
Ndeye Penda Ndiaye
Department of Animal Biology, Faculty of Sciences and Techniques, Cheikh Anta Diop University of Dakar, P.O. Box 5005, Dakar, Fann, Senegal
LiveDNA: 221.25770
Adama Sow
Laboratory of Endocrinology and Radio-Immunology, Inter State School of Veterinary Sciences and Medicine of Dakar, P.O. Box 5077, Dakar, Fann, Senegal
Germain Jerome Sawadogo
Laboratory of Endocrinology and Radio-Immunology, Inter State School of Veterinary Sciences and Medicine of Dakar, P.O. Box 5077, Dakar, Fann, Senegal
Mbacke Sembene
Department of Animal Biology, Faculty of Sciences and Techniques, Cheikh Anta Diop University of Dakar, P.O. Box 5005, Dakar, Fann, Senegal
Asian Journal of Applied Sciences
Year: 2019 | Volume: 12 | Issue: 4 | Page No.: 149-157







ABSTRACT
Background and Objective: Improvement and conservation strategies of animal genetic resources require first of all, a full knowledge of breeds’ diversity and genetic structure. In order to disclose the current genetic structure of Senegalese local cattle breeds, this study has used the population genetics approaches. Materials and Methods: The DNA samples were taken from 120 unrelated cattle animals sampled from the main livestock breeding areas in Senegal. Genotyping was carried out using 11 microsatellite loci selected within the panel recommended by ISAG-FAO for cattle breeds’ genetic characterization. Genetic differentiation and structure analysis were performed using bioinformatics’ softwares. Results: The FST, measure of population differentiation indicated a mean value of 0.047±0.011, (p<0.05), followed with a mean gene flow (Nm) of 5.06. The Bayesian analysis indicated three clusters. The first one grouped mainly the N’Dama cattle, the second the majority of Djakore “hybrid” population and the last one clustered the zebu’s populations. Conclusion: Population structure analysis has revealed at present that Senegalese local cattle breeds were not running towards genetic erosion, nevertheless they deserve to be closely monitored to avoid the disappearance of important adaptive traits.
PDF Abstract XML References Citation
Received: December 10, 2018;
Accepted: February 01, 2019;
Published: August 27, 2019
How to cite this article
Ndeye Penda Ndiaye, Adama Sow, Germain Jerome Sawadogo and Mbacke Sembene, 2019. Diversity and Genetic Structure of Senegalese Local Cattle Breeds. Asian Journal of Applied Sciences, 12: 149-157.
DOI: 10.3923/ajaps.2019.149.157
URL: https://scialert.net/abstract/?doi=ajaps.2019.149.157
DOI: 10.3923/ajaps.2019.149.157
URL: https://scialert.net/abstract/?doi=ajaps.2019.149.157
INTRODUCTION
Animal genetic resources are playing a vital role in livestock production and are essential to a sustainable agriculture and food security1,2. The efforts of conservation of animal genetic resources in developing countries are minimal. The conservation programs that had been launched were running without a full spectrum of conservation strategies1. With the advent and development of artificial insemination during the last 50 years, the replacement of locally adapted breeds by a narrow range of high-yielding international trans-boundary breeds, is becoming an area of concern affecting animal genetic diversity in developing countries3,4. This development has led to mounting concerns about the genetic erosion5. Consequently, livestock breeds are threatened by the devastating effects of genetic erosion, the most significant of which are the dilution of genetic pool and the disappearance of important adaptive traits1.
In Africa, several studies have been conducted on cattle breeds for improvement and conservation purposes6-9. From these studies, all management strategies are elaborated on the basis of genetic diversity maintaining for better livestock productivity. Gobra zebu, Maure zebu and N’Dama cattle have been the subject of population genetics analysis among studies conducted at a continental scale to retrace indicine and taurine migration across Africa10-12. Ndiaye et al.13 reevaluated the genetic diversity and the phylogenetic relationships of Senegalese local cattle breeds including the Djakore cattle which have not been characterized up to now at molecular level. Importantly, the levels of genetic diversity of these cattle breeds have considerably increased under indiscriminate crossbreeding factors and changes in livestock breeding systems13. This might be attributed to inbreeding among the populations or the occurrence of population substructure. Following these facts, it appears that Senegalese local cattle have undergone significant modifications in their genetic pool. Therefore, it was crucial to know the changes occurred in the genetic structure of these populations regarding their breeding production environments whose factors influenced directly their fitness traits. In the context of breed improvement and conservation purposes, this study aimed at determining the current genetic structure of Senegalese local cattle by using population genetics approaches.
