
E. Nagyzbekkyzy
RSE Republican Collection of Microorganisms, SC MES RK, Astana, Kazakhstan
G. Abitayeva
RSE Republican Collection of Microorganisms, SC MES RK, Astana, Kazakhstan
S. Anuarbekova
RSE Republican Collection of Microorganisms, SC MES RK, Astana, Kazakhstan
D. Shaikhina
RSE Republican Collection of Microorganisms, SC MES RK, Astana, Kazakhstan
K. Li
RSE Republican Collection of Microorganisms, SC MES RK, Astana, Kazakhstan
S. Shaikhin
RSE Republican Collection of Microorganisms, SC MES RK, Astana, Kazakhstan
LiveDNA: 7310.12583
K. Almagambetov
RSE Republican Collection of Microorganisms, SC MES RK, Astana, Kazakhstan
A. Abzhalelov
RSE Republican Collection of Microorganisms, SC MES RK, Astana, Kazakhstan
S. Saduakhassova
Nazarbayev University, Astana, Kazakhstan
A. Kushugulova
Nazarbayev University, Astana, Kazakhstan
F. Marotta
ReGenera Research Group for Aging Intervention and Milano Medical, Milano, Italy
Asian Journal of Applied Sciences
Year: 2016 | Volume: 9 | Issue: 4 | Page No.: 143-158







ABSTRACT
Background and Objective: Much attention is paid to obtaining novel promising Lactobacillus strains with probiotic and technological properties in the food industry. Probiotics also have higher possibility to become pathogens due to acquiring antibiotic resistance genes. The purpose of the present study was to select potential probiotic Lactobacillus strains among isolates from Kazakh traditional fermented dairy foods that will be promising candidates as starter cultures in the production of probiotic fermented foods. Materials and Methods: Samples were prepared from homemade and industrial fermented dairy foods purchased in Kazakh markets. Identification of lactic acid bacteria was carried out by 16S rRNA gene sequence analysis using the BLAST algorithm. Tolerance of lactobacilli to simulating gastrointestinal tract conditions was evaluated in a consecutive exposure to hydrochloric acid (pH 3.0 for 1 h) and bile acids (0.3% for 1 h) and statistical significance was determined by the overlap rule for SE bars. Antimicrobial activity was measured by deferred antagonism procedure. Antibiotic resistance was tested by agar disk diffusion assay. Results: Sixteen Lactobacillus strains exposed to simulating gastrointestinal tract conditions showed almost identical responses, only two strains showed about 2 times less tolerance (p<0.05). Six novel isolates had strong antimicrobial activity against 2 pathogens, 3 novel isolates had strong antimicrobial activity against three pathogens and 2 novel isolates had a broad antimicrobial spectrum including antifungal activity against C. albicans. Resistance/intermediate resistance to tetracycline, β-lactams, streptomycin and gentamicin of 8 novel isolates were found. Conclusion: Novel strains L. casei BI005 and 1LB were selected as potential probiotics because of beneficial properties viz., tolerance to simulating gastrointestinal tract conditions, high antimicrobial activity and lack of acquired genes of antibiotic resistance. Detection of resistance to tetracycline for 4 isolates (3 novel isolates inclusive) from three different phylogenetic groups among 16 leads to the opinion that lactobacilli with tetracycline resistance are common in dairy foods consumed in Kazakhstan.
PDF Abstract XML References Citation
Received: May 23, 2016;
Accepted: August 15, 2016;
Published: September 15, 2016
How to cite this article
E. Nagyzbekkyzy, G. Abitayeva, S. Anuarbekova, D. Shaikhina, K. Li, S. Shaikhin, K. Almagambetov, A. Abzhalelov, S. Saduakhassova, A. Kushugulova and F. Marotta, 2016. Investigation of Acid and Bile Tolerance, Antimicrobial Activity and Antibiotic Resistance of Lactobacillus Strains Isolated from Kazakh Dairy Foods. Asian Journal of Applied Sciences, 9: 143-158.
DOI: 10.3923/ajaps.2016.143.158
URL: https://scialert.net/abstract/?doi=ajaps.2016.143.158
DOI: 10.3923/ajaps.2016.143.158
URL: https://scialert.net/abstract/?doi=ajaps.2016.143.158
INTRODUCTION
The purpose of the present study was to select potential probiotic Lactobacillus strains among isolates from Kazakh traditional fermented dairy foods that will be promising candidates as starter cultures in the production of probiotic fermented foods.
Probiotics are non-pathogenic microorganisms that, when ingested in adequate amounts, exert a positive influence on their host’s health1. A variety of microorganisms, mainly lactic acid bacteria (LAB) have been evaluated for their probiotic potential and are applied in various types of food products or in therapeutic preparations2.
The LAB are widely distributed in nature and occur usually as indigenous microflora in raw milk, yoghurt, etc. They are Gram positive bacteria that play an important role in many food fermentation processes3. The genus Lactobacillus is the largest group among the LAB and likely most widely used as a probiotic in a variety of foods, mainly meat and fermented dairy products. Phenotypic traits of lactobacilli viz., specie’s obligate/facultative, homo/hetero-fermentation abilities play a crucial role in souring raw milk and in the manufacturing of fermented dairy products viz., fermented milk, cheese, yoghurt and probiotics3.
Characterization of novel promising probiotic strains is a comprehensive process. Functionality and safety of testing isolates are important. The bacteria should be able to survive food processing, storage and digestion conditions. Acid and bile tolerance is essential for its delivery to the small intestine, where it can exert its positive effect4,5. Zago et al.4 screened for probiotic potential a number of L. plantarum strains isolated from Italian and Argentinean cheeses and found a considerable heterogeneity in their properties. In general, the selected strains showed good adaptation to simulated gastric juice and a moderate to low bile tolerance. Among the selected strains, only three L. plantarum strains showed the best probiotic potential and would be promising candidates for inclusion as starter cultures for the manufacture of probiotic fermented foods4. In study of Reale et al.5 13 novel strains among 184 isolates of L. paracasei, L. casei and L. rhamnosus were isolated from wine, human faeces and sourdough niches. They showed better tolerance to high acidity, bile salts and NaCl. The study highlighted a noticeable heterogeneity in the ability of the strains to withstand harsh growth conditions suggesting that different stress factors should be applied to identify the most promising candidates for technological uses5. Observation made by several investigators shows that bile tolerance is a strain-specific trait and tolerances of species cannot be generalized6. But, inspite of the extensive use of Lactobacillus strains in food applications and as commercial probiotics, there is little information on the diversity in the stress tolerance within these species.
Fermented dairy products are the most perishable products in the market. Consumption of Kazakh traditional fermented dairy foods plays a significant role in the supply of important nutrients required for good health. These products form an essential part in the Kazakh diet and hence their contamination that leads to health hazard is of concern. Among functional and probiotic properties antimicrobial activity apparently supports a healthy digestive tract of host organism as well as extends and improves the hygienic safety of fermented dairy products via producing antibacterial and antifungal substances such as acetic, lactic and succinic acids, ethanol, hydrogen peroxide, bacteriocins, cyclic dipeptides, pyroglutamic acid, etc7,8. Rushdy and Gomaa7 isolated L. brevis strain with best antimicrobial activity, high tolerance to low pH and bile salts and antibiotic susceptibility. The results showed the potential of L. brevis strain as a probiotic culture and hence it can be utilized in manufacturing of pharmaceuticals and dietary supplements. In study of Belicova et al.8 among 125 slovak bryndza cheese isolates only eleven L. plantarum isolates with the highest antimicrobial activity against 8 bacterial pathogens were further characterized in vitro for some probiotic and safety properties. Only 3 isolates showed the ability to grow over 50% in the presence of 0.3% bile. Out of 26 Lactobacillus strains with strong antimicrobial activity, 20 were identified as L. plantarum and 6 as L. fermentum.
The misuse of antibiotics in human medicine is believed to be the principal cause of the antibiotic resistance problem. Another aspect is the selection of resistant bacteria in the food chain due to the heavy utilization of antimicrobial agents in animal husbandry9. Considerable attention has been focused on bacterial species that actually cause disease however, harmless and beneficial bacteria continuously exchange genes, leading to a "Reservoir" of resistance genes and providing multiple complex pathways by which resistance genes propagated in animals can directly or more likely indirectly make their way into human pathogen via food10. Consuming products containing starter cultures or probiotic bacteria with transmissible antibiotic resistance could be detrimental instead of beneficial to gut and public health. Traditionally, antibiotic resistance genes were not part of the standard screening assays for starter cultures and probiotic bacteria used in foods or as food supplements11. Thus, fewer physiological and molecular data are available on the antibiotic resistances of lactobacilli present in fermented foods9.
Currently, it is generally accepted that the possibility of transfer is related to the genetic basis of the resistance mechanism, i.e., whether the resistance is intrinsic, acquired as a result of a chromosomal mutation(s) or acquired by horizontal gene transfer12. The vancomycin-resistant phenotype of some lactobacilli is perhaps the best characterized intrinsic resistance in LAB13. Vancomycin binds to the D-alanine/D-alanine terminus of the pentapeptide, preventing polymerization of peptidoglycan precursors. In several species of LAB the terminal D-alanine residue is replaced by D-lactate or D-serine in the muramyl pentapeptide, preventing vancomycin binding and therefore becoming resistant to the antibiotic13. Florez et al.14 identified a single mutation in the 23S rRNA gene reducing the affinity of erythromycin to the ribosome. This mutation conferred macrolide resistance in a strain of L. rhamnosus. In this study, the transfer risk is considered to be very low for intrinsic resistance or acquired resistance due to chromosomal mutation(s).
