
M. Senthil Kumar
Deptartment of Biotechnology, Bharathiar University, Coimbatore, India
K. Selvam
Deptartment of Biotechnology, Dr. NGP College of Arts and Science, Coimbatore, India
Singaravel
Central Institute of Brackishwater Aquaculture (ICAR), Chennai, India
Asian Journal of Applied Sciences
Year: 2012 | Volume: 5 | Issue: 5 | Page No.: 252-265







ABSTRACT
Streptomyces radiopugnans MS1 was isolated from marine environment and identified as a potent strain for producing L-Asparaginase from Tapioca effluent an inexpensive substrate. To optimize the L-Asparaginase production of this bacterial strain, 23 fermentation variables were screened using a Plackett-Burman design and were then further optimized via response surface methodology based on a Box-/Behnken design. Four significant variables, i.e., Tapioca Effluent, Corn steep liquor, L-Aspaparagine and Areation, were selected. The optimum values of the tested variables by response surface methodology were; Tapioca Effluent, 5 (% v/v); Corn steep liquor, 2 (% w/v); L-Aspaparagine, 0.003 (% w/v); Aeration, 0.7 (vvm) found to be optimum for L-Asparaginase production. The experimental result (19.5 IU mL-1) in a medium optimized for L-Asparaginase production was in good agreement with the predicted value of a quadratic model (19.8 IU mL-1). A verification experiment was carried out to examine model validation and revealed more than 99% validity. The model was very satisfactory as the coefficient of determination was 0.99. L-Asparaginase production in the optimized medium (19.5±0.33 IU mL-1) increased 4.81-fold over that of the basal medium (4.05±0.84 IU mL-1) in the fermenter.
PDF Abstract XML References Citation
Received: November 15, 2011;
Accepted: January 26, 2012;
Published: March 26, 2012
How to cite this article
M. Senthil Kumar, K. Selvam and Singaravel, 2012. Statistical Assessment of Medium Components by Factorial Design and Surface Methodology of L-Asparaginase Production by Isolated Streptomyces radiopugnans MS1 in Submerged Fermentation using Tapioca Effluent. Asian Journal of Applied Sciences, 5: 252-265.
DOI: 10.3923/ajaps.2012.252.265
URL: https://scialert.net/abstract/?doi=ajaps.2012.252.265
DOI: 10.3923/ajaps.2012.252.265
URL: https://scialert.net/abstract/?doi=ajaps.2012.252.265
INTRODUCTION
L-asparaginase is an important enzyme as therapeutic agent used in treatment of acute lymphocytic leukemia (mainly in children), Hodgkin disease, acute myelocytic leukemia, acute myelomonocytic leukemia, chronic lymphocytic leukemia, lymphosarcoma treatment, reticlesarcoma and melanosacroma (Stecher et al., 1999; Verma et al., 2007).
L-asparaginase is widely distributed in both prokaryotic and eukaryotic cells and has been intensively studied over the past five decades. This enzyme is existing in many animal tissues, plants and bacteria. L-asparaginase produced by a large number of microorganisms such as E. coli (Khushoo et al., 2004; Derst et al., 1994), Erwinia cartovora (Aghaiypour et al., 2001; Borisova et al., 2003), Erwinia chrysanthemi (Kotzia and Labrou, 2007), Enterobacter aerogenes (Mukherjee et al., 2000), Pseudomonas aeruginosa (El-Bessoumy et al., 2004), Candida utilis (Kil et al., 1995), Thermus thermophilus (Pritsa and Kyridio, 2001) Staphylococcus aureus (Muley et al., 1998) and Filamentous Fungi (Elzainy and Ali, 2006). Moreover, L-asparaginase was demonstrated and characterized in higher plants on account of the vital role of this enzyme in the nitrogen nutrition (Sodek et al., 1980; El-Shora et al., 2005; Cho et al., 2007). Actinomycetes have been shown to be a good source for L-Asparaginase too.
Nowadays, marine actinomycetes have been widely recognized as a potential source of new drug candidates in particular marine Streptomyces are promising source of antitumour drugs and therapeutic enzymes (Jensen et al., 2005). Marine actinomycetes have been reported to produce functionally unique metabolites and enzymes that are not found in their terrestrial counterparts. This is due to their extreme living conditions within the marine environment, Recent studies have shown that few antitoumor compounds isolated from marine actinomycetes are under clinical trials (Ye et al., 2009). Hence, as a new source of antitumour drug candidates, marine actinomycetes have attracted serious attention in the last decade.