MATERIALS AND METHODS
Sample collection: The survey was carried out from October-December, 2013 in three agro-pastoral regions of Senegal: Saint-Louis, Kaolack and Kolda. These regions represent the main livestock breeding areas of Gobra zebu, Maure zebu, Djakore and N’Dama cattle. The sampled population consisted of 120 unrelated cattle animals selected from 15 different localities and the Zootechnical Research Center of Kolda. As no parentage records were available, so to ensure that animals were unrelated and held standard characters of the targeted population, it used the knowledge and experience of farmers and shepherds via questionnaire and interviews whenever possible.
Blood samples were collected from jugular vein into 4 mL vacutainer tubes containing the disodium salt of ethylene diamine tetra-acetate (EDTA) as anti-coagulant and stored ±4°C until DNA extraction. During the blood sampling, adequate measures were taken to minimize pain and discomfort in animals. The Ethics Committee of Cheikh Anta Diop University of Dakar approved this study. Participants’ approval was obtained after the study was fully explained.
Microsatellite loci genotyping: Genomic DNA was isolated from whole blood using Gentra Puregene Blood kit standard protocol developed by QIAGEN® group. The simplex PCR amplifications of 11 ISAG/FAO microsatellite loci were carried out in a 15 μL reaction volume. The PCR products were labelled with fluorescent dyes and genotyped using Li-Cor® automated sequencer (DNA Analyzer Model 4300) following the manufacturer’s procedures in genotyping lab of CIRDES (www.cirdes.org). After the migration profiles analysis by using SAGAGT GENERATION 2.0 software, a data set of 1320 genotypes was generated to perform genetic analysis (Table S1).
Genetic analysis: The distribution of genetic diversity among the four cattle populations was assessed by analyzing the F-statistics of Weir and Cockerham14 (FST (θ) and FIT (F)) and the gene flow (Nm) of Wright15, by using16 GENETIX ver. 4.05.2 and17 FSTAT ver. 2.9.3.2. Pairwise FST values between populations were computed using GENETIX by permutation method.
The factorial correspondence analysis from allele frequencies was performed using GENETIX to decipher the population structure. The genetic assignment tests were done using “simulation-exclusion” approach implemented18 in GENECLASS program ver. 2.0 and proposed by Cornuet et al.19. The “exclusion-simulation” tests were carried out using the allele frequencies method of Paetkau et al.20 among all cattle populations (mean FST = 0.047) between the most differentiated pair of populations (Djakore and N’Dama: FST = 0.107) and between the least differentiated pair of populations (Gobra zebu and Maure zebu: FST = 0.003).
To search the occurrence of independent genetic clusters (K) and to assign correctly individuals to populations where their genotype had the greatest belonging likelihood according to the ancestry proportion (Q), an alternative Bayesian clustering procedure implemented21 in STRUCTURE ver. 2.3.4. was used.