Antibiotic resistance genes, particularly those which are carried within mobile genetic elements, viz., plasmids and transposons the most likely to be horizontally transmitted and this deserve particular attention15. In study of Nawaz et al.16 the tetracycline resistance tet genes were identified in 12 strains of lactobacilli from traditional China foods. The researchers identified tet(M) and tet(S) genes from L. brevis NWL59 and tet(M) from L. plantarum NWL22. These genes were successfully transferred to Enterococcus faecalis 181 by filter mating. In this respect, the transfer risk is considered to be very high for acquired resistance due to horizontally transferred antibiotic resistance genes carried within plasmids. Although, some effort has been made in this direction, study has only been carried out for some antibiotics and particular Lactobacillus species. These include the most commonly used probiotic species, such as L. casei, L. acidophilus, L. reuteri or L. rhamnosus, among others or the yogurt starter bacteria L. delbrueckii9.
Checking for traits of transmissible antibiotic resistance in novel probiotic bacteria intended for use as feed and food additives is essential, quite apart from the need for their complete physiological and technological characterization13. However, there are difficulties in differentiation between the intrinsic and acquired/transmissible resistances to antibiotics of probiotic bacteria due to the multiplicity of methods available and the complexity of lactobacilli taxonomy. There is a lack of agreement about the resistance/susceptibility breakpoints for the majority of antibiotics in LAB and bifidobacteria. Further, some assays seem not to be reliable for particular antibiotics13. For this reason there are large gaps in MIC breakpoints for LAB standards of the National Committee for Clinical Laboratory Standards9. This problem is important, since it may affect decision on whether resistance can be considered as intrinsic.
Currently, researcher’s attention is focused on obtaining from indigenous dairy products probiotic strains with satisfactory technological properties suitable in the food and pharmaceutical industries17. Such indigenous dairy products are sources of entirely new strains, which have often unique properties. The literature review did not lead to the works in which the potential of probiotics have been studied from the Kazakh traditional homemade products, such as mare’s milk, Airan, etc., within the framework of experimental design presented in this study. Traditionally analyses of antibiotic resistance profiles was not part of the standard screening assays for starter cultures and probiotic bacteria used in foods or as food supplements in Kazakhstan. Therefore the results obtained in the present work and especially those relating to antibiotic resistance or susceptibility are new and original. The purpose of the present study was to select potential probiotic Lactobacillus strains among isolates from Kazakh traditional fermented dairy foods that will be promising candidates as starter cultures in the production of probiotic fermented foods. To achieve the purpose of the work several problems had to be solved, viz., isolation of lactic acid bacteria and evaluation of their tolerance to bile in order to choose more tolerant isolates, then identification of the lactic acid bacteria isolates as Lactobacillus spp., by nucleotide sequence analysis of 16S rRNA gene and evaluation of their tolerance to simulated gastrointestinal tract conditions. In addition, the evaluation of antimicrobial activity against common foodborne pathogens and analysis of antibiotic resistance profiles of the tested Lactobacillus strains had to be performed. As a result of the study done on the project, the following significant new findings were obtained. Two novel strains viz., L. casei BI005 and strain 1LB of L. casei group were isolated from homemade traditional fermented beverage Koumiss (mare’s milk) and selected as potential probiotics because of they have beneficial properties viz., tolerance to simulating gastrointestinal tract conditions, high antimicrobial activity and lack of acquired genes of antibiotic resistance which render them promising candidates as starter cultures in the production of probiotic fermented foods. Antibiotic resistance to tetracycline were detected in 4 isolates (3 novel isolates inclusive) from three different phylogenetic groups among 16 tested Lactobacillus strains. This finding led to the opinion that lactobacilli with tetracycline resistance are common in dairy foods consumed in Kazakhstan.
MATERIALS AND METHODS
Strain isolation and culture conditions: Two Lactobacillus strains isolated previously18 were used in the study. The L. casei BI005 from homemade beverage Koumiss and L. rhamnosus BSR from industrial beverage "Curdled milk Bio-S immune+" of Food Master company, Kazakhstan (Table 3). Other 14 isolates of lactobacilli were purified from homemade and industrial dairy products (Table 1). Industrial products were consisted of yogurt of LLP «Danone Berkut» manufactured in Kazakhstan; Sourdough "Prikorm 2" from Russia company «Svoyi yogurt», composed of Lactobacillus acidophilus, L. fermentum, L. plantarum, L. casei, Streptococcus thermophilus, Bifidobacterium bifidum, Bifidobacterium breve, Bifidobacterium infantis according to the manufacturer’s prescription, cheese of JSC "Kostanai Dairy Plant DEP", Kazakhstan. Homemade products included solid consistency kurt (similar to dry cottage cheese), cottage cheese and fermented milk beverages airan (resembles yogurt), koumiss (made from mare’s milk and fermented by its microbiota) and sour cream were sampled from rural regions in suburbs of Astana and Karaganda located in Central Kazakhstan and Kostanai located in North Kazakhstan (Table 1). Approximately 2 g of kurt were aseptically crushed to powder using a porcelain mortar transferred into 2 tubes with 5 mL of De Man, Rogosa and Sharpe (MRS) broth (HiMedia Laboratories Pvt. Ltd., India) and incubated in microaerophilic conditions in BBL™ CampyPak™Plus system (BD company, USA) for 72 h at 37°C. After incubation, samples were 10-fold serially diluted in sterile saline peptone water and treated as exactly described below for other dairy samples. Ten grams of each dairy sample (beverage) was separately suspended in 50 mL of sterile di-potassium hydrogen phosphate (2% w/v) pH 7.5 at room temperature and vigorously mixed on vortex (MicroSpin FV 2400, Biosan, Latvia). Approximately 10 g of cheese and cottage cheese were homogenized separately in a porcelain mortar and then transferred into 50 mL of sterile sodium citrate solution (2% w/v) followed by vortexing. Ten fold serial dilutions of the samples in sterile saline peptone water were prepared and aliquots of 0.1 mL of these dilutions and those kurt dilutions described above were plated in duplicate on the surface of MRS agar (HiMedia Laboratories Pvt. Ltd., India) containing bromocresol green (25 mg L–1). The plates were incubated in a BBL™ CampyPak™ Plus system (BD company, USA) for 48-72 h at 37°C.
| Table 1: | Percent of cell growth of lactic acid bacteria isolates in MRS broth supplemented with 1% bovine bile |
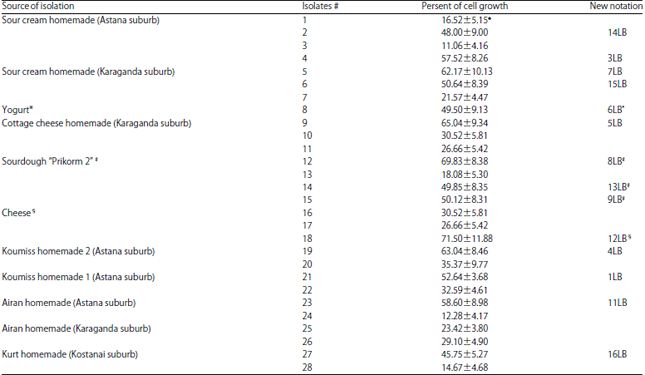 | |
♠Mean value with standard deviation (n = 3), *LLP «Danone Berkut», Kazakhstan, #Russia company «Svoyi yogurt», consisted of Lactobacillus acidophilus, L. fermentum, L. plantarum, L. casei, Streptococcus thermophilus, Bifidobacterium bifidum, Bifidobacterium breve, Bifidobacterium infantis according to the manufacturer’s prescription, §JSC "Kostanai Dairy Plant DEP", Kazakhstan | |
Different colonies of acid producing bacteria grown after incubation were determined by a yellow zone in the media around each colony, subcultured in the MRS broth and purified by plating on MRS agar. All isolates were examined with an optical microscope for Gram stain reaction and tested for catalase production. Those purified isolates classified as Gram-positive, catalase negative non-spore-forming, motionless rods were cultured on MRS broth and were stored for further analyses as lactic acid bacteria isolates. The isolates were periodically grown and stored in MRS broth at -20°C supplemented with glycerol (20% v/v). Before use, the isolates were subcultured twice in the MRS broth19.
Tolerance of lactic acid bacteria isolates to bile: The bile tolerance of tested isolates was investigated according to Vinderola and Reinheimer20. Each subcultured twice isolate of our study was inoculated (2% v/v) into MRS broth with 1.0% of bovine bile B-8381 (Sigma-Aldrich, USA) containing a mixture of conjugated CBS-FBS bile acids. All tubes were thoroughly stirred and incubated for 24 h at 37°C. After cultivation, the optical density at 560 nm was measured and compared to a control culture (without bile acids). The results were expressed through the percentage of cell growth in the presence of bile acids compared with the control. The reported data of percent of cell growth in Table 1 are mean values after three independent tests (n = 3) with standard deviation calculated using Excel software (Microsoft Corporation, USA).
