4 and Folin-Ciocalteu (diluted 3 times) was read at 578 nm.
Dehydrogenase activities: Dehydrogenase activities were determined by estimating the rate of reduction of 2, 3, 5- triphenyl tetrazolium chloride (TTC) to triphenyl formazan (TPF) after incubation at 30°C for 24 h. Aliquots of the supernatants were thereafter read at 485 nm.
Acid and alkaline phosphatase activities: Acid and alkaline phosphatase activities assay relied on p-nitrophenol phosphate (PNP) determination for 1 h at 37°C. The supernatants containing aliquots of 0.5 M CaCl2 and 0.5 M NaOH were read at 485 nm.
Respiratory activity: The substrate (glucose) induced method was employed for soil microbial respiratory activity estimation where screened (2 mm sieve) soil sample was introduced into the respiratory flask in addition to a glucose solution. After 24 h at 25°C, the respiratory activity was determined by titrating the CO2 captured using NaOH with HCl and phenolphthalein indicator.
Soil microbial analyses: Total Culturable Heterotrophic Bacteria Count (TCHBC) and Total Heterotrophic Fungi (THF) TCHBC and TFC were determined by spread plate on nutrient agar (NA) and potato dextrose agar (PDA) methods respectively, as well as plate count agar (PCA) method as adopted by Seeley and VanDemark24 and Ogunmwonyi et al.25. Following decimal dilutions (5 fold) of soil suspensions, TCHBC were plated out on NA medium and incubated at 30°C for 24 h while fungal isolates were plated out on PDA medium and incubated for 3 days at 28±2°C.
Physicochemical analysis
Soil Organic Matter (SOM): Organic matter was determined by loss of weight on ignition method as reported by Motsara and Roy26 where sieved (2 mm) soil was dried at 105°C for 4 h, weighed, re-dried at 400°C and reweighed.
Statistical analysis: Results are expressed as means±standard deviation of triplicate determination. One way analysis of variance was employed to determine the significant difference between the groups. Data between groups were ascertained by the Bonferroni test using Statistical Package for the Social Science (SPSS®) Version 20 statistics software at 95% (p = 0.05) confidence level, while data between periods were ascertained by the Student t-test.
RESULTS
Protease activities: Protease activities of the various treated soil groups as shown in Table 1 revealed an increase in the absolute values of M. alternifolius treated soil over time, while those of F. ferruginea treated soil increased 30 DAP but subsequently decreased 60 and 90 DAP.
Dehydrogenase activities: The dehydrogenase activities (Table 2) of both treated soil groups significantly (p<0.05) increased 30 and 60 DAP albeit no significant difference (p<0.05) was recorded 90 DAP.
Acid phosphatase activities: Acid phosphatase activities of the treated soil groups are shown in Table 3. Results revealed that 90 DAP, a significant (p<0.05) increase in acid phosphatase activities of M. alternifolius treated soil was recorded while a significant (p<0.05) decrease in activities was recorded in F. ferruginea treated.
Alkaline phosphatase activities: Alkaline phosphatase activities are presented in Table 4. There was no significant difference (p<0.05) recorded in the alkaline phosphatase activities of M. alternifolius and F. ferruginea treated soil groups.
Soil respiratory activities: The soil respiratory activities of the treated soil groups are presented in Table 5. There was a significant (p<0.05) decrease in soil respiratory activities of M. alternifolius treated soil group while no significant difference (p<0.05) was recorded in F. ferruginea treated soil, 90 DAP.
Soil organic matter (SOM): The result of soil organic matter content (SOM) of the treated soil groups as presented in Table 6. There was a significant (p<0.05) decrease in SOM over time.