| Table S1: | Microsatellite genotypes of four Senegalese local cattle populations |
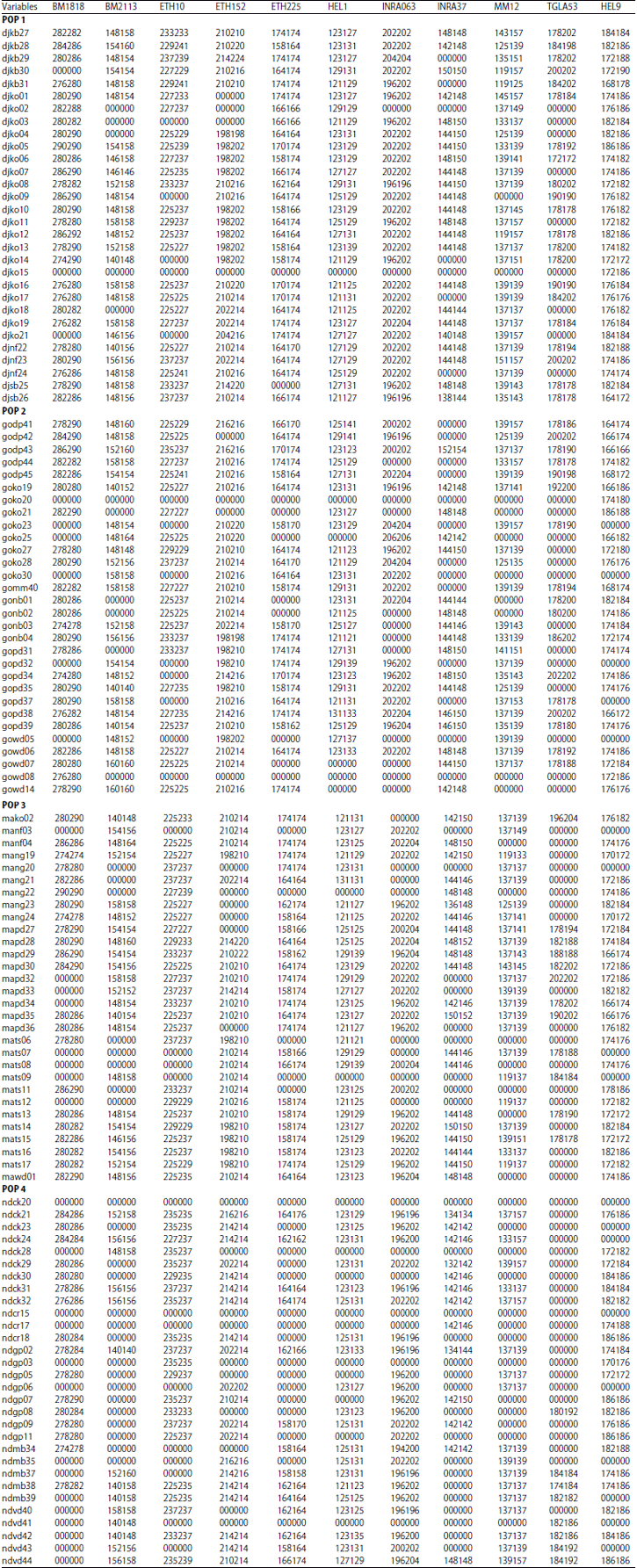 | |
Each population constituted of 30 individuals corresponding to a total of 1320 genotypes, dj: Djakore, go: Gobra zebu, ma: Maure zebu, nd: N’Dama, Pop: Population | |
Simulations were performed using a burn-in period of 10,000 iterations followed by 1,000,000 iterations of the Markov chain Monte Carlo algorithm. Independent runs of K were performed from 1-8 clusters and were repeated 20 times to check the reliability of results. To determine the most appropriate number of clusters (K) that best fit the data, the results generated by STRUCTURE have been submitted to STRUCTURE HARVESTER web site program22, which implemented the mean esteemed log-likelihood of K, L (K) and the Evanno method23.
RESULTS
Population differentiation analysis: Results of F-statistics measures were presented in Table 1. Considering all cattle populations, a global deficit of heterozygotes (FIT) of 11.7% (p<0.05) was observed. The multi-locus FST measure of differentiation indicated that 4.7% (p<0.05) of the total genetic diversity were due to unique allelic differences among populations. The mean gene flow (Nm) between populations was valued to 5.06 (Table 1).
The pairwise FST values between populations indicated that Gobra zebu and Maure zebu were the least differentiated populations (0.003), even so the highest pairwise FST value was observed between Djakore and N’Dama cattle (0.107) (p<0.05) (Table 2). Considering the esteemed Nm, the highest value was obtained between zebu’s populations (68.81), whereas the lowest value was found between N’Dama and Djakore (2.08) (Table 2).
Population structure analysis: Factorial Correspondence Analysis (FCA) revealed that the first axis with 59.03% of the total genetic variability separated properly N’Dama from the other populations (Fig. 1). Whereas, the second axis that contributed to 21.56% of the total variability showed that Djakore population was genetically closer to zebu’s populations and more especially to Gobra zebu.
