
A. Nithya
School of Bio Sciences and Technology, Vellore Institute of Technology University, Vellore, 632014, India
S. Babu
School of Bio Sciences and Technology, VIT University, Vellore 632014, India
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 2 | Page No.: 65-71







ABSTRACT
In this study, we optimized microbial killing methods specific to tomato, carrot and cucumber pulp were optimized by applying different treatments, moist heat, dry heat and UV-C irradiation. The ability of Listeria monocytogenes, Bacillus cereus, Escherichia coli, Shigella flexneri, Aeromonas hydrophila, Staphylococcus aureus and Salmonella enterica to grow on tomato, carrot, cucumber and onion pulp was investigated. Population of A. hydrophila and S. enterica were high in tomato, carrot and cucumber. The population of A. hydrophila was maximum at 36 h in tomato and carrot while it was high in cucumber at initial 24 h. The population of S. enterica was maximum at 24 h in tomato, 36 and 48 h in carrot and 48 h in cucumber. Onion pulp supported less growth of bacteria. The observed difference in the ability of different bacteria to grow in different vegetable pulp warrants development of vegetable specific sanitizing methods to avoid salad-borne enteropathogenic infection.
PDF Abstract XML References Citation
Received: December 27, 2014;
Accepted: February 21, 2015;
Published: June 11, 2015
How to cite this article
A. Nithya and S. Babu, 2015. Population Dynamics of Enteropathogenic Bacteria in Pulp of Salad Vegetables. Plant Pathology Journal, 14: 65-71.
DOI: 10.3923/ppj.2015.65.71
URL: https://scialert.net/abstract/?doi=ppj.2015.65.71
DOI: 10.3923/ppj.2015.65.71
URL: https://scialert.net/abstract/?doi=ppj.2015.65.71
INTRODUCTION
Tomatoes, cucumbers, carrots and onions are the most common salad vegetables which make up the vegetable salad recipe in India. Contamination of these fresh produce can occur anywhere from farm to table (Ohlsson and Bengtsson, 2002), since they are handled unhygienic and sold as prewashed or in ready-to-eat form and not subject to further microbial killing steps. The risk of pathogen contamination and growth is one of the main safety concerns associated with fresh-cut produce, as highlighted by the increasing number of produce-linked foodborne outbreaks in recent years (Rodoni et al., 2012; WHO., 2011).
While much is known about the ecology of human pathogenic bacteria in foods of animal origin, the growth and interaction with fruits and vegetables is ill-defined (Beuchat, 2002). Differences in surface morphology, internal tissue composition and metabolic activities of plant tissues provide a wide range of diverse ecological niches selective for specific species or groups of microorganisms (Beuchat, 2002). Noel et al. (2010) identified set of metabolic and regulatory genes which are differentially regulated in planta in response to Salmonella.
Recently, the reports on occurrence and interaction of human pathogenic bacteria in fruits and vegetables have been reviewed (Nithya et al., 2014). Numerous studies have revealed that human pathogens can also penetrate internal plant tissues via the root (Gorbatsevich et al., 2013) and translocate and survive inside the plants. Internalized bacteria are protected from removal by washing and disinfection and therefore become a primary concern when the vegetables are eaten raw. In addition to internalization from the field, agricultural produce get contaminated during harvest, transit, storage and during marketing. While consumers are very concerned about "Food safety" in general terms, they are much less concerned about microbiological hazards (Brewer et al., 1994). In addition, they generally have inadequate knowledge about measures to prevent foodborne illnesses in the home; only 54% of consumers during a survey said they would wash a cutting board with soap and water after chopping fresh meat and before cutting fresh vegetables for a salad (Williamson et al., 1992).
Understanding the ability of human pathogenic bacteria to grow in the pulp of vegetables and fruits used as salads is the key to combat them from causing infections through food of vegetarian origin. Most of the population dynamics studies are oriented towards growth of E. coli and Salmonella in fruit pulps, fruit juices and in tomatoes, among the vegetables. The aim of the present study is to determine the population dynamics of selected food borne bacteria in tomato, carrot, cucumber and onion pulp under artificial inoculation conditions. This will help to develop technological means for use in food industries, restaurants and household to decontaminate fruits and vegetables before they are consumed raw.