Identification of lactic acid bacteria isolates by nucleotide sequence analysis of 16S rRNA gene: Bacterial DNA of each unidentified testing isolate was extracted using a set of Wizard® Genomic DNA Purification Kit («Promega», A11125) as recommended by the manufacturer. Quality of genomic DNA was monitored by 1.0% agarose gel electrophoresis. Quantitative DNA analysis was carried out using a Nano Drop ND 1000 spectrophotometer. The isolates were identified to genus and species grouping level by PCR (polymerase chain reaction) using 16S rRNA gene sequencing with the universal primers 8F (5’–AgAgTTTgATCCTggCTCAg-3’) and 806R (5’–ggACTACCAgggTATCTAAT–3’)21. The PCR reaction was performed in a total volume of 30 μL. The PCR mixture contained 25 ng of DNA, 1 U Maxima Hot Start Taq DNA Polymerase, 0.2 mM of each dNTP, 1x PCR buffer (Fermentas, Lithuania), 2.5 mM MgCl2, 10 pmol of each primer. The PCR amplification was carried out on Thermocycler GeneAmp PCR System 9700 (Applied Biosystems, USA) programmed as follows: Long denaturation 95°C for 7 min, followed by 30 cycles comprising the successive temperature and time regimes as (1) 95°C for 30 sec, (2) 55°C for 40 sec, (3) 72°C for 50 sec and final elongation during 7 min at 72°C. Purification of PCR products was carried out by enzymatic method22 using exonuclease and alkaline phosphatase (Fermentas, Lithuania). Sequencing reaction was carried out by using BigDye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, USA) according to manufacturer’s instructions, followed by separation of the fragments on automated Genetic Analyzer 3730×l DNA Analyzer (Applied Biosystems, USA). The fragments of 16S rRNA gene of the studied 14 cultures were analyzed and joined together into a common contig using software SeqScape 2.6.0 (Applied Biosystems, USA). The obtained sequences were aligned to 16S rRNA gene sequences in the GenBank database using the BLAST algorithm.
Tolerance of Lactobacillus strains to simulated gastrointestinal tract conditions: Lactobacilli tolerance to stress factors in simulated GI tract conditions was investigated in a two-step assay according to Haller et al.23 with slight modifications. Briefly, tested Lactobacillus strains were cultured overnight in MRS broth at 37°C and settled by centrifugation 5,000×g for 5 min, washed twice in phosphate-buffered saline (PBS, pH 7.2) and adjusted to 2 McFarland (A600 = 0.451) namely 6.0×108 CFU mL–1 in PBS. In the triplicate assays 0.1 mL aliquots of adjusted bacterial suspension of each strain was transferred to test-tubes with 0.9 mL of MRS broth (Control for two-step treatment) and test-tubes with 0.9 mL of hydrochloric acid (pH 3.0) and after thorough stirring incubated at 37°C for 1 h. After incubation aliquots (0.1 mL) of this mixture were transferred to test-tubes with 0.9 mL of MRS broth supplemented with 0.3% of bovine bile B-8381 thoroughly stirred and incubated for another 1 h at 37°C. The control test tubes were continued to remain in the thermostat simultaneously with bile test tubes. After incubation the aliquots (0.1 mL) from control and test-tubes were taken for enumeration of survived cells. Enumeration was carried out by 10-fold serial dilutions with sterilized PBS pH 7.2 in microcentrifuge tubes. The appropriate dilution factor was determined and serial dilutions of the culture were plated in duplicate on MRS agar then incubated in microaerophilic conditions in a BBL™ CampyPak™ Plus system (BD company, USA) for 48 h at 37°C. After incubation bacteria colonies were counted and the number of lactobacilli was calculated according to the standard ISO 1521424. The reported data are mean values of a percentage of survival after three independent assays with standard deviations calculated using Excel software (Microsoft Corporation, USA). Percentage of survival was calculated as follows:
Percentage of survival = Nt/Nc×100 | (1) |
| Nt | = | log CFU mL–1 after two-step treatment |
| Nc | = | log CFU mL–1 (Control) |
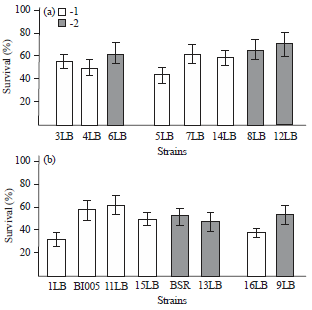 | |
| Fig. 1(a-b): | Tolerance of the Lactobacillus strains to simulated GI tract conditions, 1: Homemade products Lactobacillus strains, 2: Industrial products Lactobacillus strains, error bars represent standard deviations (n = 3) |
Estimation of statistical significance of the differences between the Lactobacillus strains on percentage of survival (Fig. 1) was carried out by using the overlap rule for SE bars25.
Indicator organisms: Gram negative pathogens Escherichia coli 157-B-RKM-0040, Salmonella typhimurium 0162, Gram positive pathogens Staphylococcus aureus 209 P-B-RKM–0057 as well as yeast pathogen Candida albicans ATCC 885-653 were obtained from Central Museum of the RSE "Republican Collection of Microorganisms". Indicator organisms except for C. albicans ATCC 885-653 were routinely grown to stationary phase in the nutrient broth medium (HiMedia Laboratories Pvt. Ltd., India) at 37°C under aerobic conditions. The C. albicans ATCC 885-653 was subcultured and grown in the sabouraud broth (HiMedia Laboratories Pvt. Ltd., India) at 37°C under aerobic conditions. Before experiment the overnight cultures were adjusted to uniform concentrations of 1.0×107 CFU mL–1 of bacteria by spectroscopy (OD600) and maintained under refrigerated conditions at 4°C19.
Detection of antimicrobial activity: For detection of antimicrobial activity of Lactobacillus strains the deferred antagonism procedure of Tagg et al.26 with slight modifications was used. Overnight cultures were adjusted to uniform concentrations of bacteria by spectroscopy (OD600 = 1.0), spotted onto the surface of MRS agar plates with microbiological loop holder (10 μL) and incubated in microaerophilic conditions for about 48 h at 37°C. After incubation the Lactobacillus strains were kept in chloroform vapor for 30 min and aired for 20 min. This step is necessary for killing cultures, in order to cease their growth which could mask inhibition zones of indicator cultures. Then 7 mL aliquots of soft nutrient agar (containing 0.7% agar) were seeded with 1 mL of appropriate indicator strain (1.0×107 cells), poured over the treated plates and allowed to solidify. In the case of C. albicans 7.0 mL of soft sabouraud dextrose agar instead of nutrient agar was used. After overnight aerobic incubation at 37°C the plates were checked for the formation of inhibition zones around the producer colonies. Inhibition zones diameters (IZD) were measured in millimeters (Table 3). For convenience the length of zones of inhibition were divided into 5 intervals (Table 3) for Gram negative pathogens and 3 intervals (Table 3) for Gram positive pathogen. In case of antifungal activity against C. albicans length of inhibition zones were divided into two intervals according to Okkers et al.27 (Table 3). Three independent experiments were carried out with each Lactobacillus strain and mean values were calculated followed by standard deviation determination by Excel Program (Microsoft Corporation, USA).
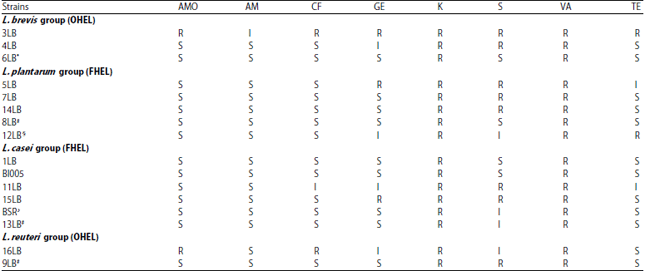
Analysis of antibiotic resistance profile: Antibiotic resistance of Lactobacillus strains was tested by agar disk diffusion assay according to the CLSI28 with the following modifications. Overnight Lactobacillus cultures were adjusted to uniform concentrations of 1.0×107 CFU mL–1 of bacteria by spectroscopy (OD600). The entire volume of 100 μL of standardized bacterial suspension was streaked onto MRS agar using a sterile cotton swab until homogeneity. Then commercial antibiotic disks (HiMedia, India) were placed onto the overlaid plates. The plates were then incubated in microaerophilic conditions at 37°C for 24 h. Zones of growth inhibition around the disks were measured in millimeters and used to assess antibiotic resistance based on the cut-off values proposed by Charteris et al.29 for lactobacilli. The result of 3 independent experiments (mean values) was expressed as sensitive (S), resistant (R) or intermediate (I) in Table 4. Antibiotic resistance profiles of tested strains were obtained using amoxicillin (AMO, 10 μg), ampicillin (AM, 10 μg), cefazolin (CF, 30 μg), gentamycin (GE, 10 μg), kanamycin (K, 30 μg), streptomycin (S, 10 μg), vancomycin (VA, 30 μg) and tetracycline (TE, 30 μg). To check the properties of the antibiotic discs 2 reference microorganisms Escherichia coli ATCC 25922 В-RKM 0447 and Staphylococcus aureus ATCC 25923 В-RKM 0682 (Central Museum of Microorganism Cultures of RSE "Republican Collection of microorganisms", Kazakhstan) were tested for susceptibility to the respective antibiotics.
RESULTS
Strain isolation and culture conditions: As a result of isolation, 52 cultures of Lactic Acid Bacteria (LAB) were obtained. According, to the cultural, morphological and physiological features these cultures were Gram-positive, non-spore-forming, motionless, catalase negative rods (28 cultures) and Gram-positive, non-spore-forming, motionless, catalase negative cocci (24 cultures). Twenty eight rod-shaped LAB were chosen for further study (Table 1).