With a view to characterize enzymes with less toxic side effects, several members of a larger family of homologous l-ASNases have been thoroughly investigated over many years (Boyse et al., 1967; Ehrman et al., 1971; Cammack et al., 1972; Wriston and Yellin, 1973; Krasotkina et al., 2004; Kotzia and Labrou, 2005). In addition, because the anti-tumour activity of L-Asparaginase is also a function of its halflife in the blood (Fernandes and Gregoriadis, 1997), attempts have been made to increase the half-life, for example by entrapment of the enzyme in liposomes (Neerunjun and Gregoriadis, 1976) or microcapsules (Chang, 1984) and by covalent coupling to macromolecules such as dextran (Wileman et al., 1986), albumin (Poznansky et al., 1982) or monomethoxypolyethylene glycol (mPEG) (Kamisaki et al., 1982) which is on the market. Unfortunately, none of these approaches have managed to eliminate the disadvantages of L-Asparaginase treatment, leaving scientists with the need to identify and characterize new enzymes with better properties.
Medium composition is one of the most important parameters when enzymes are produced for industrial purposes because around 30-40% of the production cost is estimated to be the cost of the growth medium (Kirk et al., 2002; Youssef and Al-Omair, 2008). Statistical approaches have helped to enhance product yield and reduce the cost of production, thereby making the fermentation process economical and cost effective (Kaur and Satyanarayana, 2005; Shah et al., 2010).
In the optimization of media compounds, Plackett-Burman designs are used as a screening method in order to select the variables that influence a system. However, they do not give an optimum value for each variable (Jalbani et al., 2006; Youssef and Berekaa, 2009) and further optimization is needed. Response Surface Methodology (RSM) has been widely used to evaluate and understand the interactions between different physiological and nutritional parameters (Bandaru et al., 2006; Charyulu and Gnanamani, 2010). It is an efficient mathematical approach widely applied in the optimization of the fermentation process (Panda et al., 2007) and media component, e.g., production of enzymes, biomass, spore, and other metabolites (Ghosalkar et al., 2008; Jaiswal et al., 2011). RSM which includes factorial design and regression analysis, can be used to help evaluate the effective factors and build models (De Coninck et al., 2000; Akhir et al., 2009). It can give information about the interaction between variables and can be used to select optimum conditions of variables for a desirable response and multiple responses at the same time (Lo et al., 2009). Finally, after model building and optimization, the predicted model is verified.
In this investigation, an attempt has been made to improve L-Asparaginase production from Streptomyces species by statistical approaches using a Plackett-Burman design and RSM in submerged culture.
MATERIALS AND METHODS
Sample collection and strain isolation: A total of 254 sediment samples were collected during six separate research expeditions to Bay of Bengal, Tamilnadu, India.
Samples of the top 1 cm of sediment were collected in sterile 50 mL strile plastic bags by divers using scuba gear when necessary. Sediment sample depth ranges, numbers, locations and dates were as follows: 0 to 30 m, 42 sediment samples (chennai, Nov 2009); 0 to 30 m, 65 sediment samples (Chidambaram, Dec 2010); 10 to 30 m, 42 sediment samples (Pondicherry, Jan, 2010); 0 to 30 m, 25 sediment samples (Tuticorin, Feb, 2010); 0 to 30 m, 57 sediment samples (Rameshwaram, March, 2010); 0 to 30 m, 23 sediment samples (Kanyakumari, Apr, 2010); Samples were kept at room temperature during the expedition and at 4°C upon return to the laboratory.
Enrichment and screening of slowly growing marine microorganisms: One gram each of the sieved soil samples was treated with 100 mL of tapioca effluent and incubated at an ambient temperature for about a week at 200 rpm (Mincer et al., 2002; Pisano et al., 1986). The dilution-and-heat-shock method was carried out as follows with Actinomycete isolation agar and Starch casein agar medium supplemented with phenol red (0.009% final concentration) (Himedia, Mumbai, India) (Gulati et al., 1997). Media were prepared with 100% filtered natural sea water. The media components included cycloheximide (20 mg L-1), pravastatin (10 mg L-1), trimethoprim (2 mg L-1) and nalidixic acid (10 mg L-1) to prevent other non-actinomycete bacteria and fungal growth (Kuster and Williams, 1964).
One millilitter of enriched wet sediment was added to 4 mL of sterile seawater, heated for 6 min at 55°C, vigorously shaken and further diluted (1:4) in sterile seawater and 50 μL of each dilution was suspensions were spread, in triplicate onto agar-based isolation media. Actinomycetes generally appeared after week of incubation at 25 to 28°C. Colonies with pink zones were considered as L-asparaginase producing strains.
The colonies were picked and streaked on AIA an SCA by quadrant streak method to isolate the single pure culture. The isolated colonies were sub cultured on AIA and SCA media in refrigerated conditions for further work to carried.