MATERIALS AND METHODS
Bacterial cultures and media: The lyophilized cultures of Listeria monocytogenes MTCC-657, Bacillus cereus MTCC-6840, Escherichia coli MTCC-1588, Aeromonas hydrophila MTCC-646, Shigella flexneri MTCC-1457, Staphylococcus aureus MTCC-7405, Salmonella enterica MTCC-1167 were obtained from culture collection of the Microbial Type Culture Collection and Gene bank (MTCC), Institute of Microbial Technology, Chandigarh, India. Each culture were retrieved by adding 0.3-0.4 mL of sterile Luria-Bertani broth and mixed with the lyophilized powder to make the culture suspension and each culture were inoculated by quadrant streak on recommended media (L. monocytogenes -brain-heart infusion agar; B. cereus, A. hydrophila, S. flexneri, S. aureus, S. enterica-nutrient agar; E. coli-Luria-Bertani agar with streptomycin) and incubated at 37°C for 24 h. Single isolated pure colony was selected and maintained on nutrient broth media with 20% glycerol at -80°C. Working cultures were maintained on nutrient agar slants.
Determination of generation time of bacteria in broth culture: Generation time of each bacterium was calculated to find the growth rate of each pathogenic bacterium in broth culture and to compare with growth rate of each pathogen in vegetable pulps. A total of seven pathogenic bacteria cultures were maintained at 4°C on nutrient agar slants and sub-cultured periodically. Starter cultures of each pathogen were prepared by inoculating a loop full of cultures from the nutrient agar slants in to 250 mL side arm flask containing 100 mL of nutrient broth. Initial sampling was done immediately after inoculation (time 0 h) to determine the initial optical density at 600 nm by calorimetric method. The cultures were incubated at 37°C in shaker at 120 rpm and sampling was done at the interval of every half-an-hour up to 8 h and their optical density was determined. The generation time of each bacterium was calculated by using the equation:
g = t/n
where, g is the generation time, t is the time interval (hours or minutes), n is the number of generations (number of times the cell population doubles during the time interval). Where n is calculated by using the equation:
n = 3.3 log b/B
where, b is the as equation of bacteria at the end of the time interval and B is the No. of bacteria at the beginning of the time interval.
Preparation of inocula for pulp inoculations: Each of seven pathogenic cultures from the nutrient agar slants were transferred to 15 mL sterile test tube containing 3 mL of sterile nutrient broth and incubated at 37°C in shaker at 120 rpm for 24 h, then the cultures were transferred aseptically from the nutrient broth to 5 mL of 0.85% NaCl saline solution to adjust the suspension to the concentration of 2×108 cells mL-1 according to MacFarland turbidity scale and using spectrophotometric analysis. Each bacterial suspension from the saline was serially diluted in sterile 0.1% peptone water up to 104 dilution and finally 0.1 mL from the 104th dilution was spread plated on nutrient agar plate and incubated at 37°C for 24 h to determine the initial concentration of viable cells and about 1 mL from the same dilution was used to inoculate the vegetable pulps.
Preparation of salad vegetable pulp: Fresh, damage free carrots, cucumbers, onions and tomatoes were purchased from the local market in the city of Vellore, state of Tamil Nadu, India. The external surface of each vegetable samples was washed with sterile distilled water to remove the external contaminants and air dried in a laminar air flow hood and the surface of each vegetable was rinsed with 70% ethanol and air-dries. The outer layer of each vegetable was peeled with sterile knives and about 50 g portion from internal pulp tissues was weighed aseptically and mixed with 50 mL of sterile 0.1% peptone water and blended with surface sterilized (70% ethanol) commercial blender to form a homogenized mixture, transferred to sterile 250 mL Erlenmeyer flask and used for further study.