Tolerance of lactic acid bacteria isolates to bile acids: In Table 1 percent of cell growth of lactic acid bacteria isolates after 24 h exposure at 1.0% of bile acids are presented. As it turned out microbes grew differently in the presence of bile acids. Percent of cell growth varied from minimal value of isolate# 3 equal to 11.06% to maximal value of isolate# 18 equal to 71.5%. Several most tolerant LAB isolates showed percentage of growth ranging from 40-71%, for example, numbers: # 2, # 4, # 5 etc and were combined in group of most tolerant LAB. Some isolates viz., # 3, # 24 (12.28%), # 28 (14.67%), # 1 (16.52%) and # 13 (18.08%) amounted group of least tolerant to bile isolates. Remaining isolates amounted group with intermediate tolerance from 20-40% of cell growth (# 7, # 11, etc.).
Identification of LAB isolates by nucleotide sequence analysis of 16S rRNA gene: The results of phylogenetic analysis of fourteen LAB isolates with highest tolerance to bile acids were obtained and illustrated in Table 2. According, to conducted identification isolates were placed into 4 phylogenetic groups. The isolates designated as: "Old notation/new notation" namely # 4/3LB, # 19/4LB and # 8/6LB got into phylogenetic group named L. brevis group. Isolates numbered as: # 2/14LB, # 5/7LB, # 9/5LB, # 12/8LB and # 18/12LB got into L. plantarum group. Isolates numbered as: # 21/1LB, # 23/11LB, # 6/15LB and # 14/13LB got into L. casei group. Isolates numbered as: # 27/16LB and # 15/9LB got into L. reuteri group.
Tolerance of Lactobacillus strains to simulated gastrointestinal tract conditions: Figure 1 illustrates the results of two-step assay procedure. The value of tolerance to simulated gastrointestinal tract conditions expressed as percent survival (72±10%) for industrial strain 12LB§ in L. plantarum group was the highest. Other strains of this phylogenetic group were close to the 12LB§ by the value of the tolerance with the exception of strain 5LB (44±5%). The strain 1LB of L. casei group had the lowest value of percent survival (35±6%) and the strain 11LB had the highest value of this parameter in L. casei group (63±8%). As for the remaining strains of this group they had close to strain 11LB values of percent survival. In the L. reuteri group strains 16LB and 9LB# had values of percent survival equal to (44±4%) and (56±8%), respectively.
Detection of antimicrobial activity: In Table 3 isolates of all phylogenetic groups except for L. reuteri group had wide scope in size of inhibition zone diameters determining antimicrobial activity (from + to ++++) against Gram-negative indicator pathogens E. coli 0040 and S. typhimurium 0162 and less range (from + to +++) against Gram-positive pathogen S. aureus 209p. The recorded inhibition zone diameter (IZD) for tested Lactobacillus strains ranged from 9.7 mm (5LB) to 35.2 mm (4LB) against E. coli 0040 and from 10.3 mm (12LB§) to 32.7 mm (3LB) against S. typhimurium 0162 (Table 3). Regarding Gram-positive food pathogen S. aureus 209p tested isolates showed IZD ranged from 7.4 mm (9LB) to 20.7 mm (8LB).
Isolates from L. brevis and L. reuteri group present obligate heterofermentative lactobacilli (OHEL) and isolates from two other groups present facultative heterofermentative lactobacilli (FHEL). In Lactobacillus strains 3LB, 8LB# and 11LB representing three phylogenetic groups anti-Candida activity was found (Table 3). Three Lactobacillus strains 5LB, 14LB and 12LB§ representing L. plantarum group had low antimicrobial activity against indicator pathogens and Lactobacillus strains 9LB# and 16LB of L. reuteri group, had moderate and weak antimicrobial activities against Gram-negative indicator pathogens.
Analysis of antibiotic resistance profile: Antibiotic resistance profile of Lactobacillus strains was obtained by disk diffusion method (Table 4). Growth inhibition zones around the disks with glycopeptide vancomycin and aminoglycoside kanamycin were not observable for all tested strains. On the left part of the Table 4 susceptibility of tested strains to antibiotics of β-lactam group (amoxicillin, ampicillin and cefazolin) is presented. Except for the strains 3LB, 11LB and 16LB all other strains were susceptible to β-lactams. Strain 3LB in group L. brevis was notable in that showed resistance to all tested antibiotics including intermediate resistance to ampicillin. The rest of the strains were susceptible to β-lactams, except for strain 11LB (L. casei group) carried intermediate resistance to cefazolin and strain 16LB (L. reuteri group) carried resistance to amoxicillin and cefazolin (Table 4). Most cultures were susceptible to amoxicillin (14/16) and cefazolin (13/16). In the centre and right part of the Table 4 total resistance to glycopeptide vancomycin and aminoglycoside kanamycin and different variants of susceptibility and resistance (S, I and R) of the tested strains to antibiotics of aminoglycoside group (gentamicin and streptomycin) and tetracycline are presented. None of the strains was susceptible to all antibiotics. Moreover strain 3LB was not susceptible to any of the 8 antibiotics. Strains 11LB and 16LB were susceptible to two antibiotics. Strain 5LB was susceptible to 3 antibiotics. The most sensitive strains namely L. casei В I005, 6LB, 1LB and 8LB were susceptible to 6 antibiotics. Percent of susceptibility to GE and S of all tested isolates was 56 and 25%, respectively. Antibiotic resistance of the novel strain 3LB and industrial strain 12LB§ to tetracycline and intermediate resistance of two novel strains 5LB and 11LB to tetracycline were detected.
| Table 2: | Identification of lactic acid bacteria isolates by nucleotide sequence analysis of 16S rRNA genes |
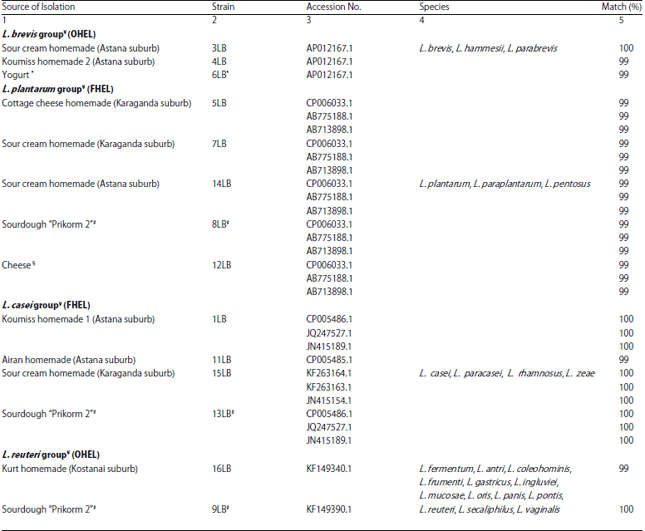 | |
¥ Phylogenetic grouping according to Felis and Dellaglio32, *LLP «Danone Berkut», Kazakhstan, #Russia company «Svoyi yogurt», consisted of Lactobacillus acidophilus, L. fermentum, L. plantarum, L. casei, Streptococcus thermophilus, Bifidobacterium bifidum, bifidobacterium breve, Bifidobacterium infantis according to the manufacturer’s prescription, §JSC "Kostanai Dairy Plant DEP", Kazakhstan | |
| Table 3: | Antimicrobial activity of Lactobacillus strains |
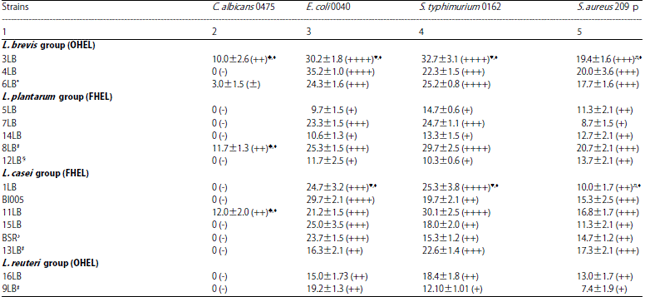 | |
♣Inhibition zone: 5-10 mm (+), inhibition zone: 10–15 mm (++) according to Okkers et al.27, ♥Inhibition zone: 0 mm no activity, <5 mm: Doubtful (±), 5-15 mm: Weak (+), 15-20 mm: Moderate (++), 20-25 mm: Strong (+++), ≥25 mm: Very strong (++++), ♬inhibition zone: 5-10 mm (+), 10-15 mm (++), ≥15 mm (+++), ♦Mean values with standard deviation (n = 3), *Yogurt of LLP «Danone Berkut», Kazakhstan, #Sourdough "Prikorm 2" of Russia company «Svoyi yogurt», consisted of Lactobacillus acidophilus, L. fermentum, L. plantarum, L. casei, Streptococcus thermophilus, Bifidobacterium bifidum, bifidobacterium breve, Bifidobacterium infantis according to the manufacturer’s prescription, §Cheese of JSC "Kostanai Dairy Plant DEP", Kazakhstan, ♪Curdled milk Bio-S Imune +" of Food Master company, Kazakhstan | |
| Table 4: | Antibiotic resistance profiles of Lactobacillus strains |
 | |
*Yogurt of LLP «Danone Berkut», Kazakhstan, #Sourdough "Prikorm 2" of Russia company «Svoyi yogurt», consisted of Lactobacillus acidophilus, L. fermentum, L. plantarum, L. casei, Streptococcus thermophilus, Bifidobacterium bifidum, bifidobacterium breve, Bifidobacterium infantis according to the manufacturer’s prescription, §Cheese of JSC "Kostanai Dairy Plant DEP", Kazakhstan, ♪"Curdled milk Bio-S Imune +" of Food Master company, Kazakhstan | |
DISCUSSION
Samples were prepared from homemade and industrial dairy products purchased in Kazakh markets. Strain isolation and culture conditions were performed by standard procedures19.