The assignment tests based on allele frequencies method revealed that 50% of the cattle individuals were correctly assigned to the population they belong to with the exclusion probabilities of 1 and 0.1% (Table 3).
| Table 1: | Global F-statistics measured from the 11 microsatellite loci considering four cattle populations |
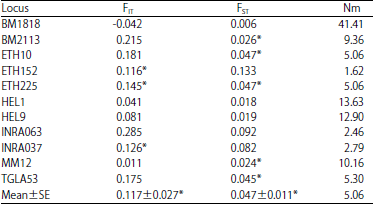 | |
| *p<0.05 = significant | |
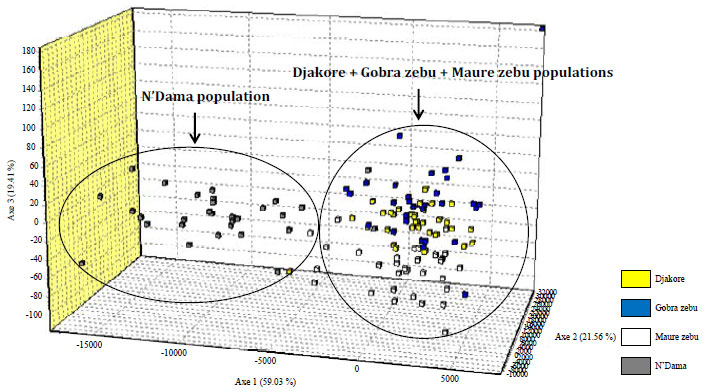 | |
| Fig. 1: | Factorial correspondence analysis of cattle individuals’ genotypes |
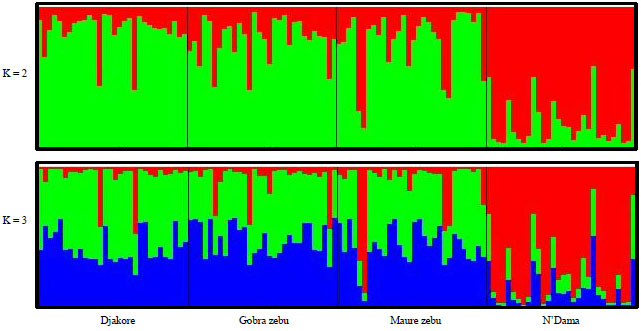 | |
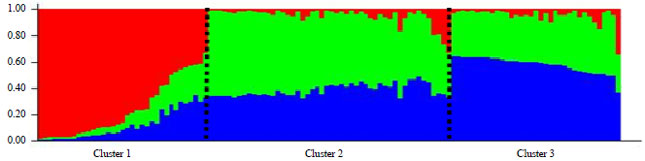
| Fig. 2: | Genetic structure of four Senegalese local cattle populations |
Each cattle individual is represented by a thin vertical bar, often partitioned into colored segments with the length of each segment representing the proportion of individuals’ genotypes from K = 2, 3. Thin black lines separated cattle populations | |
 | |
| Fig. 3: | Assignment of cattle individuals to 3 different genetic clusters inferred from the genotype membership coefficient (Q) |
| The dashed lines delimited the genetic clusters | |
| Table 2: | Pairwise FST and Nm estimates between pair of populations |
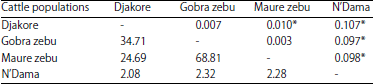 | |
The FST values are above the diagonal and those of the Nm below. *Values corresponded to significant tests (the repeating percentages of FST values lower than those observed, i.e., >95%) | |
Djakore and N’Dama cattle appeared as the most homogeneous populations because of 53.3 and 71.4% of their individuals were correctly assigned to the most critical threshold (p<0.001), respectively. Among zebu’s populations, the percentage of individuals correctly assigned was 43.3 and 16.6%, respectively. These percentages remained significantly lower than 50%.
