
Edokpolor Osazee Ohanmu
Department of Plant Biology and Biotechnology, Faculty of Science, Edo University, Iyamho, Edo State, Nigeria
LiveDNA: 234.24989
Beckley Ikhajiagbe
Environmental Biotechnology and Sustainability Research Group, Department of Plant Biology and Biotechnology, University of Benin,
Benin, Nigeria
Asian Journal of Applied Sciences
Year: 2018 | Volume: 11 | Issue: 3 | Page No.: 125-134







ABSTRACT
Background and Objective: Environmental stress such as heavy metal is a major factor affecting reduction in agricultural productivity. Heavy metals at toxic levels hamper normal plant functioning and act as an impediment of metabolic processes by the production of Reactive Oxygen Species (ROS). Plant has evolved various enzymatic and non-enzymatic mechanism to cope against the ROS produced. This study investigated the biochemical response of S. stenocarpa to cadmium stress. Materials and Methods: S. stenocarpa (Fabaceae) seeds were sown in the control (0 ESV) and the different concentrations (2.5, 5 ESV) of cadmium chloride (CdCl2). The soil samples were collected from 10 different points at a depth of 0-30 cm using a soil auger, pooled together, air-dried and grind to pass through a 2 mm sieve before subjecting to physico-chemical analysis. The five accessions consisted of 3 blocks, 15 treatments per block, two plant per polythene bag arranged in a RBD. Enzymes such as superoxide dismutase (SOD), catalase (CAT), malondialdehyde (MDA), proline as well as non-enzymes such as tocopherol, ascorbate were determined. Results: Increased cadmium concentration resulted in increased translocation of the metal in the plant leaves and decreased chlorophyll a/b content of TSs-accessions. Increase in cadmium concentration results to variability in enzymatic and non-enzymatic response of the studied accessions. Take for instance overall MDA, SOD and proline activities were significantly increased in the Cd-5ESV as compared to the control. However, the accessions varied in their response to cadmium. For instance, the SOD of TSs-95 was reduced with increased cadmium concentration. Similarly, the ascorbate and tocopherol content decreased in TSs-92 and TSs-95, respectively with increased Cd concentration. Individual plants showed different antioxidant responses either between the plants or among the accessions. The MDA activity significantly increased in leaves with metal increase while SOD activity was heighten in TSs-91 whereas in TSs-95 there were no significant difference in SOD activity between control and Cd-exposed plant. Conclusion: The increase in antioxidant activities of the metal exposed plants compared to their control counterpart suggested possibility of free radicals instigated by metal presence. This further gave trendies to plants’ resistance capacities with SOD been the first line of defense.
PDF Abstract XML References Citation
Received: September 16, 2018;
Accepted: October 12, 2018;
Published: November 17, 2018
How to cite this article
Edokpolor Osazee Ohanmu and Beckley Ikhajiagbe, 2018. Enzymatic and Non-Enzymatic Response of Sphenostylis stenocarpa to Cadmium Stress. Asian Journal of Applied Sciences, 11: 125-134.
DOI: 10.3923/ajaps.2018.125.134
URL: https://scialert.net/abstract/?doi=ajaps.2018.125.134
DOI: 10.3923/ajaps.2018.125.134
URL: https://scialert.net/abstract/?doi=ajaps.2018.125.134
INTRODUCTION
Cadmium is highly toxic to all living organisms and is one of the most toxic heavy metals in plants due to its high solubility in water and phytotoxicity. The detrimental effects of cadmium are manifested in decreasing soil biological activity and plant metabolism1. There are a number of natural and anthropogenic processes which add Cd to soils and waters. Natural sources include volcanic eruptions but the amount of Cd released is usually small in comparison to man-made sources2. The latter include wastes from factories, cement manufacturing, waste incineration, fossil fuel combustion, sewage sludge, phosphate fertilizers and most importance, nonferrous mining, especially Pb and Zn mining, which is the biggest input source of Cd to soils. Cadmium has received great attention in soil science and plant nutrition mainly due to its phytotoxic impact ranging from growth reduction, wilting and chlorosis to cell death3; its relative high mobility in the soil–plant system, which implies Cd dissemination throughout the food chain, even becoming a serious threat to ecosystem and human health4.