Optimization of conditions for the sterility of vegetable pulp: Before the inoculation study, each vegetable pulp was checked for its sterility by taking 0.1 mL aliquot from each vegetable pulp and plated on nutrient agar plate by standard spread plate technique, followed by incubating at 37°C for 24 h to find the presence of any internal flora and contaminants. Since raw vegetable pulp samples showed variable contamination load, they were subjected to different treatment prior to their inoculation with pathogenic bacteria. Two different moist heat microbial killing methods were validated viz., Pasteurization (85°C for 20 min) and boiling (100°C for 1 min), dry heat microbial killing method viz., ultra Pasteurization (149°C for 2 sec) followed by sudden cooling and UV-C irradiation method to sanitize tomato, cucumber, carrot and onion, in which the timing and temperature were optimized by us by inserting a thermometer in each vegetable pulp and the time required to reach appropriate temperature was monitored. Moist heat treatments were done in water bath, whereas dry heat treatment was done in hot-air-oven and UV-C irradiation was applied in a UV-C cabinet keeping the samples at a distance of 18 cm for 30 min to achieve 10 kJ m‾2. After treatment, about 0.1 mL of each vegetable pulp was plated on nutrient agar plate by using standard spread plate technique and incubated at 37°C for 24 h to ensure the absence of background microflora. Experiment was repeated thrice.
Determination of growth of pathogenic bacteria in vegetable pulp: Homogenized triplicate test portions of vegetable pulps (50 g) carrot, cucumber, onion and tomato in duplicates was inoculated with 1 mL (104 CFU) suspension of L. monocytogenes, B. cereus, E. coli, A. hydrophila, S. flexneri, S. aureus and S. enterica, separately and mixed with the sterile glass-rod aseptically to form the homogenate mixture of pulp and the bacteria and incubated for 0, 24, 36 and 48 h at room temperature and an un-inoculated pulp for each vegetables were kept in separate flask as a control to check the sterility of pulp before and after the incubation. At each sampling time from 0-48 h, about 1 mL pulp from each vegetable were taken aseptically and serially diluted with 0.1% peptone water up to 106th dilution and plated on nutrient agar plates (3 replications) by standard spread plate technique and incubated at 37°C for 24 h. The colonies formed after incubation were counted by using colony counter and expressed in CFC g-1. The experiment was repeated twice and an average colony count was determined.
Statistical analysis: The data of bacterial population in inoculated salad vegetable pulp were analyzed statistically using GraphPad InStat® software, version 3.06. Error bars in figures indicate standard error deviations of mean value.
RESULTS AND DISCUSSION
In the present study, observations made in generation times of seven pathogenic bacteria in liquid media are presented in Table 1 and the corresponding growth curves are presented in Fig. 1. Generation time for Shigella and Staphylococcus was found to be 145 and 136 min, respectively. Other bacteria tested had 41 (E. coli) or less than 40 min as generation time. This observation helped us in deciding the time for colony counting in vegetable pulp inoculated with these bacteria.
In this study, two different moist heat microbial killing methods were validated viz., Pasteurization (85°C for 20 min), boiling (100°C for 1 min), dry heat microbial killing method viz., ultra Pasteurization (149°C for 2 sec) and UV-C irradiation method to sanitize tomato, cucumber, carrot and onion, in which the timing and temperature were optimized by us. Bacterial colonies were observed in agar plates after different treatments and their colony count is presented in Table 2. Moist heat treatment at 100°C for 1 min was able to completely kill bacteria in tomato pulp and cucumber pulp, where as no visible effect was seen in tomato and cucumber at 85°C for 20 min and the number of bacterial colonies were comparable to untreated control. UV-C irradiation for 30 min demonstrated little effect with countable number of colonies per gram of tissues of tomato, carrot and cucumber pulp. Heat treatment at 85°C for 20 min was found to have the best effect (no bacterial growth) in carrot, respectively. It is interesting that a comparatively low temperature of 85°C but an extended time of 20 min was able to kill all bacteria in carrot. Raised temperature of 100°C was effective, to kill all bacteria in tomato and cucumber. At the same time, 149°C had no effect implicating the importance of time of heat treatment in sanitizing carrot and cucumber. Tomato and cucumber required 100°C for 1 min where as carrot required 85°C for 20 min to eliminate background microbial flora in them and other heat treatments had no effect. The onions used in the study had no cultivable endogenous bacteria as observed by the absence of colonies even in the untreated pulp.