The results in Table 1 regarding the most tolerant group are in good agreement with the data of Horackova et al.30 who evaluated the bile tolerance after 24 h exposure of selected lactobacilli to 1% bile acids in comparison with the commercial probiotic strain. They found that all lactobacilli tested had the ability to adapt in the environment of bile salt. This adaptability was the least in the case of commercial probiotic strain L. casei LAFTI L26, where only 40% growth was detected, compared to the lack of bile control. In contrast, the other strains reached about 75% growth as compared to the control sample lack of bile30. Similar percentages of cell growth under 12 h exposures with bile concentration equal to 1% were obtained by Hyacinta et al.31 Their findings showed tolerance to 1% bile as survival percent of tested lactobacilli and had the following values: L. murinus C 96%, L. mucosae D 35%, L. reuteri E 85%, L. reuteri KO4b 98%, L. reuteri KO4m 40%, L. reuteri KO5 50%, L. plantarum KG1z 60%, L. plantarum KG4 50%, L. reuteri CCM3625 55%. Only L. mucosae D and L. reuteri KO4m fall within the group with intermediate tolerance.
In a study of Belicova et al.8, 11 L. plantarum strains isolated from cheese bryndza showed percent of cell growth varied in the range from 2-19% after 24 h incubation of microbes in a medium with 1% of bile acids at 37°C. Values of tolerance determined in present study for LAB isolates from group with least tolerance to bile (Table 1) are in good agreement with findings of Belicova et al.8 According to the findings of several investigators bile tolerance is a strain-specific trait and bile tolerance of species cannot be generalized6. In cited above studies there was extreme variability in tolerance that was found within a species or genus. Thus, findings of present study highlighted a noticeable heterogeneity in the ability of the strains of lactic acid bacteria to withstand harsh growth conditions with bile acids and are in good agreement with cited studies. To identify the most promising candidates for technological uses 14 isolates of LAB among 28 isolates viz., # 2, # 4, # 5, # 6, # 8, # 9, # 12, # 14, # 15, # 18, # 19, # 21, # 23 and # 27 with tolerance as percent of cell growth higher than 40% were selected for further experiments characterizing probiotic properties.
For the 14 LAB isolates with highest tolerance to bile acids (Table 1) preliminary phylogenetic identification was carried out by determining the nucleotide sequence of 16S rRNA gene fragment, followed by determination of nucleotide identity with the sequences deposited in the international data base GenBank (Table 2). Phylogenetic grouping was carried out according to Felis and Dellaglio32 and resulted to 4 phylogenetic groups. Taking into account the peculiarities of bacteria species assessment on the basis of identity of a fragment of 16S rRNA gene, 14 strains of the study were identified only provisionally and unambiguous identification to the species level requires additional study33. However, the results of molecular analysis provided a good opportunity for probiotic properties comparison with analogs isolated in other studies.
Metabolic and functional properties of Lactobacillus isolates of present study were evaluated in the two-step assay procedure which meant measuring of their ability to tolerate acid and bile rich environment in simulated gastrointestinal tract conditions of a human organism23. Such two-step approach with certain approximation simulates the conditions of stress in the stomach and upper intestine which are usually faced by lactobacilli during their moving in the digestive tract. It should be noted that cell surface of Lactobacillus strain protects it from the action of acids (low pH). Reduction of protons penetration occurs via changes in the structure of the cell membrane or cell wall34. Intracellular acidification may reduce the activity of acid-sensitive enzymes and cause damage to DNA and proteins35. Regarding the changes of metabolic and functional properties of lactobacilli in bile toxicity, which appears to occur within the second step of this assay, there was evidence that some genes are induced by the action of bile acids and involved in biosynthesis of membrane, peptidoglycan and surface proteins36.
There was an opportunity to compare the results for strains of L. plantarum and L. casei groups with data of Haller et al.23, who studied tolerance of strains of L. paracasei, L. plantarum and L. sakei to the consecutive exposure to hydrochloric acid (pH 2.5 for 1 h) and cholic acid (10 mM for 1 h). Recalculation of final concentration of cholic acid from cited study into percentage units resulted to a value equal to 0.4% which was close to 0.3 % of bovine bile used in present study. Probiotic strain L. johnsonii La1 in cited work was taken as a reference strain. For comparison purposes the percentage of survival of the strains in cited paper was calculated via the formula (1) Given in paragraph # 2.4 in present study. Calculated percent of tolerance for the reference probiotic strain L. johnsonii La1 was equal to 91%. Percent survival for L. plantarum strains numbered as # 2963, # 2966 and # 2833 in the study, was 87, 92 and 77% respectively. These values were close in magnitude to values of percent survival of the strains in L. plantarum group except for strain 5LB (44±5%). Strain 5LB was statistically different (p<0.05) from the strain 12LB (72±10%) on this parameter. Strain 1LB of L. casei group had the lowest value of percent survival (35±6%) and with p<0.05 survived worse than the strain 11LB (63±8%). On the other hand in L. reuteri group for strains 16LB (44±4%) and 9LB (56±8%) there was no difference (p>0.05) in the values of percent survival. Thus, exposing these isolates to simulating GI-tract conditions, including, low pH = 3.0 and 0.3% concentration of bile revealed nearly the same responses and only two out of the 16 isolates showed about 2 times less tolerance (p<0.05) to the stress factors of GI-tract simulating conditions. Basically all other strains of present study showed approximately the same percentages of tolerance within the measurement errors and most likely would be alive in GI tract in vivo.
Producing antibacterial and antifungal substances, such as short-chain fatty acids, ethanol, hydrogen peroxide, cyclic dipeptides, pyroglutamic acid, bacteriocins, etc., is very important probiotic property. These substances may enhance the ability of lactobacilli to compete against other GI microbes and could potentially inhibit pathogenic (disease-causing) bacteria37 and fungy38-43. Lactic acid is a key antimicrobial compound synthesized in lactobacilli34. Positive correlation between the accumulation of lactic acid and a strong antimicrobial activity in L. rhamnosus GG against S. typhimurium was found by De Keersmaecker et al.44 and approved by several authors7,45. Lactic acid performs its activity via lowering the pH and via its undissociated form as the outer membrane permeabilizer in Gram-negative bacteria allowing other compounds to enter into the synergistic relationship with lactic acid34. In addition, organic acids and lactic acid, among them can capture elements necessary for growth, such as iron, because of their chelating properties34. In Table 3 antimicrobial activity of L. rhamnosus BSR♪ against S. typhimurium 0162 was equal to 15.3 mm (IZD). Haghshenas et al.46 used other method, viz., modified well-agar diffusion assay and found for several L. rhamnosus strains the values of IZD against S. typhimurium equal to 12.8, 16.7, 13.3 and 15.3 mm. Apparently antimicrobial activity of L. rhamnosus BSR♪ was caused by action of secreted lactic acid similar to study of De Keersmaecker et al.44. However, findings of Haghshenas et al.46 may be caused by other secreted substances (hydrogen peroxide and bacteriocin etc.) because the antimicrobial effect caused by acid was neutralized by sodium alkaline. Inside L. casei group (Table 3) there were more active strains whose antimicrobial activities may be caused by more intensive secretion of short-chain fatty acids or synthesis of other substances (bacteriocin and hydrogen peroxide etc.). Similar assumption about the nature of antimicrobial activity may be proposed for members of other phylogenetic groups in Table 3 against all three indicator pathogens. Thus antimicrobial activities of the lactobacilli against Gram-negative or Gram-positive pathogens were very variable and strain-specific (Table 3).
Antimicrobial activity against S. aureus of facultative heterofermentative lactobacilli (FHEL) in the present study varied from minimal value of 8.7 mm (isolate 7LB) to maximal value of 20.7 mm (for isolate 8LB#). Mean value of IZD equal to 13.9 mm found for antimicrobial activity of 11 members from FHEL group in this study was close to mean value of IZD equal to 15.9 mm obtained by Annuk et al.45 for 13 strains of FHEL group. Assay in cited work was carried out in microaerobic conditions by a streak line procedure on plates containing MRS medium, which is vaguely reminiscent of the method of the present study. Interestingly mean value of IZD for 5 strains of FHEL group equal to 12.0 mm (i.e., a bit less than mean value of IZD equal to 13.9 mm found in present study) was obtained by Haghshenas et al.46 who used other assay.
Mean values of antimicrobial activity against S. aureus for Lactobacillus strains of obligate heterofermentative group (OHEL) were assessed as 15.5 mm (5 strains) in present study and 23.4 mm (18 strains) in study of Annuk et al.45 Detected differences in activity may be associated with different numbers of tested strains in 2 compared studies or difference in sample and indicators loading in 2 assays etc. Alternatively Annuk et al.45 studied intestinal Lactobacillus strains isolated from the faecal samples of 1-24-month-old healthy infants, which perhaps had higher antimicrobial activity.