The results given by the Bayesian clustering approach, indicated that the likeliest number of clusters considering all cattle populations was obtained for K = 3 (Fig. 2). Only the N’Dama individuals were grouped in their own cluster. However, the zebu’s populations and Djakore were assigned into two clusters (Fig. 3). Esteemed membership percentage of N’Dama individuals to cluster 1 was 76.8% (Table 4) and therefore, it was widely constituted of the N’Dama genotype. In the genetic clusters 2 and 3, Djakore and Gobra zebu cattle were mainly assigned with significant percentages of 51.3 and 49%, respectively. Maure zebu individuals were found unevenly distributed amongst the three genetic clusters with 12.3, 44.1 and 43%, respectively.
| Table 3: | Genetic assignment of individuals to their original population using the “exclusion-simulation” approach |
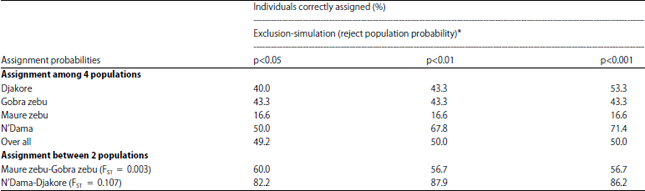 | |
*With the “exclusion-simulation” approach, an individual is considered as correctly assigned to a population when it is excluded from all excepting his original population (probability of belong to a population under the threshold p<0.05, 0.01 or 0.001) | |
| Table 4: | Percentages of individuals correctly assigned to the three inferred clusters |
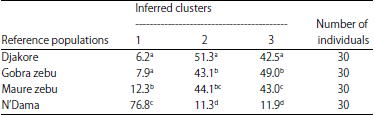 | |
| a,b,c,dPercentages of assignment within a column with no common superscripts differed significantly (p<0.001) | |
DISCUSSION
The mean FST value has shown a moderate and significant genetic differentiation among populations that could be attributed to population origin24 and geographic distribution25. Compared to these results, the same mean FST value (0.047) was obtained among Mozambican cattle breeds7. In contrast with this study, several reports on Cameroonian cattle breeds (FST = 0.061)8, seven European cattle breeds (FST = 0.112)26 and five Asian cattle breeds (FST = 0.117)27 showed higher genetic differentiation. According to Wright28, the differentiation was not negligible if the FST was smaller than 0.05 or even less. Following this interpretation, the cattle populations surveyed were moderately differentiated and structured.
The estimations of pairwise FST values between populations showed that N’Dama population was genetically differentiated from zebu’s populations and more particularly from the Djakore population. The divergence observed between N’Dama and the other cattle populations was supported by genetic distances, phylogenetic relationships and PCA analysis13. The genetic differentiation found between zebu’s populations were very low and suggesting at molecular level, Gobra and Maure might become one breed. In most cases, African zebu’s populations were characterized by low FST values and high levels of genetic diversity relative to a large effective population size and a high migration rate in a usual breeding transhumant system12,29. The relatively low gene flow observed between N’Dama and the other cattle populations could be explained by the farming system and a high reproductive isolation that might imply the mechanisms of speciation30. In contrast, the values of gene flow appraised between zebu’s populations and Djakore showed substantial allelic flow exchanges. This could confirmed between these three populations that the reproduction occurred without none control during the transhumance period13.
The Bayesian approach revealed that Senegalese local cattle populations were grouped into 3 genetic clusters. The cluster 1, where 76.8% of the N’Dama individuals were properly assigned was represented mainly by the N’Dama genotype. At molecular level, N’Dama is a “genetic breed”. As the N’Dama breed was affected by genetic introgression of zebu (Gobra and Maure) with percentages of 7.9 and 12.3%, respectively, therefore it was not a “purebred taurine”. In comparison with the previous studies, MacHugh et al.10 reported that 5.1% of zebu specific-alleles and 12.4% of genetic admixture proportion were found in the N’Dama breed of Senegal. These findings were confirmed by those of Hanotte et al.11. Accordingly, the current genotypic structure observed in most trypanotolerant breeds and more particularly in the N’Dama breed of Senegal was similar to that described in previous studies following the same context12.