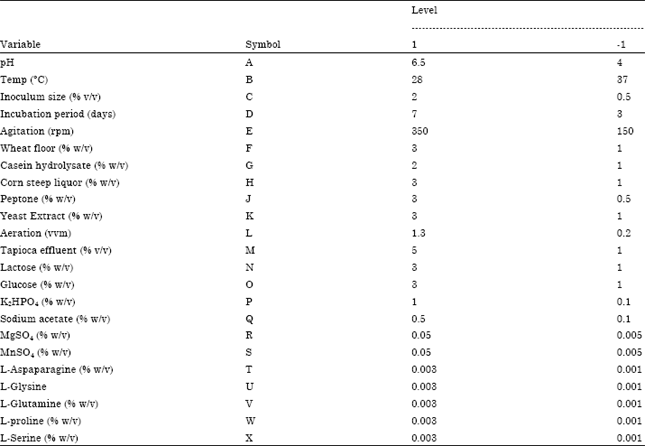
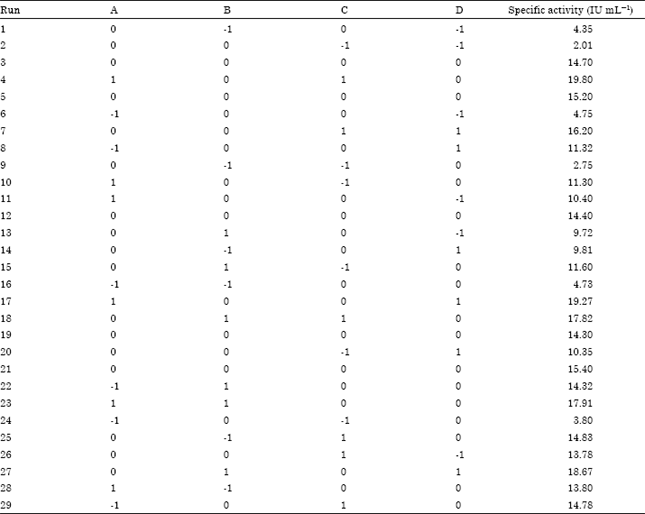
Selection of the significant media components for process modeling: The purpose of Plackett-Burman factorial design was to identify significant medium components affecting the L-Asparaginase production. This factorial design is important when large numbers of factors are to be considered for optimization. Thirty two experiments were obtained for 23 factors namely Glucose ,Fructose, Galactose, Lactose ,Tapioca Effluent, Wheat floor, Tryptone ,Yeast Extract, Corn steep liquor, peptone, Casein Hydrolysate, L-Aspaparagine, L-Glysine, L-Cystein, L-Serine, L-Glutamine, L-proline, K2HPO4, MgSO4, MnSO4, sodium acetate, pH and Temperature. Each variable was represented at two levels, upper (“high (1)”) and lower (“low (-1)”) levels of the range covered by each variable and the response (Abdel-Fettah et al., 2002) Table 1 shows a 24-run Plackett-Burman experimental design (Plackett and Burman, 1946).
This model does not describe the interaction among factors and it is used to screen and evaluate important factors that influence the response. From the regression analysis of the variables, the factors having significant effect on L-Asparaginase production were further optimized by Response Surface Methodology (RSM).
| Table 1: | Assigned concentrations of variables at different levels in Plackett–Burman design for production |
 | |
Optimization using response surface methodology: Response surface methodology was adopted for improving L-Asparaginase find the interactive effects of the four variables found to be significant from the Plackett-Burman experiments. Box-/Behnken design of response surface method was used to obtain data that fits a full second order polynomial model. Table 5 represents the design matrix of 29 trials experiment. Regression analysis was performed on the data obtained. A second-order polynomial equation was used to fit the data by multiple regression procedure. Using this design, factors of highest confidence level percentage are prescribed into three levels, coded -/1, 0 and /1 for low, middle and high concentration (or value). For predicting the optimal point, a second order polynomial function was fitted to correlate relationship between variables and response (asparaginase activity). The three dimensional graphical representation of model equation represents the individual and interactive effect of the test variables on the response.
Statistical analysis of the data: The data on enzyme activity was subjected to multiple linear regression using Design-Expert® software version 8.0.6.(Stat-Ease,Inc., 2021 East Hennepin Ave., Suite 480, Minneapolis, MN 55413) to estimate t-value, p-value and confidence level. The significance level (p-value) was determined using the Students t-test. The t-test for any individual effect allows an evaluation of the probability of finding the observed effect purely by chance. If this probability is sufficiently small, the idea that the effect was caused by varying the level of the variable under test is accepted. Confidence level is an expression of the p-value in percent.
Scale-up studies in a fermenter: After optimization studies in shaken flasks, protease was produced in a 3.5-liter Laboratory Fermenter L1523, (Bioengineering AG ,Sagenrainstrasse 7 CH-8636 Wald, Switzerland). Two percent of the 24 h seed culture was inoculated into the optimal medium and fermented at pH 6.5 and 28°C with 350 rpm agitation. The dissolved oxygen concentration was maintained at 80% air saturation throughout the fermentation process.