Total culturable heterotrophic bacteria count (TCHBC): The results of total heterotrophic bacteria count of the treated soil groups are presented in Table 7. There was a significant (p<0.05) increase in the TCHBC over time.
| Table 1: | Protease activities (tyrosine mg kg–1 dry matter h–1) of the soil groups |
 | |
Values are Mean±standard deviations of triplicate determinations, values in the same column with different letters (a,b) are significantly different at p<0.05, *p<0.05 compared to the corresponding values before planting, BP: Before planting, DAP: Day(s) after planting, NA: Not applicable | |
| Table 2: | Dehydrogenase activities (TPF mg kg–1/day) of the soil groups |
 | |
Values are Mean±standard deviations of triplicate determinations, values in the same column with different letters (a,b) are significantly different at p<0.05, *p<0.05 compared to the corresponding values before planting, BP: Before planting, DAP: Day(s) after planting, NA: Not applicable | |
| Table 3: | Acid phosphatase activities (PNP mmol kg–1 dw h–1) of the soil groups |
 | |
Values are Mean±standard deviations of triplicate determinations, values in the same column with different letters (a,b,c) are significantly different at p<0.05, *p<0.05 compared to the corresponding values before planting, BP: Before planting, DAP: Day(s) After planting, NA: Not Applicable | |
| Table 4: | Alkaline phosphatase activities (PNP mmol kg–1 dw h–1) of the soil groups |
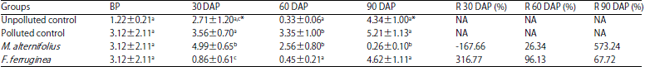 | |
Values are Mean±standard deviations of triplicate determinations, values in the same column with different letters (a,b,c) are significantly different at p<0.05, *p<0.05 compared to the corresponding values before planting, BP: Before planting, DAP: Day(s) after planting, NA: Not applicable | |
| Table 5: | Soil microbial respiratory activities (mg) of the soil groups |
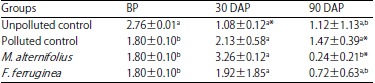 | |
Values are Mean±standard deviations of triplicate determinations, values in the same column with different letters (a,b) are significantly different at p<0.05, *p<0.05 compared to the corresponding values before planting, BP: Before planting, DAP: Day(s) after planting | |
| Table 6: | Soil Organic matter (SOM) content (%) of the soil groups |
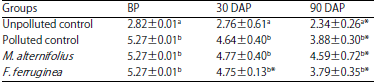 | |
Values are Mean±standard deviations of triplicate determinations, values in the same column with different letters (a,b) are significantly different at p<0.05, *p<0.05 compared to the corresponding values before planting, BP: Before planting, DAP: Day(s) after planting | |
Total fungi count (TFC): The result of total fungi count as shown in Table 8 revealed significant (p<0.05) increase in the HUF over time.
| Table 7: | Total culturable heterotrophic bacteria count (TCHBC) (in Log10 CFU g–1) of the soil groups |
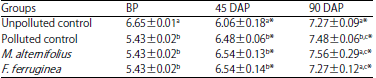 | |
Values are Mean±standard deviations of triplicate determinations, values in the same column with different letters (a,b,c) are significantly different at p<0.05, *p<0.05 compared to the corresponding values before planting, BP: Before planting, DAP: Day(s) after planting | |
| Table 8: | Total fungi count (TFC) (in Log10 CFU g–1) of the soil groups |
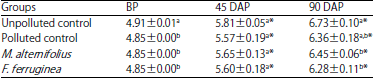 | |
Values are Mean±standard deviations of triplicate determinations, values in the same column with different letters (a,b) are significantly different at p<0.05, *p<0.05 compared to the corresponding values before planting, BP: Before planting, DAP: Day(s) after planting | |
Pearson’s correlation coefficient (PCC) of observed enzyme activities versus soil organic matter (SOM): Statistical analysis has shown a trend between organic matter, enzyme and respiratory activities (Table 9).