Trinetta et al. (2011) have used non-thermal inactivation techniques like chlorine dioxide gas, ozone gas and e-beam irradiation on tomato, cantaloupe and lettuce. These methods are not cost effective and it is also reported that gaseous chemicals have disadvantages like toxicity (Lopez-Galvez et al., 2012). UV-C irradiation was found to be effective and rapid method for disinfection and extended shelf life in fresh-cut bell pepper (Rahman et al., 2011). However, UV treatment has to be used correctly in different produce and different varieties (Erickson, 2010). Alternative to the aforesaid methods, heat treatment in water bath is considered to be the simple and effective method in controlling pathogens by enzyme inactivation. Raising the temperature to 85°C for 20 min or 100°C for 1 min will quickly kill microbes and retains product’s natural viscosity. Ultra pasteurization at 149°C for 2 sec is used commonly in processing milk and we have observed this method not effective in processing vegetable pulp.
The requirement of different temperature and timing to make the vegetable pulp bacteria-free is expected to be due to the differences in tissue morphology, biochemical composition, texture and water content etc., of each vegetable.
| Table 1: | Generation time of pathogenic bacteria in broth culture |
 | |
| Table 2: | Optimization of conditions for the sterility of salad vegetable pulps |
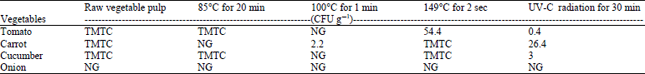 | |
| TMTC: Too many to count NG: No growth, CFU: Colony forming unit | |
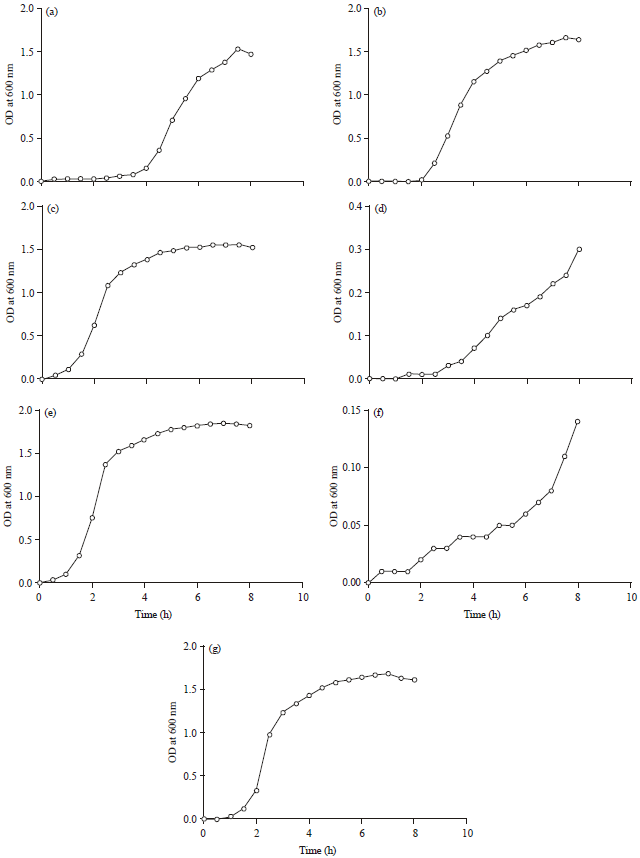 | |
| Fig. 1(a-g): | Generation time of pathogenic bacteria in broth culture, growth curve for (a) Listeria monocytogenes, (b) Bacillus cereus, (c) E. coli, (d) Shigella flexneri, (e) Aeromonas hydrophila, (f) Staphylococcus aureus and (g) Salmonella enterica |
Moreover, the phytochemical constituents are different in fresh-cut vegetables than in fresh vegetables (Beuchat, 1998). Among the vegetables used in the study, for fresh-cut carrot, very few reports on microbial quality and sanitization are available (Trinetta et al., 2011; Alarcon-Flores et al., 2014) and to our knowledge there are no reports in cucumber.