RESULTS AND DISCUSSION
Isolation and identification of the isolated strain: From 254 marine sediment samples, 29 Actinomycetes were obtained. The number of actinomyycetes isolated from East coastal region of India (Chennai-3 colonies, Chidambaram-6 colonies, Pondicherry-2 colonies, Tuticorin-5 colonies, Rameshwaram-8 colonies, Kanyakumari-5 colonies). According by morphology and chemotaxonomy about 89% of the isolates were presumed to be in genus streptomyces and 11% non streptomyces. The streptomyces strains were screened L-Asparaginase activity on phenol red incorporated medium and 6 (3, 6, 10, 14, 23 and 28) strains were found to produces significant range of L-Asparaginase. Farther than six strains, strain 2 (Genebank accession No. HQ623051) was expressed premier activity.
It was identified as Streptomyces radiopugnans MS1 based on partial 16 sRNA gene sequence homology and a standard method for bacterial identification. These isolate was maintained in AIA and SCA media at -20°C.
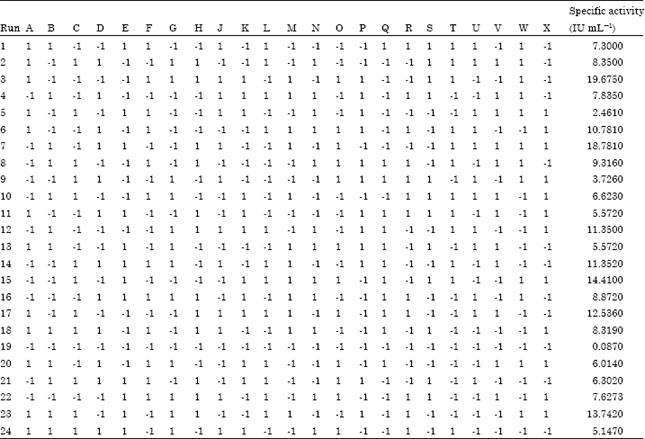
Plackett-burman designs: The first optimization step identified the significant factors for L-Asparaginase production from Streptomyces radiopugnans MS1 using a 24-run Plackett-Burman design (Table 1, 2).
The statistical significance of the ratio, between the of mean square variation, due to regression and the mean square residual error, was tested using Analysis of Variance (ANOVA). ANOVA is a statistical technique that subdivides the total variation of a set of data into component associated to specific sources of variation for the purpose of testing hypotheses for the modeled parameters. According to the ANOVA , the F values for all regressions were high, what indicates that most of the variations on the response variable can be explained by the regression equation.
Table 3 represents the effect of each variable along with the mean squares, F-values and p-values. The observed L-Asparaginase production varied from 0.087 to 19.675 IU mL-1, reflecting the importance of medium optimization to attain higher yields. Values of "Prob>F" less than 0.0500 indicate model terms are significant. In general, larger magnitudes of t and smaller of p, indicates that the corresponding coefficient term (Myers and Montgomery, 2002). The Model F-value of 844.94 implies the model is significant. There is only a 0.01% chance that a "Model F-Value" this large could occur due to noise. The adequate precision which measures the signal to noise ratio was 136.121 for response which indicates an adequate signal. A ratio of >4 is desirable. This model can be used to navigate the design space. The ‘Pred R-Squared’ of 0.9939 is in reasonable agreement with ‘Adjusted R-Squared’ of 0.9983.
The model of equation can be shows as:
 |
| Table 2: | Plackett-Burman experimental design for evaluation of factors affecting L-asparaginase activity |
 | |
| Table 3: | ANOVA for plackett-burman experimental design analysis of variance table [Partial sum of squares-Type III] |
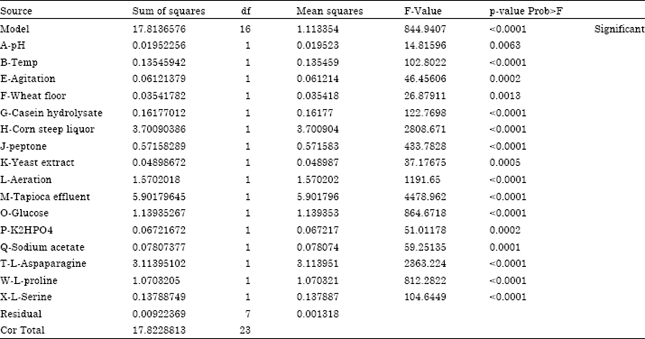 | |
| Std. Dev. 0.036; R-Squared 0.9995; Mean 2.84; Adj R-Squared 0.9983; C.V. % 1.28; Pred R-Squared 0.9939; PRESS 0.11 Adeq Precision 136.121 | |
The Pareto chart of standardization histogram graph (Fig. 1) showed that only Tapioca Effluent, Corn steep liquor, L-Aspaparagine, Areation, Glucose, L-Proline, peptone, Casein Hydrolysate, L-Serine, Temp, sodium acetate, KH2SO4, Agitation, Yeast Extract Wheat floor and pH (significance level (Threshold of t value = 2.364), crosses the critical value and was considered to significantly influence L-Asparaginase production by Streptomyces radiopugnans MS1. MgSO4, MnSO4, Lactose, L-Glysine, Incubation period, L-Glutamine and inculum size (Fig. 1-3).