| Table 9: | Pearson’s correlation coefficient (PCC) of observed enzyme activities versus soil organic matter (SOM) content of the soil groups |
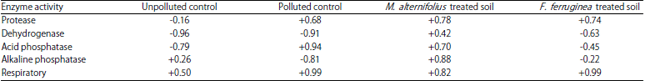 | |
| *Correlation is significant at the 0.05 level (2-tailed) | |
Pearson’s correlation coefficient (PCC, a measure of the linear correlation or dependence between two variables) of +0.68, +0.78, +0.74 (Table 9) for protease activity in polluted control, M. alternifolius treated and F. ferruginea treated soils respectively, showed a substantial positive correlation with soil organic matter while unpolluted control showed a negative correlation (-0.16 PCC) with soil organic matter. The considerable negative correlation of -0.96, -0.91 and -0.63 for dehydrogenase activity and SOM was recorded in unpolluted control, polluted control and F. ferruginea treated soil respectively, while a fair positive correlation (+0.42 PCC) was observed in M. alternifolius treated soil. Acid phosphatase activity showed a substantial positive correlation with soil organic matter for polluted control and M. alternifolius treated soil (+0.94 and +0.70 PCC, respectively) and a fair positive correlation (+0.45 PCC) for F. ferruginea treated soil while unpolluted control showed a substantial negative correlation (-0.79 PCC). On the other hand, alkaline phosphatase activity revealed a substantial negative (-0.81 PCC) and positive correlation (+0.88 PCC) with SOM for polluted control and M. alternifolius treated soil respectively, while a fair positive (+0.26 PCC) and negative (-0.22 PCC) correlation was observed for unpolluted control and F. ferruginea treated soil, respectively. Soil microbial respiratory activity showed a substantial positive correlation (+0.50, +0.99, +0.82 and +0.99 PCC) with soil organic matter for unpolluted control, polluted control, M. alternifolius and F. ferruginea treated soils.
DISCUSSION
The observed increase in protease activities over time as recorded in M. alternifolius treated soil and 30 DAP for F. ferruginea treated soil may be due to growth in microbial population, while the sharp decrease in activities may have been necessitated by the inhibitory influence of the treatment plant on micro-organisms-producing proteases. The latter corresponds with the findings of Chukwuma et al.2 who reported a decrease in protease activities of soils phytoremediated over a 12 week period. This assertion may be supported by Adeniyi et al.27 who reported the tendency of plant extracts to inhibit certain micro-organisms. The percentage recovery given by:
![]()
Chukwuma et al.2 showed that by 30 DAP, treatment using M. alternifolius restored the polluted soils towards normal value (60.78%). However, the value obtained for treatment using F. ferruginea nose-dived, indicating the failure in restoration, especially with regards to the activities of proteases. By 60 DAP, only treatment using M. alternifolius restored the polluted soil towards normal value (11.88%). Likewise, 90 DAP, treatments using M. alternifolius and F. ferruginea restored the polluted soils towards normalcy (121.65 and 8.53).
The significant (p<0.05) increase in dehydrogenase activities as recorded for both treated soil groups corresponds with previous report2. Although the microbial populations of the treated soil groups increased over time, Nath and Samanta28 has indicated that aside the microbial population, activities of these microbes and the kind of microbes present in the soil determine the activities of enzymes. Nevertheless, it may be that the ecological equilibrium of micro-organisms was disrupted which led to the decrease in production of the enzyme. By 30 DAP, recovery of M. alternifolius and F. ferruginea treated groups nose-dived demonstrating failure in restoration particularly with regards to the activities of dehydrogenases. However, by 60 DAP treatment using F. ferruginea restored the polluted soil towards normal value (1.79%) albeit the value obtained for treatment using M. alternifolius nose-dived. Likewise, by 90 DAP, only treatments using M. alternifolius restored the polluted soils towards normalcy (88.26%), while values obtained for treatments using F. ferruginea nose-dived.
The significant (p<0.05) decrease in acid phosphatase activities recorded in F. ferruginea treated soil group may be attributed to the redundancy in microbial population and/or activity 90 DAP and this corroborates with earlier report2 which in addition associated such a decrease to limiting effects of nutrients in the pots posed by its depletion over time. As previously reported29, such activities as depicted by the groups may indicate the state of the soil in terms of physicochemical properties, especially pH, which is a key determinant of soil acid and alkaline phosphatase activities. Nonetheless, the treatments restored the soil towards normal levels by 30, 60 and 90 DAP, even though treatment using M. alternifolius by 30 DAP nosedived indicating a failure in restoration.