In a study of Rushdy and Gomaa7 antibacterial activity was hypothetically associated with bacteriocin-like substance and then this hypothesis was confirmed by extraction the bacteriocin-like protein. Bacteriocins produced by Gram-positive bacteria usually possess little or no activity against Gram-negative pathogens37. So the values of activity (IZD) against Gram-negative and Gram-positive indicator cultures were the same or the latter was more than the former. For example in a study of Rushdy and Gomaa7 antimicrobial activity of L. brevis was found as values of IZD equal to 7.0, 6 and 8.3 mm against E. coli, S. typhimurium and S. aureus respectively, i.e., L. brevis were more effective against Gram-positive pathogen. The same conclusion about more effective activity against Gram-positive pathogen follows from study of Al-Madboly and Abdullah47 for L. plantarum strains isolated from homemade fermented milk "Yogurt". Higher inhibition zone diameters ranging from 36-44 mm against vancomycin-resistant virulent S. aureus isolate and maximal IZD equal to 35.0 mm against E. coli and 28.0 mm against S. typhimurium were recorded and indicated the more effective activity against Gram-positive pathogen47. Trend of superiority or equality of antimicrobial activity against Gram-positive pathogens in comparison with the Gram-negative ones found7,47 also was found for some isolates, viz., 4LB (against S. typhimurium), 5LB, 14LB, 12LB§, L. rhamnosus BSR♪ (against S. typhimurium), 13LB# and 16LB (against E. coli) of present study. Based on the mentioned above concept implying that bacteriocins produced by Gram-positive bacteria usually possess little or no activity against Gram-negative pathogens37 and findings of cited studies7,47 it can be hypothesized that listed Lactobacillus strains of present study have a great chances of being producers of bacteriocins.
Thus, in present study, 9 out of 16 isolates, viz., strains 3LB, 4LB, 6LB*, 7LB, 8LB#, 1LB, L. casei BI005, 11LB, 13LB# (56%) showed strong antimicrobial activity against at least 2 food-borne pathogens either Gram-positive or Gram-negative. Among them 5 isolates (31%) carried strong antimicrobial activity against all 3 indicator pathogens (3LB, 4LB, 6LB*, 8LB# and 11LB) and among these 5 potent isolates three isolates (19 %) namely 3LB, 8LB# and 11LB had a broad antimicrobial spectrum because they performed potent antifungal activity against Candida albicans additionally to strong antimicrobial activities against Gram-positive and Gram-negative pathogens (Table 3). Anti-fungal activities in L. brevis strains were studied by Tropcheva et al.39 and Falguni et al.40, in L. casei strains by Hassan and Bullerman41 and Delavenne et al.42 in L. plantarum strains by Sharma and Srivastava38 and Wang et al.43.
Thus, according to tested properties (acid and bile tolerance, antimicrobial activities (Table 1, 3, Fig. 1) novel strains 3LB, 4LB from L. brevis group, 7LB from L. plantarum group, 1LB, L. casei BI005, 11LB from L. casei group and industrial food strains 6LB* from L. brevis group, 8LB# from L. plantarum group and 13LB# from L. casei group are promising probiotic strains with potent antimicrobial activity provided they are safety to consumption. Due to the fact that they produce antibacterial and antifungal substances they may be beneficial sources for extraction of such substances that might be helpful in the fields of food and pharmaceutical industry.
Intrinsic susceptibility of the most Lactobacillus strains in Table 4 toward the inhibitors of cell wall synthesis, such as amoxicillin, ampicillin, cefazolin and protein synthesis inhibitor, such as tetracycline, streptomycin and gentamicin was in accordance with bibliography data9-13. Antibiotic resistance profile of strain 16LB in L. reuteri group shows resistance to the amoxicillin and cefazolin and susceptibility to other β-lactam ampicillin. Apparently, such resistance is not associated with the activity of β-lactamase, otherwise the β-lactamase also would have to show resistance of strain 16LB to ampicillin. The same assumption was made by Botina et al.48 who analyzed antibiotic resistance profiles of the human gastrointestinal microbiome Lactobacillus spp. They have found that two L. plantarum strains were sensitive to ampicillin but were resistant to β-lactam antibiotic benzyl penicillin. A possible explanation for these results follows from study of Rosander et al.49 who found that the β-lactam resistance of L. reuteri ATCC 55730 is caused by a number of point mutations in the genes encoding "housekeeping" penicillin binding proteins Pbp1a, Pbp2a and/or Pbp2x and that this resistance can be regarded as non-transmissible. A similar scenario could take place in case of inhibition of strain 11LB from L. casei group, which carried intermediate resistance to cefazolin and susceptible towards amoxicillin and ampicillin (Table 4). Antibiotic resistance profile to β-lactams of strain 3LB in L. brevis group was different from considered above profiles of strains 11LB and 16LB because all three drags (AMO, AM and CF) did not inhibit a growth of strain 3LB. Therefore, in this case inactivation of antibiotics via chemical modification by β-lactamase or induction of penicillin binding protein acting as a competitive to PBP peptidases target cite was possible. For the both mechanisms transmissible resistance determinants have been identified as ampC and mecA, respectively49. However, in order to unambiguously determine the mechanism of antibiotic resistance, i.e., whether it is intrinsic or acquired further molecular analysis is required.
In this study, vancomycin-resistant phenotype had been obtained for all tested isolates (Table 4). These results are in good agreement with the accepted opinion that vancomycin-resistant phenotype is internal resistance13.
Traditionally resistance to aminoglycoside antibiotics was considered to be intrinsic in LAB13 and is attributed to the absence of cytochrome-mediated electron transport, which mediates drug uptake. But now these findings need to be clarified. It was shown that the lactic acid bacteria were intrinsically resistant to gentamycin (71.1%) and to streptomycin (73.3%) when the Minimal Inhibition Concentration (MIC) breakpoints of SCAN or FEEDAP were used. However, more than 80 and 60% of the strains were "transformed" to susceptible phenotype either to gentamycin or streptomycin, respectively, when the MIC breakpoints of Danielsen and Wind were used50. The specification of resistance-susceptibility MIC breakpoint values is a problem with lactic acid bacterial antibiotic resistance determination as mentioned in Introduction section. The same problem exists for specification of cutoff points in agar disk diffusion methods used in present study. Due to the study of Mayrhofer et al.51 it is possible to compare the results of two methods viz., the broth microdilution (MIC) and agar disk diffusion assays. In very few cases the results of the broth microdilution (MIC) and agar disk diffusion methods (cutoff) do not coincide52. In Table 4 resistance phenotype for kanamycin was found for all tested Lactobacillus isolates by agar disk diffusion assay and these results are in good agreement with study of Cano Roca52 who tested similar species of lactobacilli by the same assay. But when MIC of kanamycin by the broth microdilution assay was measured for the same isolates the opposite results were obtained, which testified to the antibiotic sensitivity of lactobacilli to kanamycin52. This discrepancy may be linked to the problems discussed and deserves further study.
In Table 4 percent of strains with antibiotic resistance/intermediate resistance to streptomycin in L. plantarum/L. pentosus group was equal to 80.0%. Munoz et al.53 showed that 83% of L. pentosus strains isolated from green table olives were resistant to streptomycin (MIC>256 μg mL–1). The L. pentosus strains showed bimodal MIC distribution for streptomycin, which distinguished between sensitive (MIC range 8-128 μg mL–1) and resistant (MIC>256 μg mL–1) subpopulations53. Such resistances resulted from an intrinsic mechanism of non-transmissible resistance determinants, which may in part be due to activity of chromosomally encoded efflux pumps (NorA, MepA and MdeA)53. In Table 4 resistance of members in L. casei group towards protein synthesis inhibitors streptomycin and gentamicin is something in common with the findings of Klare et al.54 who found low values of MIC of gentamicin and streptomycin to lactobacilli and yet identified 2 strains of L. rhamnosus L-015 and L-455 and one of L. paracasei L-005 with high-level resistance to streptomycin with MICs of ≥2048 mg L–1. The gene aad(E) responsible for high-level of streptomycin resistance was not detected in these 3 Lactobacillus isolates54. Apparently, the high resistance to streptomycin was associated with the intrinsic mutations similar to those studied by Kiwaki and Sato55 and Sato and Iino56. These researchers found that a high resistance to streptomycin was correlated with a mutation on the rpsL gene of ribosomal protein S12 in B. bifidum and B. breve55,56. Percent of L. casei group strains susceptible to streptomycin were 33% and to gentamicin were 66% (Table 4). The latter value was close to the findings of Liu et al.57, according to whom 70% of Lactobacillus isolates from foods and drug additives were susceptible to gentamycin. However, all of them was susceptible to high streptomycin levels (0.3 mg per filter) not supporting the intrinsic resistance of Lactobacilli to this antibiotic57.
Members of OHEL group (Table 4) also had both resistant and susceptible strains to streptomycin and gentamicin and percents of susceptible strains were equal to 20 and 40%, respectively.
Thus, analysis of bibliography data and findings of present study showed that there is a problem in determination of intrinsic resistance of lactobacilli to streptomycin and gentamicin and it depends on correct determination of MIC breakpoints even for each species of lactobacilli. Therefore, isolates of present study with resistance to streptomycin and gentamicin need further study in order to figure out, which among three possible mechanisms of lactobacilli resistance to these aminoglycosides were realized, viz., (a) Reduced uptake or decreased cell permeability53, (b) Alterations at the ribosomal binding sites55,56 and (c) Production of modifying enzymes58. Resistance to streptomycin can occur by alterations at the ribosomal binding sites since streptomycin binds to a single site on the 30S ribosomal subunit55,56. Resistance to gentamicin can occur by the most common type of aminoglycoside resistance, viz., enzyme modification. Therefore, participation of acquired genetic determinant such as aac (6’)-aph (2’’) in resistance to gentamicin cannot be ruled out58.