To cope with highly toxic metals plant has evolved complex mechanisms that serve to control the uptake, accumulation and detoxification of metals. Agricultural crops differ widely in their tolerance to metals toxicity. Legume crops are less tolerant to cadmium toxicity than cereals and grasses and encounter strong inhibition of biomass production. Identification of plant accessions differing in resistance to heavy metals is a promising approach in plants defense mechanisms against toxic metals. Metals at toxic levels hamper normal plant functioning and act as an impediment of metabolic processes in a variety of ways including disturbance or displacement of building blocks of protein structure5, hindering functional group of important cellular molecules6 and disrupt functionality of essential metals. On the other hand, elevated level of heavy metals is associated with the increased generation of reactive oxygen species (ROS) such as superoxide free radicals, hydroxyl free radicals or non-free radical species (molecular form) such as singlet oxygen and hydrogen peroxide as well as cytotoxic compounds like methylglyoxal (MG), which can cause oxidative stress by disturbing the equilibrium between prooxidant and antioxidant homeostasis cells7.
The ability to fix atmospheric nitrogen makes legumes excellent components within the various farming systems because they provide residual nitrogen and reduce the needs for mineral nitrogen fertilizers by associated non-legumes. Sphenostylis stenocarpa Hochst ex. A. Rich. (Harms) are an underutilized tropical African tuberous legume mainly cultivated in West Africa. The utilization of S. stenocarpa has links with sociocultural values in the cultures of some ethnic groups. Assefa and Kleiner8 remarked that S. stenocarpa has very high nitrogen fixing ability. It also has a remarkably low susceptibility to most field and storage leguminous pests9. The arable tuberous legume is important in most indigenous African food cultures and in peasant agriculture. In Nigeria, it is cultivated in Edo State mostly for its seeds than tubers. The seeds are high in vitamin C, dietary fiber, vitamin B6, potassium and manganese; while being low in saturated fat, sodium and cholesterol. The main objective of this research was to investigate the enzymatic and non-enzymatic response of S. stenocarpa accessions to cadmium stress. However, there is little to no information about the best defense mechanism employed and the physiological changes in plant against cadmium stress of the studied accessions in Nigeria.
MATERIALS AND METHODS
Soil collection and land preparation: The experiment was carried out in the Botanical garden of Plant Biology and Biotechnology, University of Benin, Benin city. The soils were collected from ten different points at the botanic garden, where the experiment was carried out. The samples were randomly collected from the site at 0-30 cm depths using a soil auger. The soil samples were made into a composite sample, air-dried and grinded to pass through a 2 mm sieve before subjecting to physico-chemical analysis. Polythene bags (50×50 cm) were filled with 15 kg of air dried soil. From the physical analysis, the soil type was sandy loam, pH 7.3. The soils were polluted with cadmium in the form of cadmium chloride (CdCl2) using 0, 2.5 and 5 ESV (Ecological screening value)10. The treatment designation for metal concentration had been reported (Table 1). Seeds of S. stenocarpa (Africa yam bean accessions: TSs-91, TSs-92, TSs-93, TSs-94 and TSs-95) were procured from International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria.
| Table 1: | Treatment designations for metal concentrations |
 | |
| ESV: Ecological screening value | |
Experimental design: There were 3 blocks comprising of 15 treatments per block, 2 plant per polythene bag amounting to a total of 90 plants arranged in a Randomized Block Design (RBD). The experiment setup spanned from late October, 2016 to March, 2017. The plant antioxidant defense mechanisms and chlorophyll contents were determined at young plant stage (42 days after sowing). Chlorophyll determination was according to the method described by Arnon11.
In other to assess plant antioxidant response, ascorbic acid12, catalase13, proline14, superoxide dismutase15 and tocopherol16 levels were quantified in plant leaves. Lipid peroxidation was measured by estimating the malondialdehyde levels17.