The significant (p<0.05) decrease in soil respiration observed 90 DAP M. alternifolius corroborates the report30 which previously associated such a decline in respiratory activity after attaining peak to the depletion of available carbon substrates.
The significant (p<0.05) decrease in SOM over time as recorded in the study corroborates the report31 which associated such a decrease in the degradation of crude oil Nonetheless, it may, however, be an indication of microbial activities ongoing in the soil. Microbes consume organic matter and in turn release CO2. Therefore, a decrease in SOM might be indicated high soil microbial and respiratory activity. This aligned with the report32 that the decomposition of organic matter is largely a biological process that occurs naturally and determined by soil organisms, the physical environment and the quality of the organic matter.
The increase in TCHBC and TFC of the treated soil groups over was not surprising as similar finding32 has been reported which shows the progressive utilization of organic matter and hydrocarbon, with the hydrocarbons functioning as primary substrates. Ebuehi et al.33, such increase is indicative of increased biodegradation by the microbial community.
CONCLUSION
Microbial and enzymatic activities can reflect the metabolic need and nutrient availability of soil. The soil extracellular enzymes were found to restore the polluted soil towards normalcy, probably necessitated by the type of plant involved in the treatment. Therefore, it is noteworthy to state that the application of these plant species in the treatment of crude oil polluted agricultural soils can enhance microbial and enzyme activities thereby aiding in the abatement of the pollution and restoration of the polluted soil.
SIGNIFICANCE STATEMENT
This study discovered the influence of M. alternifolius and F. ferruginea plants species in the enzymatic recovery of crude oil polluted agricultural soil over time. This study will help researchers to uncover the critical areas of soil recovery using soil extracellular enzymes.
REFERENCES
- Kivlin, S.N. and K.K. Treseder, 2014. Soil extracellular enzyme activities correspond with abiotic factors more than fungal community composition. Biogeochemistry, 117: 23-37.
CrossRefDirect Link - Chukwuma, C.C., M.O. Monanu, J.C. Ikewuchi and C. Ekeke, 2018. Variance in protease, dehydrogenase, phosphatase and respiratory activities during phytoremediation of crude oil polluted agricultural soil using Schwenkia americana L. and Spermacoce ocymoides Burm. f. Annu. Res. Rev. Biol., 28: 1-9.
CrossRef - Bell, C.W., B.E. Fricks, J.D. Rocca, J.M. Steinweg, S.K. McMahon and M.D. Wallenstein, 2013. High-throughput fluorometric measurement of potential soil extracellular enzyme activities. J. Visual. Exp., Vol. 81.
Direct Link - German, D.P., K.R. Marcelo, M.M. Stone and S.D. Allison, 2012. The Michaelis-M enten kinetics of soil extracellular enzymes in response to temperature: A cross‐latitudinal study. Global Change Biol., 18: 1468-1479.
CrossRefDirect Link - Yuan, B.C. and D.X. Yue, 2012. Soil microbial and enzymatic activities across a chronosequence of chinese pine plantation development on the loess plateau of China. Pedosphere, 22: 1-12.
CrossRefDirect Link - Yang, M., D. Yang and X. Yu, 2018. Soil microbial communities and enzyme activities in sea-buckthorn (Hippophae rhamnoides) plantation at different ages. Plos One, Vol. 13.
CrossRefDirect Link - Kamal, S., S. Rehman and H. Iqbal, 2017. Biotechnological valorization of proteases: From hyperproduction to industrial exploitation-A review. Environ. Prog. Sustain. Energy, 36: 511-522.
CrossRefDirect Link - Vranova, V., K. Rejsek and P. Formanek, 2013. Proteolytic activity in soil: A review. Applied Soil Ecol., 70: 23-32.