Relying on the literature data9,11-13,15,16 the lactobacilli are usually susceptible to tetracycline. It is very possible that the observed resistance to tetracycline in Table 4 was an acquired resistance horizontally transmissible by mobile genetic elements15,16. Like in study of Nawaz et al.16 the tetracycline resistance was found in strain 3LB (R) from L. brevis and in strain 5LB (R) and 12LB§ (I) from L. plantarum groups (Table 4). In other study the tet(M) gene was detected also in a conjugative transposon Tn916 in a food-borne strain of L. paracasei15. In present study an intermediate resistance of the strain 11LB (I) of L. casei group was also found (Table 4). The tetracycline resistance/intermediate resistance of 4 strains belonging to 3 different phylogenetic groups was identified among 16 lactobacilli therefore antibiotic resistance to tetracycline is apparently well dispersed in lactobacilli of Kazakh dairy foods.
Acquired antibiotic resistance may be controlled by removal/deletion of such mobile genetic elements carrying the resistance genes15. This "Curative strategies" has been applied to the probiotic L. reuteri DSM ATCC 55730 which harbored two plasmids carrying tet(W) tetracycline and lnu(A) lincosamide resistance genes, respectively49. After appropriate irreversible removal of plasmids daughter cells retained probiotic properties of the parent strain49. Treatment with acriflavine resulted in an irreversible loss of the plasmid15. Protoplast formation technique also confirms the loss of plasmid leading to absence of the resistance gene in the strain59. Such strains after the irreversible removal of plasmids or transposons can be used in different probiotic food preparations because of their inability of resistance gene transfer. Besides in study of El-Deeb et al.60 the ability of cellular extract of LAB to cure plasmid acquiring resistance in certain clinical antibiotic-resistant bacteria isolates was investigated. The extract mediated plasmid curing resulted in the subsequent loss of antibiotic resistance encoded in the plasmids. These may be new direction in controlling spread of plasmid mediated antibiotic resistance60.
In continuation of the study, it would be good to figure out a mechanism of antibiotic resistance of novel strains 3LB, 4LB, 7LB, 11LB and industrial product strains L. rhamnosus BSR♪, 13LB# in order to make decision whether resistances to β - lactams, streptomycin and gentamycin were intrinsic or acquired/transmissible. In a latter case "Curative strategies" might be applied.
As a result of the study done on the project in the part where the influence of stress factors (bile, acid) on the lactic acid bacteria was studied the following significant findings were obtained. Noticeable heterogeneity was found in the ability of isolates of lactic acid bacteria from Kazakh traditional fermented dairy foods to withstand harsh growth conditions at 1% bile acids. Bile tolerance with values more than 40% of cell growth was obtained for 14 among 28 isolates of lactic acid bacteria and these 14 LAB isolates were selected for implementation of the project tasks. Consecutive exposure of tested Lactobacillus strains to hydrochloric acid (pH 3.0 for 1 h) and bile acids (0.3% for 1 h) as a simulation of gastrointestinal tract conditions revealed almost the same responses and only 2 isolates showed about 2 times less tolerance (p<0.05) to the stress factors. These results led to the conclusion that all 16 tested Lactobacillus strains most likely would be alive in gastrointestinal tract in vivo and could withstand some harsh food technological conditions.
CONCLUSION
Novel strain L. casei BI005 and strain 1LB of L. casei group isolated from homemade traditional fermented beverage koumiss (mare’s milk) were selected as the potential probiotics because of beneficial properties viz., tolerance to simulating gastrointestinal tract conditions, high antimicrobial activity and lack of acquired genes of antibiotic resistance which render them promising candidates as starter cultures in the production of probiotic fermented foods.
Novel Lactobacillus strain 4LB of L. brevis group isolated from homemade traditional fermented beverage koumiss (mare’s milk) in spite of beneficial properties viz., tolerance to simulating gastrointestinal tract conditions and high antimicrobial activity was excluded from the selection as the potential probiotic because of a possibility to acquire genes of streptomycin and gentamicin resistance.
Detection of resistance/intermediate resistance to tetracycline for 4 isolates (3 novel isolates inclusive) from 3 different phylogenetic groups among 16 tested Lactobacillus strains isolated from homemade traditional and industrial fermented dairy products leads to the opinion that lactobacilli with tetracycline resistance are common in fermented dairy foods consumed in Kazakhstan.
Two novel Lactobacillus strains, namely 3LB from L. brevis group and 11LB from L. casei group isolated from traditional homemade fermented dairy foods sour cream and beverage airan respectively were excluded from the selection as the potential probiotics because of a possibility to acquire genes of tetracycline, β-lactams, streptomycin and gentamicin resistances. However, they can be used as a raw material to extract antimicrobial substances which, apparently, cause high antimicrobial activity with a broad spectrum including antifungal activity against Candida albicans evaluated in the present project and may be useful in the food and pharmaceutical industries.
ACKNOWLEDGMENTS
This study was performed within the Research Project # 0113RK00503 funding by the Scientific Committee of the Ministry of Education and Science of RK. Special thanks to Prof. Seytembetov T and Prof. Nurgozhin T for support of the study.
REFERENCES
- Hill, C., F. Guarner, G. Reid, G.R. Gibson and D.J. Merenstein et al., 2014. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol., 11: 506-514.
CrossRefDirect Link - Seale, J.V. and M. Millar, 2013. Probiotics: A new frontier for infection control. J. Hosp. Infect., 84: 1-4.
CrossRefDirect Link - Quinto, E.J., P. Jimenez, I. Caro, J. Tejero, J. Mateo and T. Girbes, 2014. Probiotic lactic acid bacteria: A review. Food Nutr. Sci., 5: 1765-1775.
CrossRefDirect Link - Zago, M., M.E. Fornasari, D. Carminati, P. Bums and V. Suarez et al., 2011. Characterization and probiotic potential of Lactobacillus plantarum strains isolated from cheeses. Food Microbiol., 28: 1033-1040.
CrossRefDirect Link - Reale, A., T. di Renzo, F. Rossi, T. Zotta and L. Iacumin et al., 2015. Tolerance of Lactobacillus casei, Lactobacillus paracasei and Lactobacillus rhamnosus strains to stress factors encountered in food processing and in the gastro-intestinal tract. LWT-Food Sci. Technol., 60: 721-728.
CrossRefDirect Link - Begley, M., C.G.M. Gahan and C. Hill, 2005. The interaction between bacteria and bile. FEMS Microbiol. Rev., 29: 625-651.
CrossRefDirect Link - Rushdy, A.A. and E.Z. Gomaa, 2013. Antimicrobial compounds produced by probiotic Lactobacillus brevis isolated from dairy products. Ann. Microbiol., 63: 81-90.
CrossRefDirect Link - Belicova, A., M. Mikulasova and R. Dusinsky, 2013. Probiotic potential and safety properties of Lactobacillus plantarum from slovak bryndza cheese. BioMed Res. Int., Vol. 2013.
CrossRefDirect Link - Patel, A.R., N.P. Shah and J.B. Prajapati, 2012. Antibiotic resistance profile of lactic acid bacteria and their implications in food chain. World J. Dairy Food Sci., 7: 202-211.
Direct Link - Morandi, S., T. Silvetti, J.M. Miranda Lopez and M. Brasca, 2015. Antimicrobial activity, antibiotic resistance and the safety of lactic acid bacteria in raw milk valtellina casera cheese. J. Food Saf., 35: 193-205.
CrossRefDirect Link - Sharma, P., S.K. Tomar, P. Goswami, V. Sangwan and R. Singh, 2014. Antibiotic resistance among commercially available probiotics. Food Res. Int., 57: 176-195.
CrossRefDirect Link - Gueimonde, M., B. Sanchez, C.G. de los Reyes-Gavilan and A. Margolles, 2013. Antibiotic resistance in probiotic bacteria. Front. Microbiol., Vol. 4.
CrossRefDirect Link - Ammor, M.S., A.B. Florez and B. Mayo, 2007. Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria. Food Microbiol., 24: 559-570.
CrossRefDirect Link - Florez, A.B., V. Ladero, P. Alvarez-Martin, M.S. Ammor, M.A. Alvarez and B. Mayo, 2007. Acquired macrolide resistance in the human intestinal strain Lactobacillus rhamnosus E41 associated with a transition mutation in 23S rRNA genes. Int. J. Antimicrob. Agents, 30: 341-344.
CrossRefDirect Link - Jose, N.M., C.R. Bunt and M.A. Hussain, 2015. Implications of antibiotic resistance in probiotics. Food Rev. Int., 31: 52-62.
CrossRefDirect Link - Nawaz, M., J. Wang, A. Zhou, C. Ma and X. Wu et al., 2011. Characterization and transfer of antibiotic resistance in lactic acid bacteria from fermented food products. Curr. Microbiol., 62: 1081-1089.
CrossRefDirect Link - Mojgani, N., F. Hussaini and N. Vaseji, 2015. Characterization of indigenous Lactobacillus strains for probiotic properties. Jundishapur J. Microbiol., Vol. 8.
Direct Link - Li, K.G., G.P. Pogossian, A.K. Moldagulova, E.E. Bekenova and A. Abdikadirova et al., 2014. Identification of L. casei and L. rhamnosus in the dairy products using Real-Time PCR assay. South Asian J. Exp. Biol., 4: 222-225.
Direct Link - Vinderola, C.G. and J.A. Reinheimer, 2003. Lactic acid starter and probiotic bacteria: A comparative in vitro study of probiotic characteristics and biological barrier resistance. Food Res. Int., 36: 895-904.
CrossRefDirect Link - De Vegas, E.Z., B. Nieves, M. Araque, E. Velasco, J. Ruiz and J. Vila, 2006. Outbreak of infection with Acinetobacter strain RUH 1139 in an intensive care unit. Infect. Control Hosp. Epidemiol., 27: 397-403.