Statistical analysis: Having assumed the experimental setup to be homogenous, a one-way analysis of variance (IBM SPSS 20) with significance level p<0.05 was adopted for data analyses. Selected parametric means were also correlated where necessary (Pearson’s correlation used).
RESULTS AND DISCUSSION
Chlorophyll content: The chlorophyll contents of African yam bean accessions (TSs-91, TSs-92, TSs-93, TSs-94 and TSs-95) to cadmium toxicity were presented in Fig. 1. Cadmium pollution resulted in an insignificant difference in the chlorophyll-a levels of TSs-91 between the control and the Cd-5ESV. The growth inhibition produced by cadmium is partially due to its effects on the photosynthesis rate18. In this study, it was reported that cadmium stress decreased the chlorophyll-a content in TSs-92 with increased metal concentration. Ayub et al.19 recorded chlorophyll contents are more sensitive to cadmium toxicity than nickel. The effects of cadmium pollution on chlorophyll-b content of Africa yam bean accessions was also reported. Cadmium pollution resulted to an increased chlorophyll-b content of TSs-91 from 0.022-0.136 mg g–1 FW in the control and Cd-5ESV, respectively. In general, HM reduce photosynthetic rates, affecting both light and dark reactions. Moreover, the action of metal ions on plants is modulated by other metal ions and the nature of this interaction seems to be complex20.
Modal foliar colour: The effects of cadmium pollution on the modal foliar colour of Africa yam bean accessions (TSs-91, TSs-92, TSs-93, TSs-94 and Tss-95) (Table 2). Cadmium pollution resulted to change in the leaves pigment colour from yellow green to chartreuse and olive drab in TSs-91, TSs-92, TSs-93, TSs-94 and TSs-95 sown in Cd-5ESV. The incidence of necrosis and chlorosis exasperated by increased metal concentration in the plant accessions.
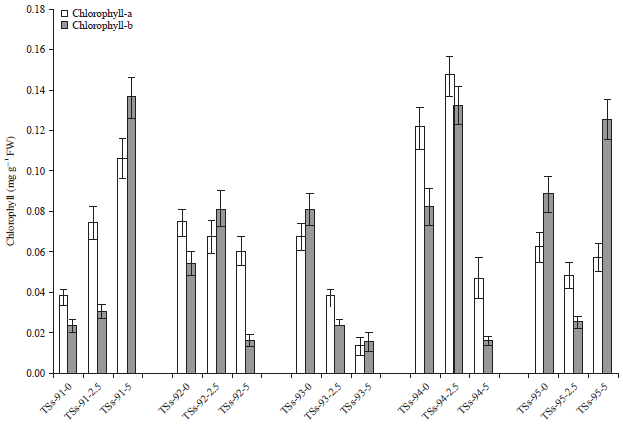 | |
| Fig. 1: | Effect of cadmium stress on chlorophyll contents |
| Table 2: | Effects of cadmium pollution on modal foliar colour of African Yam Bean |
 | |
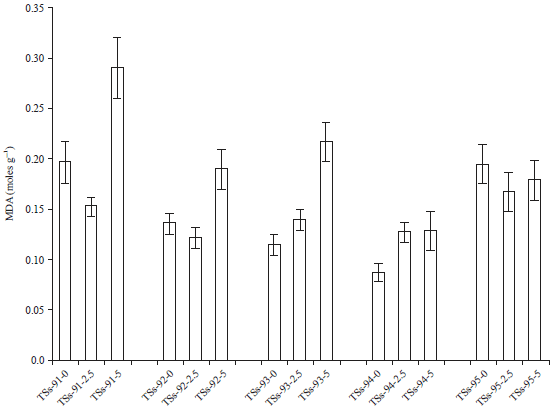 | |
| Fig. 2: | Effect of cadmium pollution on malondialdehyde activity |
Leaf chlorosis is one of the most commonly observed consequences of Cd toxicity. The chlorosis were more pronounced in the older leaves than new leaves irrespective of accession type. Maybe the plants employed this as a survival mechanism to cope with the HM stress.