Optimization by response surface methodology: Box-Behnken is the most accepted statistical technique for bioprocess optimization to examine the L-asparaginase activity.
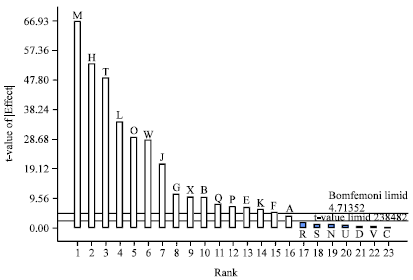 | |
| Fig. 1: | Pareto chart of standardadized effects of twenty three factor screening design for the production of Lasparaginase. Values of "Prob > F" less than 0.0500 indicate model terms are significant. In this study M, H,T, L,O, W, J,G, X, B,Q, P, E, K, F and A are significant model terms and R, S, N, U, D, V and C are Non significant (Negative effect) |
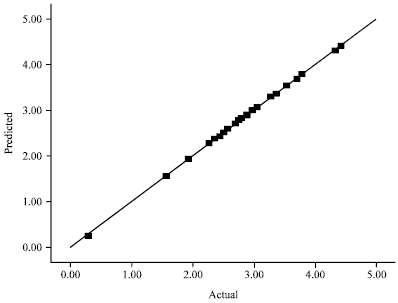 | |
| Fig. 2: | Parity plot showing the distribution of Actual vs. predicted values of L-Asparaginase activity |
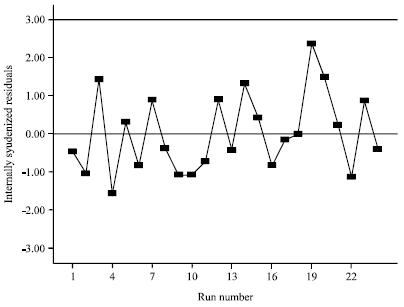 | |
| Fig. 3: | Residual distribution plot for L-asparaginase activity |
| Table 4: | Experimental design obtained by applying the Box-/Behnken design methodology for four factors |
 | |
The design matrix and the corresponding results of the experiments to determine the effect of Tapioca Effluent, Corn steep liquor, L-Aspaparagine and Areation are depicted in Table 4 and 5.
 |
At the model level, The Model F-value of 742.17 implies the model is significant (Table 6). There is only a 0.01% chance that a "Model F-Value" this large could occur due to noise. Values of "Prob > F" less than 0.0500 indicate model terms are significant.In this case A, B, C, D, AB, AC, BC, CD, A2, B2, C2, D2 are significant model terms. Values greater than 0.1000 indicate the model terms are not significant. The "Lack of Fit F-value" of 0.29 implies the Lack of Fit is not significant relative to the pure error. There is a 94.66% chance that a "Lack of Fit F-value" this large could occur due to noise. Non-significant lack of fit is good. The "Pred R-Squared" of 0.9955 is in reasonable agreement with the "Adj R-Squared" of 0.9973."Adeq Precision" measures the signal to noise ratio. A ratio greater than 4 is desirable. Your ratio of 96.042 indicates an adequate signal. This model can be used to navigate the design space.
| Table 5: | Box-/Behnken factorial experimental design |
 | |
Presenting experimental results in the form of surface plots (Fig. 4a-d) showed that near to moderate levels of Tapioca Effluent, Corn steep liquor, L-Aspaparagine and Areation supported high asparaginase activity.
From statistical analysis, it can be concluded that among the test variables, Tapioca Effluent had the most significant effect on L-asparaginase activity and specific activity.
The optimal levels of the four components as obtained from the maximum point of the polynomial model were estimated using the Solver function of Design-Expert® software version 8.0.6 tools and found to be: Tapioca Effluent, 5 (% v/v); Corn steep liquor, 2 (% w/v); L-Aspaparagine,0.003 (% w/v); Aeration, 0.7 (vvm) with a predicted activity of 19.87.
Validation of the model: Optimal conditions realized from the optimization experiment were verified experimentally and compared with the calculated data from the model. The estimated L-asparaginase activity was 19.5 IU mL-1, where the predicted value from the polynomial model was 19.8 IU mL-1. This verification revealed a high degree of accuracy of the model of more than 99%, which is an evidence for the model validation under the investigated conditions.