Djakore cattle were mainly assigned in the genetic cluster 2. However, 43.1 and 44.1% of Gobra and Maure cattle individuals were assigned in this cluster, respectively. The measures of zebu admixture coefficient (qi) resulted from the studies of Freeman et al.12 supported these results and confirmed the significant presence of zebu alleles in “hybrid” populations from west Africa. According to these findings, Djakore is genetically a stable “hybrid” population and not a “genetic breed”. This confirmed the theory of Broutin et al.31, according to which: Djakore was considered as a “hybrid” population because of its phenotypic characters and geographical distribution. The current genotypic structure of Djakore revealed that zebu introgression was the upshot of the high levels of genetic diversity found in this population.
In the genetic cluster 3, the zebu genotype was predominant with 49 and 43% of Gobra and Maure cattle individuals properly assigned. Compared to the results obtained by MacHugh et al.10, 39.7 and 36.4% of zebu alleles were found, respectively in Maure and Gobra cattle sampled from Senegal. This implies that the increasing of zebu alleles’ proportion followed up significantly the rate of gene flow and the zebu gradient introgression. Since zebu cattle have shared the same genotype, it was inferred that these two populations might become one breed at molecular level. These results contradicted the phenotypic classification that considered them as two distinct breeds32. To sum up, at molecular level, Senegalese local cattle populations are consisted of 3 genotypes: N’Dama genotype, Djakore genotype and zebu genotype.
CONCLUSION
This study was the first to provide, at molecular level, a complete understanding of the diversity and genetic structure of local cattle breeds in Senegal. So, it was imperative to report that Senegalese local cattle populations were characterized by high levels of diversity and a moderate genetic structure. Based on these results, it has been recommended to apply the separate conservation of zebu’s populations (Gobra and Maure) as the highest priority to avoid loss of genetic diversity. N’Dama and Djakore populations should be conserved together to reduce costs and preserve adaptive traits that are valuable for improving cattle breeds’ productivity and conservation.
SIGNIFICANCE STATEMENT
This study has discovered high levels of diversity and a moderate genetic structure in Senegalese cattle populations underlining a wide range of adaptive traits that can be beneficial for breed improvement and conservation strategies. This study has helped the researcher to uncover the critical areas of animal breeding and conservation genetics that many researchers were not able to explore. Thus a new theory on cattle breeds’ diversity and genetic structure may have been reached.
ACKNOWLEDGMENTS
The authors wish to acknowledge the “Services Régionaux d’Elevage de Koalack, Saint-Louis et Kolda” and the Direction of Zootechnical Research Center in Kolda for their kind support during the animal sampling and data collection. We appreciated the help of the “Laboratoire de Biologie des Populations Animales Sahélo-Soudaniennes (BIOPASS) du CBGP/IRD” where DNA extraction was carried out. We are thankful also to the “Centre International de Recherche-Developpement sur l’Elevage en zone Subhumide (CIRDES)” where genotyping of microsatellite loci was performed. Thanks are due also to farmers and shepherds for their collaboration.
REFERENCES
- Belew, A.K., K. Tesfaye, G. Belay and G. Assefa, 2016. The state of conservation of animal genetic resources in developing countries: A review. Int. J. Pharma Med. Biol. Sci., 5: 58-66.
Direct Link - Kristensen, T.N., A.A. Hoffmann, C. Pertoldi and A.V. Stronen, 2015. What can livestock breeders learn from conservation genetics and vice versa? Front. Genet., Vol. 6.
Direct Link - Groeneveld, L.F., J.A. Lenstra, H. Eding, M.A. Toro and B. Scherf et al., 2010. Genetic diversity in farm animals: A review. Anim. Genet., 41: 6-31.
CrossRefDirect Link - Maozami-Goudarzi, K., D.M. Belemsega, G. Ceriotti, D. Laloe and F. Fagbohoum et al., 2001. Caracterisation de la race bovine Somba a l'aide de marqueursmoleculaires. Rev. Elev. Med. Vet. Pays Trop., 54: 129-138.
Direct Link - Bessa, I., I. Pinheiro, M. Matola, K. Dzama, A. Rocha and P. Alexandrino, 2009. Genetic diversity and relationships among indigenous Mozambican cattle breeds. S. Afr. J. Anim. Sci., 39: 61-72.
Direct Link - Ema, P.N., Y. Manjeli, F. Meutchieyie, C. Keambou and B. Wanjala et al., 2014. Genetic diversity of four Cameroonian indigenous cattle using microsatellite markers. J. Livestock Sci., 5: 9-17.