Malondialdehyde (MDA) activity: The increased contents of lipid peroxides are indication of more production of toxic singlet oxygen species than normal. Figure 2 showed the effect of cadmium pollution on the malondialdehyde activity of 5 accessions of Africa yam. Cadmium pollution resulted in an increase in the MDA activities with increased metal concentration from 0.13-0.22 moles g–1 between the control and Cd-5ESV in TSs-93. When plants are grown in stress environments, free-radicals generates in excess and accumulate in the cells.
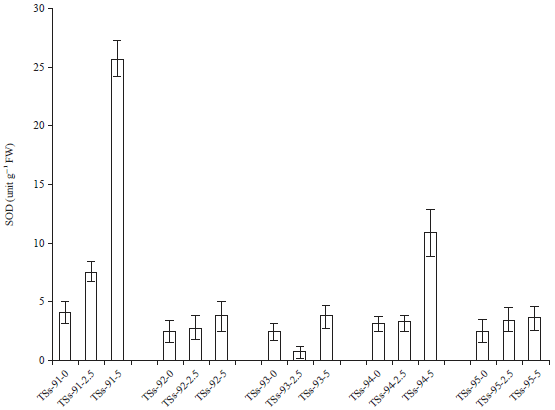 | |
| Fig. 3: | Effect of cadmium pollution on superoxide dismutase activity |
This leads to lipid peroxidation of biomembranes and its end product is MDA. Therefore, the MDA-concentration is an indicator of physiological stress and the aging process21. However, each accessions responded differently to the cadmium exposure. For example, the MDA activity of TSs-95 was reduced with increase in metal concentration as compared to the other accessions. However, there was no significant difference at p = 0.34.
Superoxide dismutase (SOD) activity: The data in Fig. 3 presented the effect of cadmium pollution on the superoxide dismutase activity of Africa yam bean accessions. Cadmium pollution resulted in an increase in the superoxide dismutase activities with increased metal concentration. However, some accessions had significantly (p = 0.02) increased SOD activities take for instance, TSs-91 and TSs-94 from 4-26 unit g–1 FW and 3.6-11.5 unit g–1 FW between the control and Cd-5ESV respectively. Response of SOD enzymes activate the essential component of the plant antioxidative defense system as they dismutase’s two O2– to water and oxygen. The results verified an enhancement in Africa yam bean susceptibility in regards to SOD activity was heighten with increase metal concentration in TSs-91 whereas in TSs-95 there were no significant difference in SOD activity between the control and the metal exposed plant. The increase in SOD activity has been reported previously for certain plant species exposed to toxic Cd concentrations22-25.
Catalase (CAT) activity: The effect of cadmium pollution on the catalase activity of Africa yam bean accessions is shown in Fig. 4. Cadmium pollution resulted in an increased catalase activity of 0.74 unit g–1 FW in Cd-2.5 ESV compared to a reduction of 1.1 unit g–1 FW in Cd-5 ESV of TSs-92 with increase in the metal concentration. In this study, it was observed that accessions in the cadmium polluted soil had reduced CAT activities as compared to the control. However, there was no significant difference p = 0.07. According to Ahsan et al.26 under the influence of stress caused by metals, quantitative and qualitative changes of proteins occur in plants. The presence of HMs stress may be associated with impaired basic steps in the metabolism of carbohydrates and amino acids27.
Ascorbate content: The effects of cadmium pollution on the ascorbate activity of Africa yam bean accessions has been reported (Fig. 5). Although, there was no significant difference p = 0.12, cadmium exposure resulted to an increase in the ascorbate content of TSs-accessions except in TSs-91 and TSs-94 sown in the Cd-2.5 ESV. The ascorbate content of TSs-91 and TSs-94 in Cd-2.5 ESV were significantly reduced to 0.0001 μM. The increased ascorbate content reported irrespective of metal concentration is due to the productive role played by the non-enzyme.