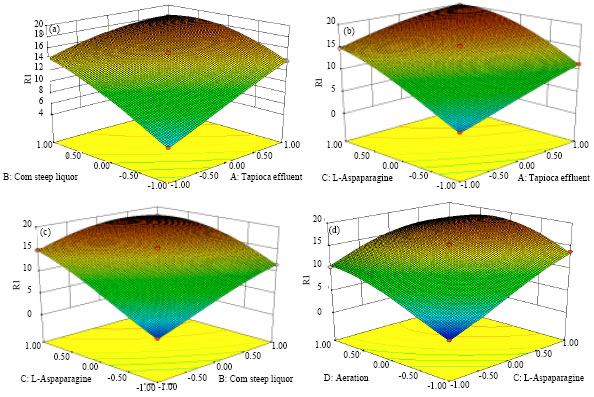 | |
| Fig. 4(a-d): | Effect of tapioca effluent, corn steep liquor, L-aspaparagine and areation on the production of L-asparaginase enzyme |
| Table 6: | ANOVA for response surface quadratic model. Analysis of variance table [Partial sum of squares-Type III] |
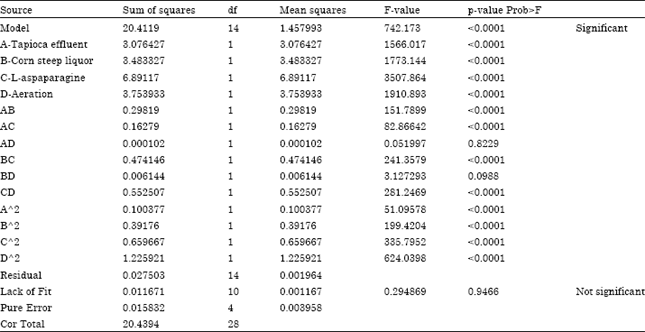 | |
| Std. Dev. 0.044; R-Squared 0.9987; Mean 3.38; Adj R-Squared 0.9973; C.V. % 1.31; Pred R-Squared 0.9955 ;PRESS 0.092; Adeq Precision 96.042. Values of "Prob > F" less than 0.0500 indicate model terms are significant. In this study A, B, C, D, AB, AC, BC, CD, A2, B2, C2, D2 are significant model terms. The "Lack of Fit F-value" of 0.29 implies the Lack of Fit is not significant relative to the pure error. There is a 94.66% chance that a "Lack of Fit F-value" this large could occur due to noise. Non-significant lack of fit is good | |
In this study, the isolated strain MS1 gave 4.05±0.84 IU mL-1 in simple basal medium that was enhanced to 19.5±0.33 IU mL-1 by use of statistical optimization tool. Thus, it can be said that the isolate MS1 is an excellent L-asparaginase producer.
In the present study a Actinomycetes strain was isolated from Bay of bengal. The strain was identified as Streptomyces radiopugnans MSl by biochemical and 16S rRNA study. Sequential statistical strategies, Plackett-Burman design followed by:
Box-Behnken were used successfully to find the optimum values of the significant factors to achieve maximum L-Asparaginase production. The predicted yield was 19.8 IU mL-1. On experimentation, 19.5 IU mL-1 L-Asparaginase yield was obtained. The experimental values were found to be very close to the predicted values and hence, the model was successfully validated. The L-Asparaginase production showed about 5 fold increases over the basal medium. The isolated strain can be used for the production of L-Asparaginase enzyme that could be of industrial value.
REFERENCES
- Abdel-Fattah, Y.R., N. Soliman, A. Gaballa, S. Sabry and A. El-Diwany, 2002. Lipase production from a novel thermophilic Bacillus sp.: Application of Plackett-Burman design for evaluating enzyme formation. Acta Microbiol. Polonica, 51: 353-366.
Direct Link - Aghaiypour, K., A. Wlodawer and J. Lubkowski, 2001. Structural basis for the activity and substrate specificity of Erwinia chrysanthemi L-asparaginase. Biochemistry, 40: 5655-5664.
CrossRefDirect Link - Akhir, S.M., S. Abd-Aziz, M.M. Salleh, R.A. Rahman, R.M. Illias and M.A. Hassan, 2009. Medium optimisation of chitinase enzyme production from shrimp waste using Bacillus licheniformis TH-1 by response surface methods. Biotechnology, 8: 120-125.
CrossRefDirect Link - Bandaru, V.V.R., S.R. Somalanka, D.R. Mendu, N.R. Madicherla and A. Chityala, 2006. Optimization of fermentation conditions for the production of ethanol from sago starch by co-immobilized amyloglucosidase and cells of Zymomonas mobilis using response surface methodology. Enzyme Microb. Technol., 38: 209-214.
CrossRefDirect Link - Panda, B.P., M. Ali and S. Javed, 2007. Fermentation process optimization. Res. J. Microbiol., 2: 201-208.