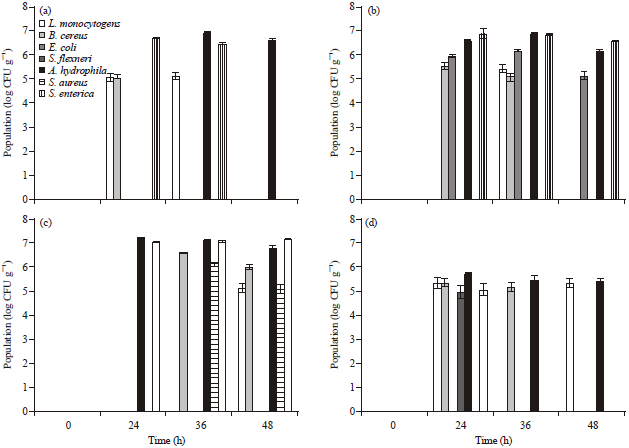 | |
| Fig. 2(a-d): | Population of pathogenic bacteria in inoculated vegetable pulp, (a) Tomato, (b) Carrot, (c) Cucumber and (d) Onion |
Cucumber is one of the major vegetable used in Indian vegetable salad and in many restaurants and bars cucumber is also used as sole vegetable salad. The current observations although needs extended validation with large number of samples and in different varieties of these vegetables, we believe that the sterilization conditions reported here could serve as starting point towards developing sanitization methods of ready-to-eat salad vegetables which could increase consumer safety.
The results of bacterial population in salad pulp are shown in Fig. 2. At the initial point of 0 h in 106th dilution the bacterial colonies were not found in any of the inoculated pulp, since the bacterial load was diluted 10,00,000 times from its initial concentration. Listeria monocytogenes, A. hydrophila and S. enterica were found to grow in tomato pulp. Bacillus cereus colonies were observed at 24 h which were not found later. The population of L. monocytogenes was comparatively less in 24 h which remained unaltered at 36 h. Aeromonas hydrophila colonies were observed late at 36 h but was the highest in number compared to other bacteria tested. The population started to decline at 48 h. Number of S. enterica colonies were high at 24 h (during which there was no growth of A. hydrophila). Tomato pulp was found to support growth of Salmonella within 24 h whereas A. hydrophila grows at 36 h. In the case of carrot pulp, L. monocytogenes, B. cereus, E. coli, A. hydrophila and S. enterica were found to multiply. In contrast to what was observed in tomato, the population of A. hydrophila as well as S. enterica reached more than 6 log CFU g-1 at 24 h. The population of these two bacteria remained high for another 12 h (observation at 36 h) which later reduced at 48 h. However, the population of S. enterica at 48 h was in similar densities as observed at 24 h, whereas the population of A. hydrophila reduced at this time point. Cucumber pulp supported the growth of L. monocytogenes, B. cereus, A. hydrophila, S. aureus and S. enterica. A. hydrophila population was very high (more than 7 log CFU g-1) at 24 h which later reduced with time. S. enterica population raised from less than 7 log CFU g-1 at 24 h to more than 7 log Cfu g-1 at 48 h. Although, L. monocytogenes, B. cereus, S. flexneri, A. hydrophila and S. enterica grew on onion pulp, compared to other vegetables tested, the population was less.
Our experiments were initially done with undiluted pulp samples. Since the number of colonies of bacteria was too many to count, we preferred 10-6 dilution of inoculated pulp. Hence the ‘no growth’ observed in plating does not really mean absence of bacteria but no observable colony at the dilution used (10-6). Similar ‘no growth’ was observed at 0 h of inoculation could raise the question of initial inoculum used. The inoculum load used were 2000 plus CFU mL-1 of bacterial culture. One milliliter of this culture was mixed with 100 mL of vegetable pulp obtained from homogenizing 50 g of tissues. At 0 h, 1 mL of the inoculated pulp was diluted 10-6 times and plated. For example, 2400 CFU mL-1 of culture of L. monocytogenes is diluted in 100 mL homogenized vegetable tissue to form 24 CFU mL-1 of pulp which further gets diluted to 10-6 times which is 0.000024 CFU mL-1. This is the reason why we observed ‘no growth’ at 0 h. Based on our trial experiments, 10-6 dilution was found to reveal a distinguishable difference in the population of bacteria in different vegetable pulp.
The pH of tomato, carrot, cucumber and red onions is 4.3-4.9, 5.3-5.8, 5.8-6.4 and 5.12-5.78, respectively (Anonymous, 1962). Since the pH of onion is similar to carrot and cucumber, we believe that the observed less growth in onion pulp may not be due to pH. Onions are known for their antimicrobial properties (Farbman et al., 1993; Zohri et al., 1995; Benmalek et al., 2013) and hence our observation of poor growth of bacteria in onion pulp is not surprising. Moreover, the onion pulp used for inoculation was free of any bacteria and hence did not require a sterilization step. Listeria monocytogenes and A. hydrophila were able to grow to some extent and reasonable number of colonies was observed until 48 h of inoculation. This indicates the possible resistance of these strains developed against the antimicrobial compounds of onion.