 | |
| Fig. 4: | Effect of cadmium pollution on catalase activity |
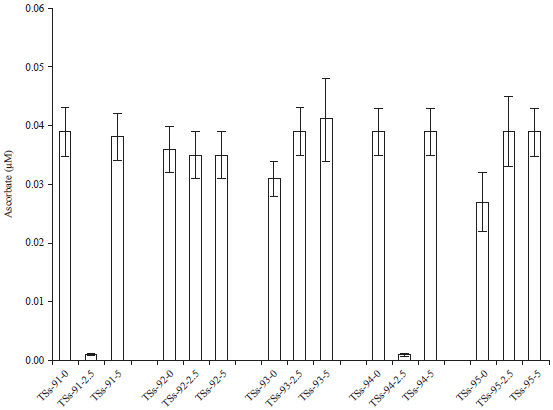 | |
| Fig. 5: | Effect of cadmium pollution on ascorbate content |
Ascorbate appeared to play an essential protective role in the scavenging process when coordinated with SOD activity28. Apparently, SOD leads to the over-production of H2O2 to eliminate the toxicity of O2–.
Proline content: The effect of cadmium pollution on the proline content of Africa yam bean accessions is presented in Fig. 6. The metabolite proline serves multiple functions in plant stress adaptions.
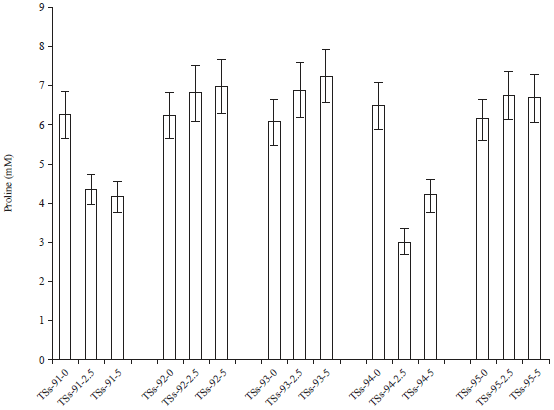 | |
| Fig. 6: | Effect of cadmium pollution on proline content |
On exposure to cadmium, the proline content of TSs-91 decreased as compare to an increase in TSs-92 with increased metal concentration. The decrease of proline content in the TSs-91 and TSs-94 Cd-stressed soils as compared to the control may be as a results of enzymatic and non-enzymatic scavengers working collectively to mitigate the free radicals produced. Proline works as protein-compatible hydrotrope, osmo-protectant, ROS scavenger and regulator of cellular redox status which maybe the reason for the increased proline content observed. It also regulates the redox signal governing the metabolite pool and expression of several genes that affect plant growth and development29. Hence there was a significant difference (p = 0.02) in cadmium exposure to proline content of Africa yam bean accessions.
Tocopherol content: The effect of cadmium pollution on the tocopherol content of Africa yam bean accessions is shown in Fig. 7. Abiotic stress triggers a variety of plant responses like changes in gene expression and cell metabolism. Cadmium pollution resulted to a significant difference in the tocopherol content of AYB accessions at p = 0.007. In this study, there was increased tocopherol content with increased cadmium stress irrespective accession type or concentration except in TSs-91, TSs-93 and TSs-94 in the Cd-5ESV. Tocopherol deactivates photosynthesis-derived ROS and prevented the propagation of lipid peroxidation.
Factorial analysis: The principal component of analysis was conducted to determine the prominent defense mechanism employed by the various accessions of African yam bean (Fig. 8). Individual accessions showed different antioxidant responses. From the principal component analysis (PCA), SOD, MDA and ASA were significant in combating the ROS produced by cadmium pollution. Significant increases in antioxidant activities of the metal exposed plants compared to their control counterpart suggested possibility of free radicals instigated by HM presence. This gave rise to plants’ resistance capacities in the metal polluted soil.
This study gives an extensive knowledge of the valuable foliar shades and defense mechanism employed by S. stenocarpa to combat cadmium stress in soil. The achievement of high SOD and ascorbate activities can be obtained by exploring their response individually or holistic. In the future, molecular studies and genetic engineering can be explored to identify and harness strand that produce high amount of SOD and ascorbate for increase plant performance against cadmium. Obviously, this will increase plant resistance to cadmium stress, reduce MDA levels and increase food productivity.