CrossRefDirect Link - Borisova, A.A., M.A. Eldarov, A.A. Zgoon, S.S. Alexandrova and N.M. Omelyanuk et al., 2003. Purification and some properties of recombinant Erwinia carotovora l-asparaginase, expressed in E.coli cells. Biomedit-Sinskaya Khimiya, 49: 502-507.
PubMedDirect Link - Boyse, E.A., L.J. Old, H.A. Campbell and L.T. Mashburn, 1967. Suppression of murine leukemias by L-asparaginase. Incidence of sensitivity among leukemias of various types: Comparative inhibitory activities of guinea pig serum l-asparaginase and Escherichia coli L-asparaginase. J. Exp. Med., 125: 17-31.
PubMed - Cammack, K.A., D.I. Marlborough and D.S. Miller, 1972. Physical properties and subunit structure of L-asparaginase isolated from Erwinia carotovora. Biochem. J., 126: 361-379.
Direct Link - Chang, T.M., 1984. Artificial cells in medicine and biotechnology. Appl. Biochem. Biotechnol., 10: 5-24.
PubMed - Cho, C.W., H.J. Lee, E. Chung, K.M. Kim and J. Heo et al., 2007. Molecular characterization of the Soybean L-asparaginase gene induced by low temperature stress. J. Mol. Cells, 23: 280-286.
PubMedDirect Link - De Coninck, J., S. Bouquele, V. Dumortier, F. Duyme and I. Verdier-Denantes, 2000. Industrial media and fermentation processes for improved growth and protease production by Tetrahymena thermophila BIII. J. Ind. Microbiol. Biotechnol., 24: 285-290.
CrossRef - Derst, C., A. Wehner, V. Specht and K.H. Rohm, 1994. States and functions of tyrosine residues in Escherichia coli asparaginase II. Eur. J. Biochem., 224: 533-540.
Direct Link - Ehrman, M., H. Cedar and J.H. Schwartz, 1971. l-Asparaginase II of Escherichia coli. Studies on the enzymatic mechanism of action. J. Biol. Chem., 246: 88-94.
PubMed - El-Bessoumy, A.A., M. Sarhan and J. Mansour, 2004. Production, isolation and purification of L-asparaginase from Pseudomonas aeruginosa 50071 using solid-state fermentation. J. Biochem. Mol. Biol., 37: 387-393.
PubMed - Fernandes, A.I. and G. Gregoriadis, 1997. Polysialylated asparaginase: preparation, activity and pharmacokinetics. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol., 1341: 26-34.
CrossRefPubMedDirect Link - Ghosalkar, A., V. Sahai and A. Srivastava, 2008. Optimization of chemically defined medium for recombinant Pichia pastoris for biomass production. J. Bioresour. Technol., 99: 7906-7910.
CrossRef - Gulati, R., R.K. Saxena and R. Gupta, 1997. A rapid screening for L-asparaginase producing micro-organisms. Lett. Applied Microbiolo., 24: 23-26.
PubMed - Kamisaki, Y., H. Wada, T. Yagura, H. Nishimura, A. Matsushima and Y. Inada, 1982. Increased antitumor activity of Escherichia coli. L-asparaginase by modification with monomethoxypolyethylene glycol. Gann, 73: 470-474.
PubMed - Kaur, P. and T. Satyanarayana, 2005. Production of cellbound phytase by Pichia anomala in an economical cane molasses medium: Optimization using statistical tools. Process Biochem., 40: 3095-3102.
CrossRef - Khushoo, A., Y. Pal, B.N. Singh and K.J. Mukherjee, 2004. Extracellular expression and single step purification of recombinant Escherichia coli L-asparaginase II. Prot. Exp. Purif., 38: 29-36.
CrossRefDirect Link - Kil, J.O., G.N. Kim and I. Park, 1995. Extraction of extracellular l-asparaginase from Candida utilis. Biosci. Biotechnol. Biochem., 59: 749-750.
PubMed - Kirk, O., T.V. Borchert and C.C. Fuglsang, 2002. Industrial enzyme applications. Curr. Opin. Biotechnol., 13: 345-351.
CrossRefPubMedDirect Link - Kotzia, G.A. and N.E. Labrou, 2007. L-Asparaginase from Erwinia chrysanthemi 3937: Cloning, expression and characterization. J. Biotechnol., 127: 657-669.
CrossRef - Kotzia, G.A. and N.E. Labrou, 2005. Cloning, expression and characterization of Erwinia Carotovora L-asparaginase. J. Biotechnol., 119: 309-323.
Direct Link - Krasotkina, J., A.A. Borisova, Y.V. Gervaziev and N.N. Sokolov, 2004. One-step purification and kinetic properties of the recombinant l-asparaginase from Erwinia carotovora. Biotechnol. Appl. Biochem., 39: 215-221.