Occurrence of E. coli and Staphylococcus in carrots, B. cereus and L. monocytogenes in cucumber, L. monocytogenes and S. enterica in tomato have been reported. Many species of Aeromonas dominated by A. schubertii followed by A. hydrophila were found in organic vegetables including carrot, tomatoes and onions in Northern Ireland (McMahon and Wilson, 2001). Salmonella, E. coli 0157:H7, Staphylococcus and Listeria are known to colonize and survive in plants using them as alternate hosts (Schikora et al., 2012). Extensive work has been done on enteropathogenic bacteria in tomatoes particularly with reference to Salmonella. Literature is deficient in the studies on growth of human pathogenic bacteria in other vegetables like carrot, cucumber and onion although occurrences of these bacteria in them are reported. We have been studying the alternate ecology of food-borne human enteric bacteria in fruits and vegetables and we have reported Staphylococcus in apples (Phukon et al., 2013) and B. anthracis like strain in onions (Srinath et al., 2014).
Most of the work so far carried out on population dynamics of food-borne bacteria focused on fruit pulp and juices (Ukuku and Sapers, 2007; Penteado and Leitao, 2004; Khan et al., 2007; Pathanibul et al., 2009). Among vegetables, tomato has been studied extensively. The growth of seven different pathogenic bacteria in vegetable pulp including tomato was reported. Ability of A. hydrophila to grow in all these vegetables provides clues for the future extensive work needed to be done on this bacteria and its interaction in agricultural produce. To proliferate inside plant tissues, these enteric pathogens may rely on their ability to acquire simple carbon sources which has been proved by transcriptome analysis of E. coli growth in lettuce leaf lysates (Kyle et al., 2010). Salmonella grows well in tomato due to the amount of simple sugars, sugar alcohols, organic and fatty acids and amino acids present in them (Noel et al., 2010). The observed difference in the ability of different bacteria to grow differently in tomato, carrot, cucumber and onion pulp warrants development of vegetable specific sanitizing methods to avoid salad-borne entero-pathogenic infection.
ACKNOWLEDGMENT
The authors acknowledge the support rendered by the management of VIT University, Vellore, India for carrying out the study.
REFERENCES
- Rodoni, L.M., A. Concellon, A.R. Chaves and A.R. Vicente, 2012. Use of UV-C treatments to maintain quality and extend the shelf life of green fresh-cut bell pepper (Capsicum annuum L.). J. Food Sci., 77: C632-C639.
CrossRef - Beuchat, L.R., 2002. Ecological factors influencing survival and growth of human pathogens on raw fruits and vegetables. Microbes Infect., 4: 413-423.
CrossRefDirect Link - Noel, J.T., N. Arrach, A. Alagely, M. McClelland and M. Teplitski, 2010. Specific responses of Salmonella enteric to tomato varieties and fruit ripeness identified by in vivo expression technology. PLoS One, Vol. 5.
CrossRefDirect Link - Nithya, A., K.M. Gothandam and S. Babu, 2014. Alternative ecology of human pathogenic bacteria in fruits and vegetables. Plant Pathol. J., 13: 1-7.
CrossRefDirect Link - Gorbatsevich, E., S.S. Saldinger, R. Pinto and N. Bernstein, 2013. Root internalization, transport and in-planta survival of Salmonella enteric serovar Newport in sweet basil. Environ. Microbiol. Rep., 5: 151-159.
CrossRefDirect Link - Brewer, M.S., G.K. Sprouls and C. Russon, 1994. Consumer attitudes toward food safety issues. J. Food Saf., 14: 63-76.
CrossRefDirect Link - Trinetta, V., N. Vaidya, R. Linton and M. Morgan, 2011. A comparative study on the effectiveness of chlorine dioxide gas, ozone gas and e-beam irradiation treatments for inactivation of pathogens inoculated onto tomato, cantaloupe and lettuce seeds. Int. J. Food Microbiol., 146: 203-206.