CrossRefDirect Link - Gu, Y., P. Wang and C.H. Kong, 2009. Urease, invertase, dehydrogenase and polyphenoloxidase activities in paddy soil influenced by allelopathic rice variety. Eur. J. Soil Biol., 45: 436-441.
CrossRefDirect Link - Salazar, S., L.E. Sanchez, J. Alvarez, A. Valverde and P. Galindo et al., 2011. Correlation among soil enzyme activities under different forest system management practices. Ecol. Eng., 37: 1123-1131.
CrossRefDirect Link - Moeskops, B. Sukristiyonubowo, D. Buchan, S. Sleutel and L. Herawaty et al., 2010. Soil microbial communities and activities under intensive organic and conventional vegetable farming in West Java, Indonesia. Applied Soil Ecol., 45: 112-120.
CrossRefDirect Link - Zhao, B., J. Chen, J. Zhang and S. Qin, 2010. Soil microbial biomass and activity response to repeated drying-rewetting cycles along a soil fertility gradient modified by long-term fertilization management practices. Geoderma, 160: 218-224.
CrossRefDirect Link - Watts, D.B., H.A. Torbert, Y. Feng and S.A. Prior, 2010. Soil microbial community dynamics as influenced by composted dairy manure, soil properties and landscape position. Soil Sci., 175: 474-486.
CrossRefDirect Link - Jarvan, M., L. Edesi, A. Adamson and T. Vosa, 2014. Soil microbial communities and dehydrogenase activity depending on farming systems. Plant Soil Environ, 60: 459-463.
Direct Link - Zhang, N., X.D. He, Y.B. Gao, Y.H. Li and H.T. Wang et al., 2010. Pedogenic carbonate and soil dehydrogenase activity in response to soil organic matter in Artemisia ordosica community. Pedosphere, 20: 229-235.
CrossRefDirect Link - Subhani, A., H. Changyong, X. Zhengmiao, L. Min and A.M. El-Ghamry, 2001. Impact of soil environment and agronomic practices on microbial/dehydrogenase enzyme activity in soil. A review. Pak. J. Biol. Sci., 4: 333-338.
CrossRefDirect Link - Chen, H., Y. Li, D. Chen, Y. Zhang, L. Wu and J. Ji, 1996. Soil phosphorus fractions and their availability in Chinese fir plantations in South China. For. Res., 19: 121-126.
Direct Link - Seeley, H.W. and P.J. VanDemark, 1981. Microbes in Action. A Laboratory Manual of Microbiology. 3rd Edn., W.H. Freeman and Company, USA., Page: 350.
Direct Link - Ogunmwonyi, I.N., O.E. Igbinosa, O.A. Aiyegoro and E.E. Odjadjare, 2008. Microbial analysis of different top soil samples of selected site in Obafemi Awolowo University, Nigeria. Scient. Res. Essays, 3: 120-124.
Direct Link - Motsara, M.R. and R.N. Roy, 2008. Guide to Laboratory Establishment for Plant Nutrient Analysis. 1st Ed. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN: 9789251059814, Pages: 204.
Direct Link - Adeniyi, T.A., P.A. Adeonipekun and E.A. Omotayo, 2014. Investigating the phytochemicals and antimicrobial properties of three sedge (Cyperaceae) species. Notulae Scient. Biol., 6: 276-281.
Direct Link - Nath, R. and R. Samanta, 2012. Soil pH, microbial population, nitrate reductase and alkaline phosphatase activities of different environment of Dibrugarh district, Assam. Adv. Applied Sci. Res., 3: 1772-1775.
Direct Link - Okolo, J.C., E.N. Amadi and C.T.I. Odu, 2005. Effects of soil treatments containing poultry manure on crude oil degradation in a sandy loam soil. Applied Ecol. Environ. Res., 3: 47-53.
Direct Link - Ebuehi, O.A.T., I.B. Abibo, P.D. Shekwolo, K.I. Sigismund, A. Adoki and I.C. Okoro, 2005. Remediation of crude oil contaminated soil by enhanced natural attenuation technique. J. Applied Sci. Environ. Manage., 9: 103-106.
Direct Link