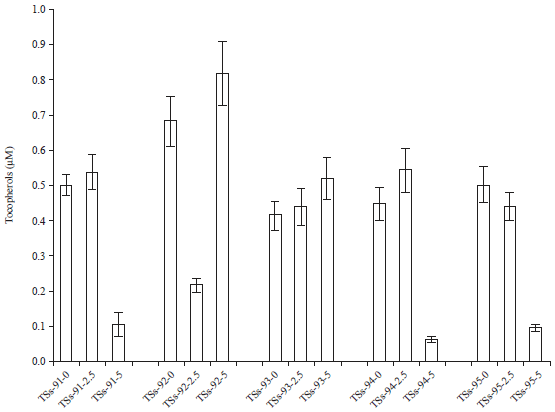 | |
| Fig. 7: | Effect of cadmium stress on tocopherol content |
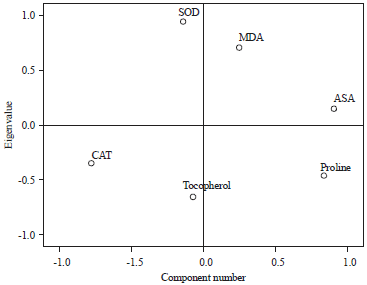 | |
| Fig. 8: | Principal component activity of the enzymatic and non-enzymatic activities |
CONCLUSION
The presence of cadmium in the cell led to the formation of Reactive Oxygen Species (ROS) with the potential to further severe oxidative damage to different cell organelles and biomolecules. These oxidative stress indirectly interferes with the activity of the antioxidant system. A constitutively high antioxidant capacity or increase in antioxidant level could prevent oxidative damage and improve tolerance to the oxidative stress established.
SIGNIFICANCE STATEMENT
This study discovers the first line of enzymatic and non-enzymatic defense mechanism employed by the accessions of S. stenocarpa against cadmium exposure was superoxide dismutase and ascorbate, respectively. This study will help researchers and farmer to reveal the critical areas of the plant inherent defense strategies against ROS in the studied accessions. In Nigeria, application of phosphate fertilizer is increasing the cadmium content in soil. Heavy metals are notoriously hard to remediate and the option is to find resistance genotypes/accessions that will perform well in such soil. Exploration of underutilized legumes in Africa have encouraged a growing research interest to combat food insecurity especially in Nigeria.
ACKNOWLEDGMENT
Enormous thanks are due to those who have contributed in this research, especially to the Environmental Biotechnology and Sustainability Research Group, University of Benin, Benin City who provided a working lab.
REFERENCES
- Di Toppi, L.S. and R. Gabbrielli, 1999. Response to cadmium in higher plants. Environ. Exp. Bot., 41: 105-130.
CrossRefDirect Link - Ohanmu, E.O., B. Ikhajiagbe and B.O. Edegbai, 2018. Nitrogen distribution pattern of african yam bean (Sphenostylis stenocarpa) exposed to cadmium stress. J. Applied Sci. Environ. Manage., 22: 1053-1057.
Direct Link - Gallego, S.M., L.B. Pena, R.A. Barcia, C.E. Azpilicueta and M.F. Iannone et al., 2012. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot., 83: 33-46.
CrossRefDirect Link - Burger, J., 2008. Assessment and management of risk to wildlife from cadmium. Sci. Total. Environ., 389: 37-45.
CrossRefDirect Link - Hall, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 53: 1-11.
CrossRefDirect Link - Hossain, M.A., P. Piyatida, J.A.T. da Silva and M. Fujita, 2012. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot.
CrossRefDirect Link - Sytar, O., A. Kumar, D. Latowski, P. Kuczynska, K. Strzalka and M.N.V. Prasad, 2013. Heavy metal-induced oxidative damage, defense reactions and detoxification mechanisms in plants. Acta Physiol. Planta., 35: 985-999.
CrossRefDirect Link - Assefa, F. and D. Kleiner, 1997. Nodulation of African yam bean (Sphenostylis stenocarpa) by Bradyrhizobium sp. isolated from Erythrina brucei. Biol. Fertil. Soils, 25: 209-210.