CrossRefDirect Link - Lopez-Galvez, F., G.D. Posada-Izquierdo, M.V. Selma, F. Perez-Rodriguez, J. Gobet, M.I. Gil and A. Allende, 2012. Electrochemical disinfection: An efficient treatment to inactivate Escherichia coli O157:H7 in process wash water containing organic matter. Food Microbiol., 30: 146-156.
CrossRefDirect Link - Rahman, S.M.E., Y.G. Jin and D.H. Oh, 2011. Combination treatment of alkaline electrolyzed water and citric acid with mild heat to ensure microbial safety, shelf-life and sensory quality of shredded carrots. Food Microbiol., 28: 484-491.
CrossRefDirect Link - Erickson, M.C., 2010. Microbial risks associated with cabbage, carrots, celery, onions and deli salads made with these produce items. Compr. Rev. Food Sci. Food Saf., 9: 602-619.
CrossRefDirect Link - Alarcon-Flores, M.I., R. Romero-Gonzalez, J.L.M. Vidal, F.J.E. Gonzalez and A.G. Frenich, 2014. Monitoring of phytochemicals in fresh and fresh-cut vegetables: A comparison. Food Chem., 142: 392-399.
CrossRefDirect Link - Farbman, K.S., E.D. Barnett, G.R. Bolduc and J.O. Klein, 1993. Antibacterial activity of garlic and onions: A historical perspective. Pediatr. Infect. Dis. J., 12: 613-614.
Direct Link - Zohri, A.N., K. Abdel-Gawad and S. Saber, 1995. Antibacterial, antidermatophytic and antitoxigenic activities of onion (Allium cepa L.) oil. Microbiol. Res., 150: 167-172.
CrossRefDirect Link - Benmalek, Y., O.A. Yahia, A. Belkebir and M.L. Fardeau, 2013. Anti-microbial and anti-oxidant activities of Illicium verum, Crataegus oxyacantha ssp monogyna and Allium cepa red and white varieties. Bioengineered, 4: 244-248.
CrossRefPubMedDirect Link - McMahon, M.A.S. and I.G. Wilson, 2001. The occurrence of enteric pathogens and Aeromonas species in organic vegetables. Int. J. Food Microbiol., 70: 155-162.
CrossRefDirect Link - Schikora, A., A.V. Garcia and H. Hirt, 2012. Plants as alternative hosts for Salmonella. Trends Plant Sci., 17: 245-249.
CrossRefPubMedDirect Link - Phukon, M., P. Sahu, R. Srinath, A. Nithya and S. Babu, 2013. Unusual occurrence of Staphylococcus warneri as endophyte in fresh fruits along with usual Bacillus spp. J. Food Saf., 33: 102-106.
CrossRefDirect Link - Srinath, R., R. Siva and S. Babu, 2014. Bacillus anthracis-like strain-carrying Pseudomonas FPVA gene occurs as endophyte in vegetables. J. Food Saf., 34: 57-61.
CrossRefDirect Link - Ukuku, D.O. and G.M. Sapers, 2007. Effect of time before storage and storage temperature on survival of Salmonella inoculated on fresh-cut melons. Food Microbiol., 24: 288-295.
CrossRefDirect Link - Penteado, A.L. and M.F.F. Leitao, 2004. Growth of Salmonella enteritidis in melon, watermelon and papaya pulp stored at different times and temperatures. Food Control, 15: 369-373.
CrossRefDirect Link - Khan, A.A., C.D. Melvin and E.B. Dagdag, 2007. Identification and molecular characterization of Salmonella spp. From unpasteurized orange juices and identification of new serotype Salmonella strain S. enteric serovar Tempe. Food Microbiol., 24: 539-543.
CrossRefDirect Link - Pathanibul, P., T.M. Taylor, P.M. Davidson and F. Harte, 2009. Inactivation of Escherichia coli and Listeria innocua in apple and carrot juices using high pressure homogenization and nisin. Int. J. Food Microbiol., 129: 316-320.
CrossRefPubMedDirect Link - Kyle, J.L., C.T. Parker, D. Goudeau and M.T. Brandl, 2010. Transcriptome analysis of Escherichia coli O157:H7 exposed to lysates of lettuce leaves. Applied Environ. Microbiol., 76: 1375-1387.
CrossRefDirect Link