Direct Link - MacHugh, D.E., M.D. Shriver, R.T. Loftus, P. Cunningham and D.G. Bradley, 1997. Microsatellite DNA variation and the evolution, domestication and phylogeography of taurine and zebu cattle (Bos Taurus and Bos indicus). Genetics, 146: 1071-1086.
Direct Link - Hanotte, O., C.L. Tawah, D.G. Bradley, M. Okomo, Y. Verjee, J. Ochieng and J.E.O. Rege, 2000. Geographic distribution and frequency of a taurine Bos Taurus and an indicine Bos indicus Y specific allele amongst sub‐Saharan African cattle breeds. Mol. Ecol., 9: 387-396.
CrossRefDirect Link - Freeman, A.R., C.M. Meghen, D.E. Machugh, R.T. Loftus and M.D. Achukwi et al., 2004. Admixture and diversity in West African cattle populations. Mol. Ecol., 13: 3477-3487.
CrossRefDirect Link - Ndiaye, N.P., A. Sow, G.K. Dayo, S. Ndiaye, G.J. Sawadogo and M. Sembene, 2015. Genetic diversity and phylogenetic relationships in local cattle breeds of Senegal based on autosomal microsatellite markers. Vet. World, 8: 994-1005.
Direct Link - Weir, B.S. and C.C. Cockerham, 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358-1370.
CrossRefDirect Link - Piry, S., A. Alapetite, J.M. Cornuet, D. Paetkau, L. Baudouin and A. Estoup, 2004. GENECLASS2: A software for genetic assignment and first-generation migrant detection. J. Heredity, 95: 536-539.
CrossRefDirect Link - Cornuet, J.M., S. Piry, G. Luikart, A. Estoup and M. Solignac, 1999. New methods employing multilocus genotypes to select or exclude populations as origins of individuals. Genetics, 153: 1989-2000.
PubMedDirect Link - Paetkau, D., W. Calvert, I. Stirling and C. Strobeck, 1995. Microsatellite analysis of population structure in Canadian polar bears. Mol. Ecol., 4: 347-354.
CrossRefDirect Link - Pritchard, J.K., M. Stephens and P. Donnelly, 2000. Inference of population structure using multilocus genotype data. Genetics, 155: 945-959.
PubMedDirect Link - Earl, D.A. and B.M. vonHoldt, 2012. Structure harvester: A website and program for visualizing structure output and implementing the Evanno method. Conservation Genet. Resour., 4: 359-361.
CrossRefDirect Link - Evanno, G., S. Regnaut and J. Goudet, 2005. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol., 14: 2611-2620.
CrossRefPubMedDirect Link - Chaudhari, M.V., S.N.S. Parmar, C.G. Joshi, C.D. Bhong, S. Fatima, M.S. Thakur and S.S. Thakur, 2009. Molecular characterization of Kenkatha and Gaolao (Bos indicus) cattle breeds using microsatellite markers. Anim. Biodivers. Conser., 32: 71-76.
Direct Link - MacHugh, D.E., R.T. Loftus, P. Cunningham and D.G. Bardley, 1998. Genetic structure of seven European cattle breeds assessed using 20 microsatellite markers. Anim. Genet., 29: 333-340.
CrossRefDirect Link - Shi, Z., J.H. Lee, Y.S. Lee, D.Y. Oh and J.S. Yeo, 2010. Analysis of genetic diversity and distances in Asian cattle breeds using microsatellite markers. J. Korean Data Infor. Sci. Soc., 21: 795-802.
Direct Link - Dayo, G.K., S. Thevenon, D. Berthier, K. Moazami-Goudarzi and C. Denis et al., 2009. Detection of selection signatures within candidate regions underlying trypanotolerance in outbred cattle populations. Mol. Ecol., 18: 1801-1813.
CrossRefDirect Link - Penda, N.N., S. Adama, N. Saliou, S. Mbacke and S.G. Jerome, 2014. Phenotypical characterization of Senegalese local cattle breeds using multivariate analysis. J. Anim. Vet. Adv., 13: 1150-1159.
Direct Link