CrossRefDirect Link - Omitogun, O.G., L.E.N. Jackai and G. Thottappilly, 1999. Isolation of insecticidal lectin-enriched extracts from African yam bean (Sphenostylis stenocarpa) and other legume species. Entomol. Exp. Applicata, 90: 301-311.
CrossRefDirect Link - Arnon, D.I., 1959. Conversion of light into chemical energy in photosynthesis. Nature, 184: 10-21.
CrossRefPubMedDirect Link - Huang, D., B. Ou and R.L. Prior, 2005. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem., 53: 1841-1856.
CrossRefPubMedDirect Link - Cohen, N.D., M.F. Utter, N.G. Wrigley and A.N. Barrett, 1979. Quaternary structure of yeast pyruvate carboxylase: Biochemical and electron microscope studies. Biochemistry, 18: 2197-2203.
CrossRefDirect Link - Marin, A.P., M.I. Aguilar, V.F. Meseguer, J.F. Ortuno, J. Saez and M. Llorens, 2009. Biosorption of chromium (III) by orange (Citrus cinensis) waste: Batch and continuous studies. Chem. Eng. J., 155: 199-206.
CrossRefDirect Link - Hossain, M.M., R. Jahangir, S.R. Hasan, R. Akter, T. Ahmed, M.I. Islam and A. Faruque, 2009. Antioxidant, analgesic and cytotoxic activity of Michelia champaca Linn. leaf. Stamford J. Pharm. Sci., 2: 1-7.
Direct Link - Yu, H.H., T. Awasaki, M.D. Schroeder, F. Long and J.S. Yang et al., 2013. Clonal development and organization of the adult Drosophila central brain. Curr. Biol., 23: 633-643.
CrossRefDirect Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Metwally, A., I. Finkemeier, M. Georgi and K.J. Dietz, 2003. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol., 132: 272-281.
CrossRefDirect Link - Chen, Y.X., Y.F. He, Y.M. Luo, Y.L. Yu, Q. Lin and M.H. Wong, 2003. Physiological mechanism of plant roots exposed to cadmium. Chemosphere, 50: 789-793.
CrossRefDirect Link - Shah, K., R.G. Kumar, S. Verma and R.S. Dubey, 2001. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings. Plant Sci., 161: 1135-1144.
CrossRefDirect Link - Vitoria, A.P., P.J. Lea and R.A. Azevedo, 2001. Antioxidant enzymes responses to cadmium in radish tissues. Phytochemistry, 57: 701-710.
CrossRefDirect Link - Iannelli, M.A., F. Pietrini, L. Fiore, L. Petrilli and A. Massacci, 2002. Antioxidant response to cadmium in Phragmites australis plants. Plant Physiol. Biochem., 40: 977-982.
CrossRefDirect Link - Ouariti, O., N. Boussama, M. Zarrouk, A. Cherif and M.H. Ghorbal, 1997. Cadmium- and copper-induced changes in tomato membrane lipids. Phytochemistry, 45: 1343-1350.
CrossRefPubMedDirect Link - Ahsan, N., S.H. Lee, D.G. Lee, K.W. Lee, D.H. Kim, J.D. Bahka and B.H. Lee, 2007. Physiological and protein profiles alternation of germinating rice seedlings exposed to acute cadmium toxicity. Comptes Rendus Biol., 330: 735-746.
CrossRefPubMedDirect Link - Rahoui, S., A. Chaoui and E. El Ferjani, 2010. Membrane damage and solute leakage from germinating pea seed under cadmium stress. J. Hazard. Mater., 178: 1128-1131.
CrossRefDirect Link - Massacci, A., M.A. Iannelli, F. Pietrini and F. Loreto, 1995. The effect of growth at low temperature on photosynthetic characteristics and mechanisms of photoprotection of maize leaves. J. Exp. Bot., 46: 119-127.
CrossRefDirect Link - Hayat, S., B. Ali, S.A. Hassan and A. Ahmed, 2007. Brassinosteroid enhanced the level of antioxidants under cadmium stress in Brassica juncea. Environ. Exp. Bot., 60: 33-41.
CrossRefDirect Link