CrossRefPubMedDirect Link - Kuster, E. and S.T. Williams, 1964. Selection of media for isolation of Streptomycetes. Nature, 202: 928-929.
CrossRefDirect Link - Ye, L., Q. Zhou, C. Liu, X. Luo, G. Na and T. Xi, 2009. Identification and fermentation optimization of a marine-derived Streptomyces griseorubens with antitumour activity. Indian J. Mar. Sci., 38: 14-21.
Direct Link - Lo, P.K., C.Y. Tan, O. Hassan, A. Ahmad, N.M. Mahadi and R.M. Illias, 2009. Improvement of excretory overexpression for Bacillus sp. G1 cyclodextrin glucanotransferase (CGTase) in recombinant Escherichia coli through medium optimization. Biotechnology, 8: 184-193.
CrossRefDirect Link - Charyulu, E.M. and A. Gnanamani, 2010. Condition stabilization for Pseudomonas aeruginosa MTCC 5210 to yield high titers of extra cellular antimicrobial secondary metabolite using response surface methodology. Curr. Res. Bacteriol., 3: 197-213.
CrossRef - Youssef, M.M. and M.A. Al-Omair, 2008. Cloning, purification, characterization and immobilization of L-asparaginase II from E. coli W3110. Asian J. Biochem., 3: 337-350.
CrossRefDirect Link - Muley, R.G., S. Sarker, S. Ambedkar and S.R. Nail, 1998. Influence of alkali treated corn steep liquor containing medium on protein production by Staphylococcus aureus. Folia Microbiol., 43: 31-34.
CrossRef - Neerunjun, E.D. and G. Gregoriadis, 1976. Tumour regression with liposome-entrapped asparaginase: Some immunological advantages. Biochem. Soc. Trans., 4: 133-134.
PubMed - Jaiswal, N., O. Prakash, M. Talat, S.H. Hasan and R.K. Pandey, 2011. Application of response surface methodology for the determination of optimum reaction conditions (Temperature and pH) for starch hydrolysis by α-amylase. Asian J. Biochem., 6: 357-365.
CrossRef - Plackett, R.L. and J.P. Burman, 1946. The design of optimum multifactorial experiments. Biometrika, 33: 305-325.
CrossRefDirect Link - Poznansky, M.J., M. Shandling, M.A. Salkie, J.F. Elliott and E. Lau, 1982. Advantages in the use of L-asparaginase-albumin polymer as an antitumor agent. Cancer Res., 42: 1020-1025.
PubMed - Shah, A.J., R.V. Karadi and P.P. Parekh, 2010. Isolation, optimization and production of L-asparaginase from coliform bacteria. Asian J. Biotechnol., 2: 169-177.
CrossRefDirect Link - Sodek, L., P.J. Lea and B.J. Miflin, 1980. Distribution and properties of a potassium-dependent asparaginase isolated from developing seeds of Pisum sativum and other plants. Plant Physiol., 65: 22-26.
Direct Link - Stecher, A.L., P.M. de Deus, I. Polikarpov and J. Abrahao-Neto, 1999. Stability of L-asparaginase: An enzyme used in leukemia treatment. Pharmaceutica Acta Helvetiae, 74: 1-9.
CrossRefDirect Link - Elzainy, T.A. and T.H. Ali, 2006. Detection of the antitumor glutaminase-asparaginase in the filamentous fungi. J. Applied Sci., 6: 1389-1395.
CrossRefDirect Link - Mincer, T.J., P.R. Jensen, C.A. Kauffman and W. Fenical, 2002. Widespread and persistent populations of a major new marine actinomycete taxon in ocean sediments. Applied Environ. Microbiol., 68: 5005-5011.
CrossRefPubMedDirect Link - Verma, N., K. Kumar, G. Kaur and S. Anand, 2007. L-asparaginase: A promising chemotherapeutic agent. Crit. Rev. Biotechnol., 27: 45-62.
CrossRefDirect Link - Wileman, T.E., R.L. Foster and P.N.C. Elliott, 1986. Soluble asparaginase-dextran conjugates show increased circulatory persistence and lowered antigen reactivity. J. Pharm. Pharmacol., 38: 264-271.
CrossRefDirect Link - Wriston, J.C. and T.O. Yellin, 1973. L-Asparaginase: A review. Adv. Enzymol. Relat. Areas Mol. Biol., 39: 185-248.
PubMed - Pisano, M.A., M.J. Sommer and M.M. Lopez, 1986. Application of pretreatment methods for the isolation of bioactive actinomycetes from marine sediments. Applied Microbiol. Biotechnol., 25: 285-288.
CrossRef - Youssef, G.A. and M.M. Berekaa, 2009. Improved production of endoglucanase enzyme by Aspergillus terreus: Application of plackett burman design for optimization of process parameters. Biotechnology, 8: 212-219.
CrossRefDirect Link