CrossRefPubMedDirect Link - Werle, E., C. Schneider, M. Renner, M. Volker and W. Fiehn, 1994. Convenient single-step, one tube purification of PCR products for direct sequencing. Nucleic Acids Res., 22: 4354-4355.
Direct Link - Haller, D., H. Colbus, M.G. Ganzle, P. Scherenbacher, C. Bode and W.P. Hammes, 2001. Metabolic and functional properties of lactic acid bacteria in the gastro-intestinal ecosystem: A comparative in vitro study between bacteria of intestinal and fermented food origin. Syst. Applied Microbiol., 24: 218-226.
CrossRefPubMedDirect Link - Cumming, G., F. Fidler and D.L. Vaux, 2007. Error bars in experimental biology. J. Cell Biol., 177: 7-11.
CrossRefDirect Link - Tagg, J.R., A.S. Dajani and L.W. Wannamaker, 1976. Bacteriocins of gram-positive bacteria. Bacteriol. Rev., 40: 722-756.
CrossRefPubMedDirect Link - Okkers, D.J., L.M.T. Dicks, M. Silvester, J.J. Joubert and H.J. Odendaal, 1999. Characterization of pentocin TV35b, a bacteriocin-like peptide isolated from Lactobacillus pentosus with a fungistatic effect on Candida albicans. J. Applied Microbiol., 87: 726-734.
CrossRefDirect Link - Charteris, W.P., P.M. Kelly, L. Morelli and J.K. Collins, 1998. Antibiotic susceptibility of potentially probiotic Lactobacillus species. J. Food Prot., 61: 1636-1643.
CrossRefDirect Link - Horackova, S., K. Zaludova and M. Plockova, 2011. Stability of selected Lactobacilli in the conditions simulating those in the gastrointestinal tract. Czech J. Food Sci., 29: S30-S35.
Direct Link - Hyacinta, M., K.S. Hana, B. Andrea and C. Barbora, 2015. Bile tolerance and its effect on antibiotic susceptibility of probiotic Lactobacillus candidates. Folia Microbiol., 60: 253-257.
CrossRefDirect Link - Felis, G.E. and F. Dellaglio, 2007. Taxonomy of lactobacilli and bifidobacteria. Curr. Issues Intest. Microbiol., 8: 44-61.
PubMedDirect Link - Shevtsov, A.B., A.R. Kushugulova, I.K. Tynybaeva, S.S. Kozhakhmetov, A.B. Abzhalelov, K.T. Momynaliev and L.G. Stoyanova, 2011. Identification of phenotypically and genotypically related Lactobacillus strains based on nucleotide sequence analysis of the groEL, rpoB, rplB and 16S rRNA genes. Microbiology, 80: 672-681.
CrossRefDirect Link - Lebeer, S., J. Vanderleyden and S.C.G. de Keersmaecker, 2008. Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev., 72: 728-764.
CrossRefDirect Link - Van de Guchte, M., P. Serror, C. Chervaux, T. Smokvina, S.D. Ehrlich and E. Maguin, 2002. Stress responses in lactic acid bacteria. Antonie Leeuwenhoek, 82: 187-216.
CrossRefPubMedDirect Link - Pfeiler, E.A., M.A. Azcarate-Peril and T.R. Klaenhammer, 2007. Characterization of a novel bile-inducible operon encoding a two-component regulatory system in Lactobacillus acidophilus. J. Bacteriol., 189: 4624-4634.
CrossRefDirect Link - Dobson, A., P.D. Cotter, R.P. Ross and C. Hill, 2012. Bacteriocin production: A probiotic trait? Appl. Environ. Microbiol., 78: 1-6.
CrossRefDirect Link - Sharma, A. and S. Srivastava, 2014. Anti-Candida activity of spent culture filtrate of Lactobacillus plantarum strain LR/14. J. Mycol. Med., 24: e25-e34.
CrossRefDirect Link - Tropcheva, R., D. Nikolova, Y. Evstatieva and S. Danova, 2014. Antifungal activity and identification of Lactobacilli, isolated from traditional dairy product katak. Anaerobe, 28: 78-84.
CrossRefDirect Link - Falguni, P., V. Shilpa and B. Mann, 2010. Production of proteinaceous antifungal substances from Lactobacillus brevis NCDC 02. Int. J. Dairy Technol., 63: 70-76.
CrossRefDirect Link - Hassan, Y.I. and L.B. Bullerman, 2008. Antifungal activity of Lactobacillus paracasei ssp. tolerans isolated from a sourdough bread culture. Int. J. Food Microbiol., 121: 112-115.
CrossRefDirect Link - Delavenne, E., R. Ismail, A. Pawtowski, J. Mounier, G. Barbier and G. Le Blay, 2013. Assessment of lactobacilli strains as yogurt bioprotective cultures. Food Control, 30: 206-213.
CrossRefDirect Link - Wang, H., Y. Yan, J. Wang, H. Zhang and W. Qi, 2012. Production and characterization of antifungal compounds produced by Lactobacillus plantarum IMAU10014. PloS One, Vol. 7.
CrossRefDirect Link - De Keersmaecker, S.C., T.L. Verhoeven, J. Desair, K. Marchal, J. Vanderleyden and I. Nagy, 2006. Strong antimicrobial activity of Lactobacillus rhamnosus GG against Salmonella typhimurium is due to accumulation of lactic acid. FEMS Microbiol. Lett., 259: 89-96.
CrossRefPubMedDirect Link - Annuk, H., J. Shchepetova, T. Kullisaar, E. Songisepp, M. Zilmer and M. Mikelsaar, 2003. Characterization of intestinal lactobacilli as putative probiotic candidates. J. Applied Microbiol., 94: 403-412.
CrossRefPubMedDirect Link - Haghshenas, B., Y. Nami, M. Haghshenas, N. Abdullah, R. Rosli, D. Radiah and A.Y. Khosroushahi, 2015. Bioactivity characterization of Lactobacillus strains isolated from dairy products. Microbiol. Open, 4: 803-813.
CrossRefDirect Link - Al-Madboly, L.A. and A.K. Abdullah, 2015. Potent antagonistic activity of Egyptian Lactobacillus plantarum against multiresistant and virulent food-associated pathogens. Front. Microbiol., Vol. 6.
CrossRefDirect Link - Botina, S.G., E.U. Poluektova, A.A. Glazova, N.V. Zakharevich and N.V. Koroban et al., 2011. Antibiotic resistance of potential probiotic bacteria of the genus Lactobacillus from human gastrointestinal microbiome. Microbiology, 80: 164-171.
CrossRefDirect Link - Rosander, A., E. Connolly and S. Roos, 2008. Removal of antibiotic resistance gene-carrying plasmids from Lactobacillus reuteri ATCC 55730 and characterization of the resulting daughter strain, L. reuteri DSM 17938. Applied Environ. Microbiol., 74: 6032-6040.
CrossRefDirect Link - Hummel, A.S., C. Hertel, W.H. Holzapfel and C.M.A.P. Franz, 2007. Antibiotic resistances of starter and probiotic strains of lactic acid bacteria. Applied Environ. Microb., 73: 730-739.
CrossRefDirect Link - Mayrhofer, S., K.J. Domig, C. Mair, U. Zitz, G. Huys and W. Kneifel, 2008. Comparison of broth microdilution, Etest and agar disk diffusion methods for antimicrobial susceptibility testing of Lactobacillus acidophilus group members. Applied Environ. Microbiol., 74: 3745-3748.
CrossRefDirect Link - Munoz, M.D.C.C., N. Benomar, L.L. Lerma, A. GAlvez and H. Abriouel, 2014. Antibiotic resistance of Lactobacillus pentosus and Leuconostoc pseudomesenteroides isolated from naturally-fermented Alorena table olives throughout fermentation process. Int. J. Food Microbiol., 172: 110-118.
CrossRefDirect Link - Klare, I., C. Konstabel, G. Werner, G. Huys and V. Vankerckhoven et al., 2007. Antimicrobial susceptibilities of Lactobacillus, Pediococcus and Lactococcus human isolates and cultures intended for probiotic or nutritional use. J. Antimicrob. Chemother., 59: 900-912.
CrossRefDirect Link - Kiwaki, M. and T. Sato, 2009. Antimicrobial susceptibility of Bifidobacterium breve strains and genetic analysis of streptomycin resistance of probiotic B. breve strain Yakult. Int. J. Food Microbiol., 134: 211-215.
CrossRefDirect Link - Sato, T. and T. Iino, 2010. Genetic analyses of the antibiotic resistance of Bifidobacterium bifidum strain Yakult YIT 4007. Int. J. Food Microbiol., 137: 254-258.
CrossRefDirect Link - Liu, C., Z.Y. Zhang, K. Dong, J.P. Yuan and X.K. Guo, 2009. Antibiotic resistance of probiotic strains of lactic acid bacteria isolated from marketed foods and drugs. Biomed. Environ. Sci., 22: 401-412.
CrossRefDirect Link - Rojo-Bezares, B., Y. Saenz, P. Poeta, M. Zarazaga, F. Ruiz-larrea and C. Torres, 2006. Assessment of antibiotic susceptibility within lactic acid bacteria strains isolated from wine. Int. J. Food Microbiol., 111: 234-240.
CrossRefDirect Link - El-Deeb, N., M.M. Sharaf and H. El-Adawi, 2015. Antibacterial and plasmid curing activity of lactic acid bacteria against multidrug resistant bacteria strains. Int. J. Pharmacol., 11: 114-121.
CrossRefDirect Link